
Verges as Fragments of Loess Grasslands in the Carpathian Basin and Their Festuca Species

 , ,
, ,
Abstract
:1. Introduction
- Is there a correlation between the different ages of the verges (documented based on maps) and the patches of remnant vegetation that have developed on them, and whether the older ones are more valuable?
- Is there a correlation between the types, location, expanse and vegetation types of the verges?
- Which of the coenosystematically important species of the genus Festuca occur in the verges and patches of loess grassland?
2. Materials and Methods
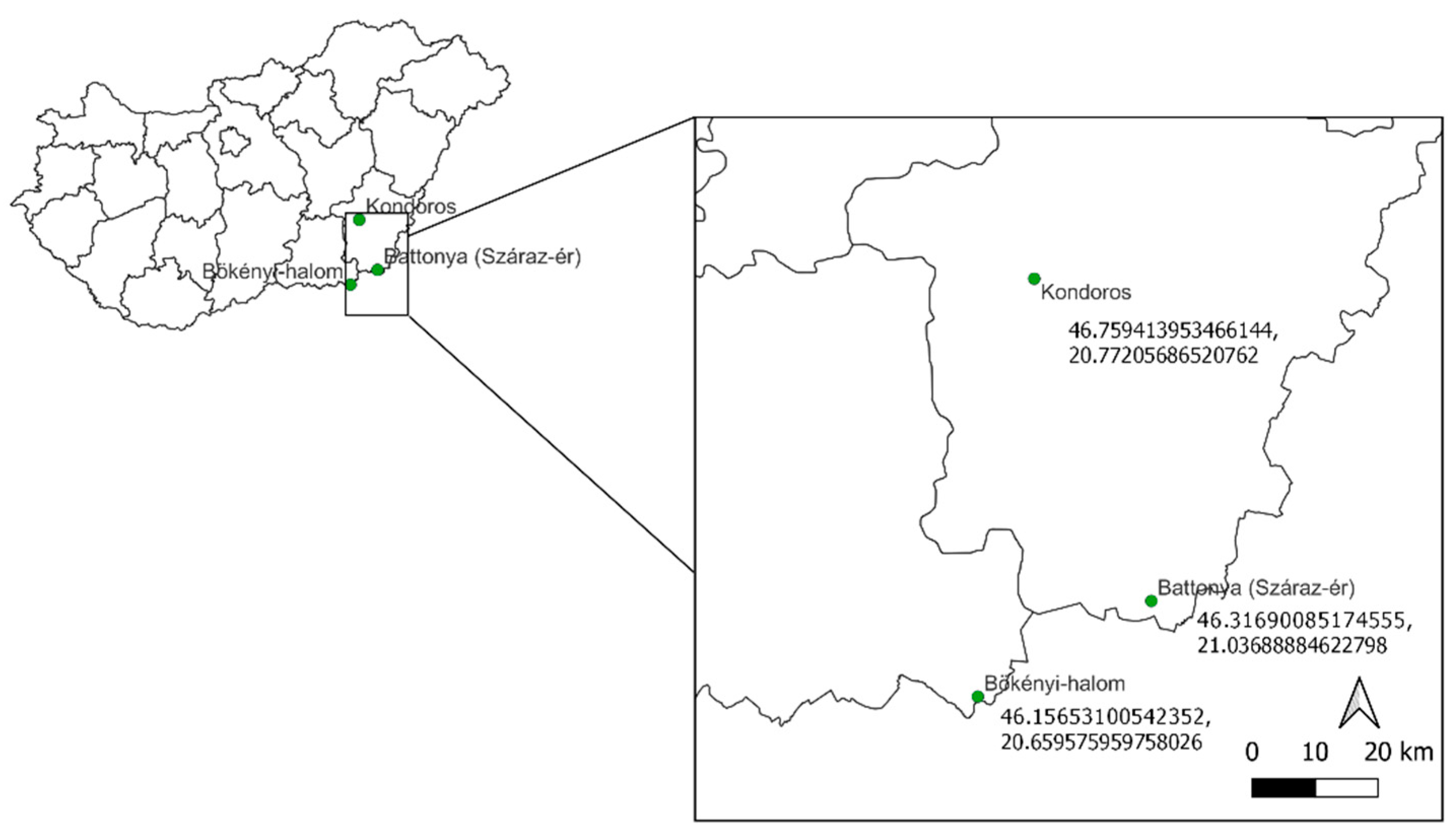
2.1. Surveyed Areas and Their Age
- I.
- group: natural condition indicative taxa
- II.
- group: degradation indicator taxa
2.2. Diversity Studies
2.3. Taxonomy Investigations
3. Results
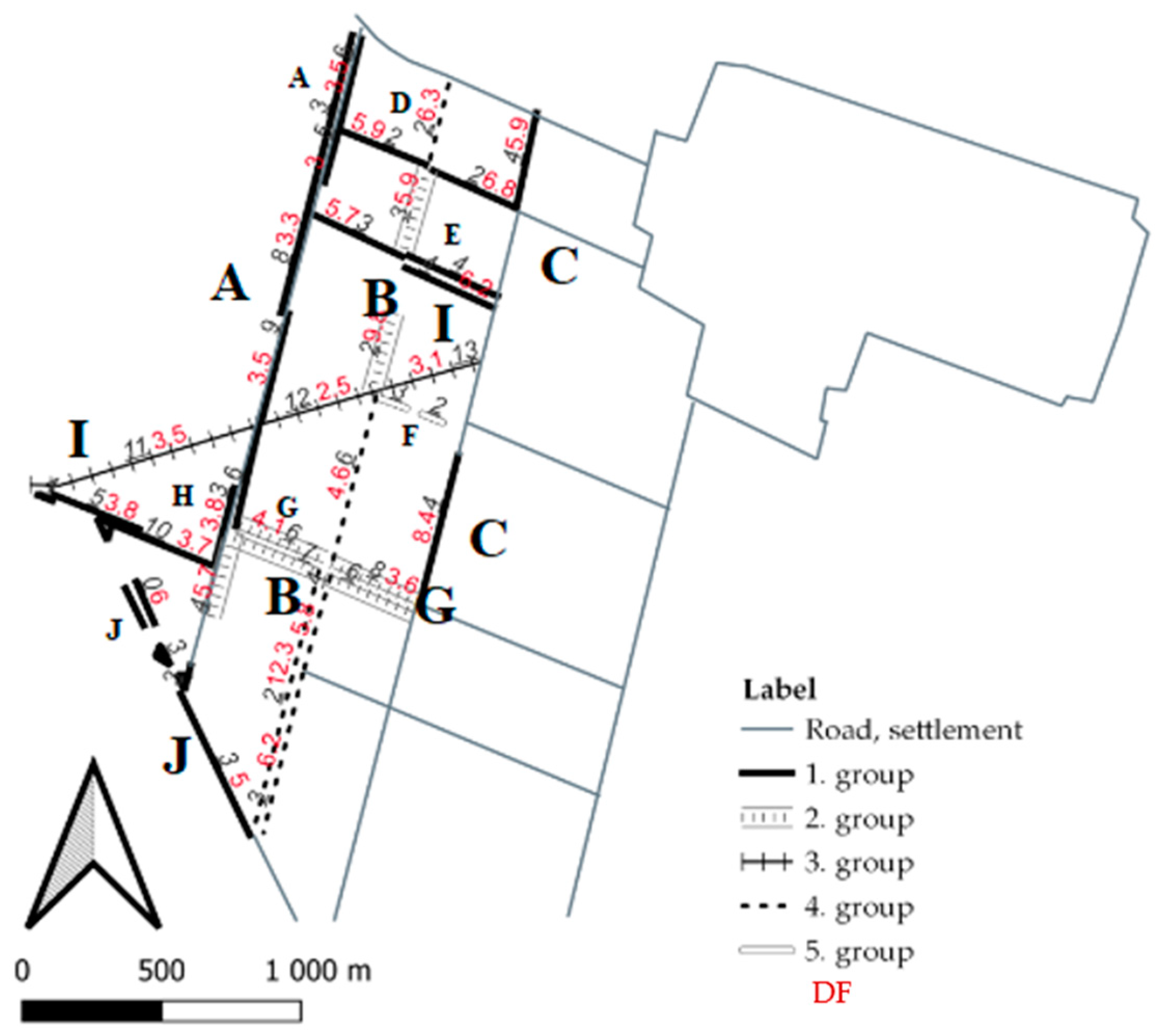
3.1. Ages and Natural Conditions of the Verges
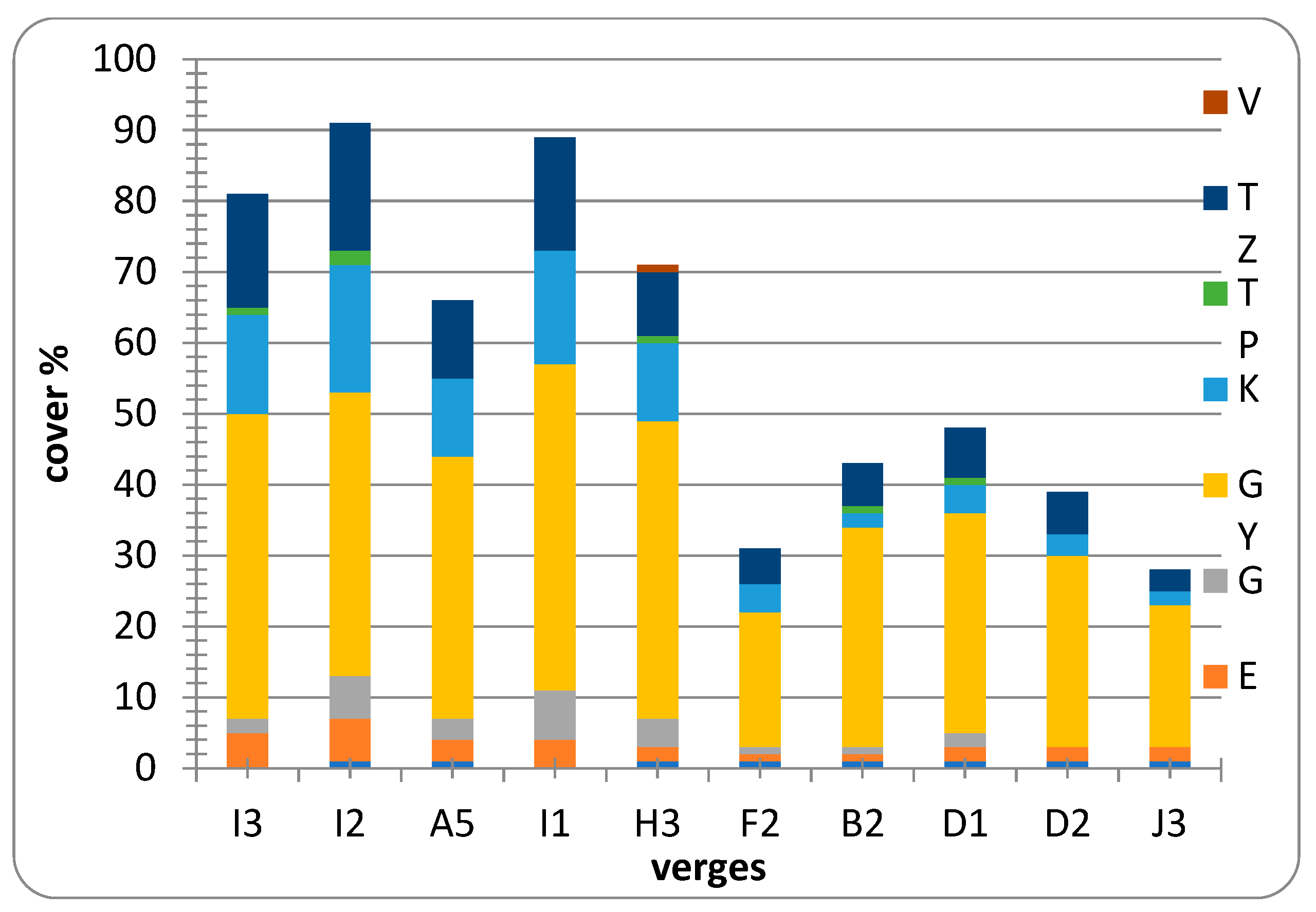
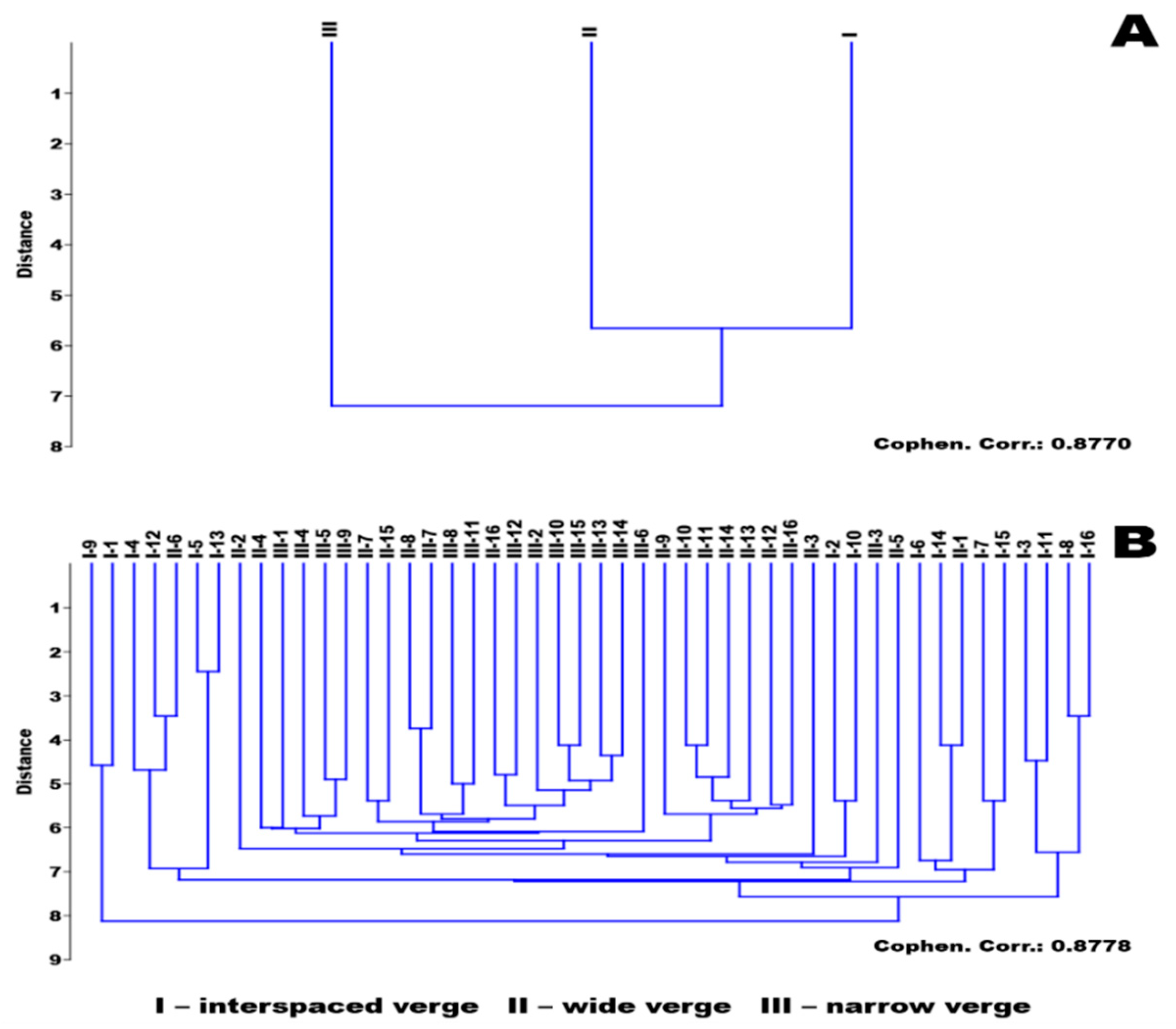
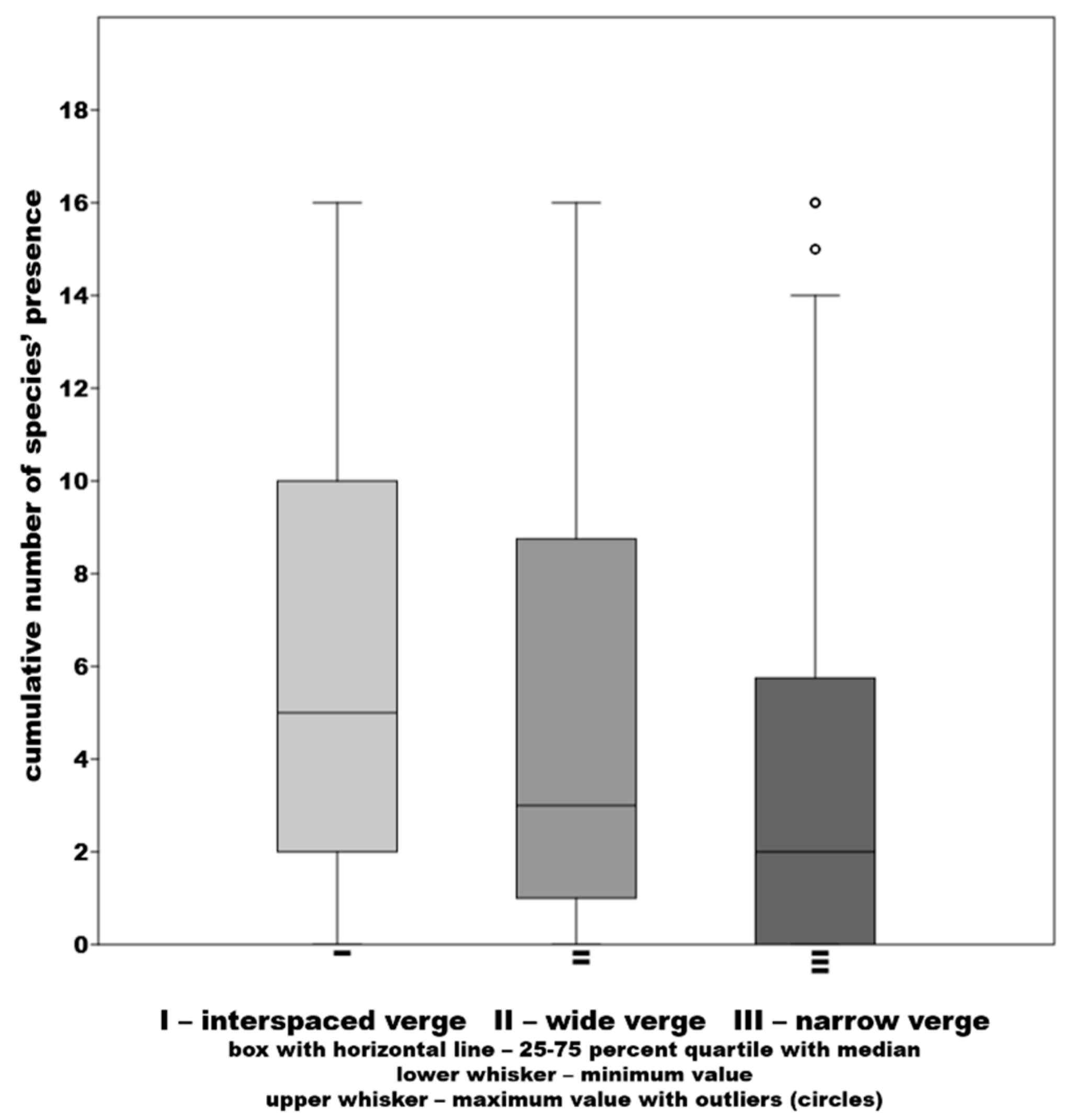
3.2. Results of the Diversity Study of the Verges
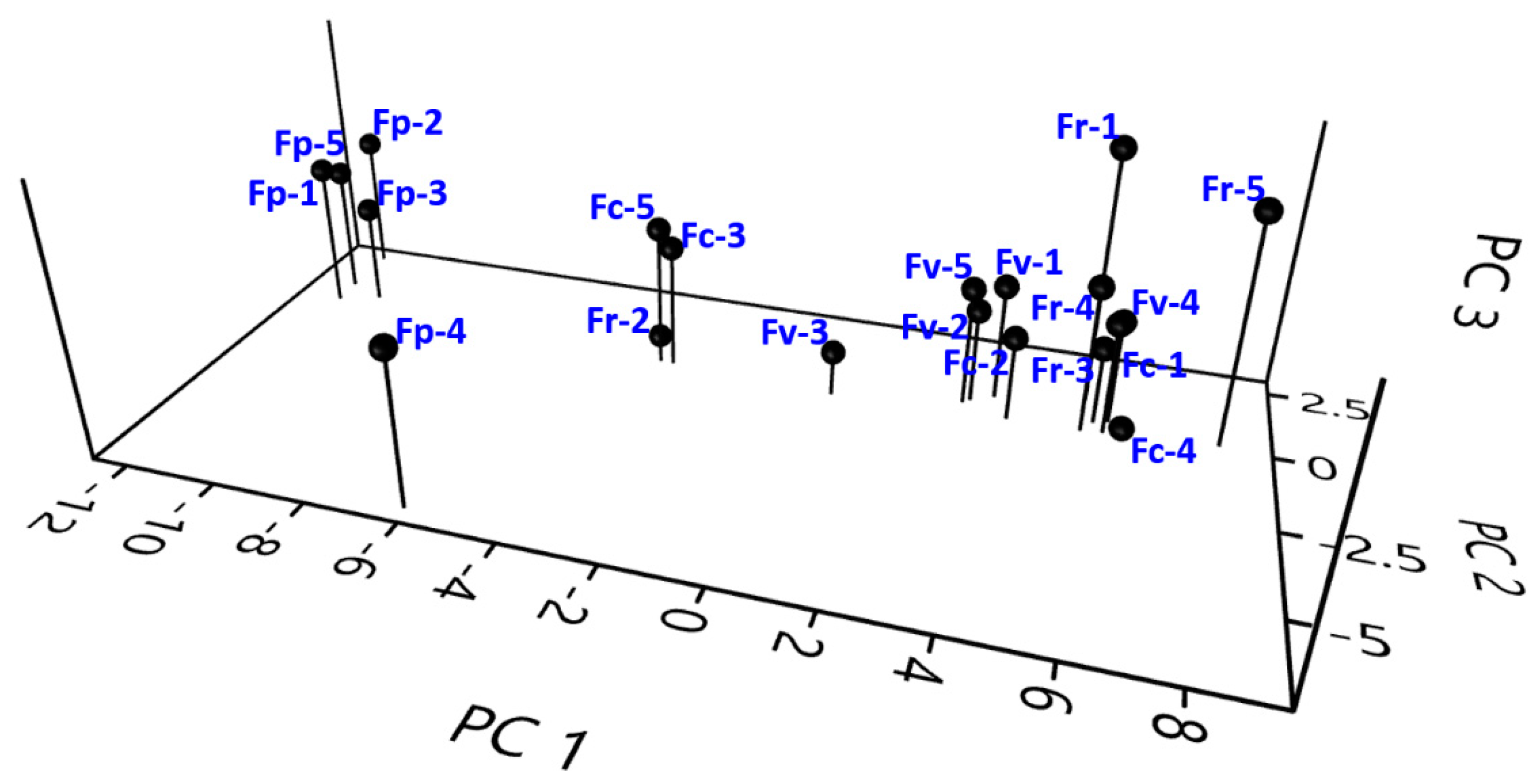
3.3. Results of Taxonomy Investigation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóth, E.; Tóthmérész, B. Managing for composition or species diversity?—Pastoral and year-round grazing systems in alkali grasslands. Agric. Ecosyst. Environ. 2016, 234, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef] [PubMed]
- Erdős, L.; Tolgyesi, C.; Cseh, V.; Tolnay, D.; Cserhalmi, D.; Körmöczi, L.; Gellény, K.; Bátori, Z. Vegetation history, recent dynamics and future prospects of a Hungarian sandy forest-steppe reserve: Forest-grassland relations, tree species composition and size-class distribution. Community Ecol. 2015, 16, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Krstonošić, D.; Kiss, P.J.; Bátori, Z.; Tölgyesi, C.; Škvorc, Ž. Plant composition and diversity at edges in a semi-natural forest–grassland mosaic. Plant Ecol. 2019, 220, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past Trends, Present State and Future Prospects of Hungarian Forest-Steppes. In Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World; Werger, M.J.A., Staalduinen, M.A., Eds.; Springer Science & Business Media: New York, NY, USA, 2012; Volume 6, pp. 209–252. [Google Scholar]
- Wesche, K.; Ambarlı, D.; Kamp, J.; Török, P.; Treiber, J.; Dengler, J. The Palaearctic steppe biome: A new synthesis. Biodivers. Conserv. 2016, 25, 2197–2231. [Google Scholar] [CrossRef]
- Bátori, Z.; Erdős, L.; Kelemen, A.; Deák, B.; Valkó, O.; Gallé, R.; Bragina, T.M.; Kiss, P.J.; Kröel-Dulay, G.; Tölgyesi, C. Diversity patterns in sandy forest-steppes: A comparative study from the western and central Palearctic. Biodivers. Conserv. 2018, 27, 1011–1030. [Google Scholar] [CrossRef]
- Dengler, J.; Janišová, M.; Török, P.; Wellstein, C. Biodiversity of Palaearctic grasslands: A synthesis. Agric. Ecosyst. Environ. 2014, 182, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Walter, H.; Lieth, H. Klimadiagramm-Weltatlas, 3rd ed.; Gustav Fischer: Jena, Germany, 1967. [Google Scholar]
- Willner, W.; Kuzemko, A.; Dengler, J.; Chytrý, M.; Bauer, N.; Becker, T.; Biţă-Nicolae, C.; Botta-Dukát, Z.; Čarni, A.; Csiky, J.; et al. A higher-level classification of the Pannonian and western Pontic steppe grasslands (Central and Eastern Europe). Appl. Veg. Sci. 2016, 20, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Zólyomi, B.; Kéri, M.; Horváth, F. Spatial and temporal changes in the frequency of climatic year types in the Carpathian Basin. Coenoses 1997, 12, 33–41. [Google Scholar]
- Hartel, T.; Dorresteijn, I.; Klein, C.; Máthé, O.; Moga, C.I.; Öllerer, K.; Roellig, M.; von Wehrden, H.; Fischer, J. Wood-pastures in a traditional rural region of Eastern Europe: Characteristics, management and status. Biol. Conserv. 2013, 10, 267–275. [Google Scholar] [CrossRef]
- Csontos, P.; Tamás, J. Fás mezsgyék növényzetének vizsgálata a Balaton-felvidéken. Acta Agron. Ovariensis 2007, 49, 3–13. [Google Scholar]
- Szerényi, J.; Kalapos, T. Löszpusztai vegetáció maradványai az Érd-százhalombattai Sánc-hegyen. In Proceedings of the Lippay János—Vas Károly Tudományos Ülésszak, Szt. István Egyetem, Budapest, Hungary, 6–7 November 2000. [Google Scholar]
- Barczi, A.; Penksza, K.; Joó, K. Alföldi kunhalmok talaj-növény összefüggés-vizsgálata. Agrokémia Talajt. 2004, 53, 3–16. [Google Scholar] [CrossRef]
- Csathó, A.I. A mezsgyék természetvédelmi jelentősége az Alföld löszvidékein. Tájökológiai Lapok 2005, 3, 363–364. [Google Scholar]
- Csathó, A.I. Mezsgyék kutatása a Körös-Maros Nemzeti Park Igazgatóság működési területén. In Kutatási Jelentés; KMNPI: Szarvas, Hungary, 2008. [Google Scholar]
- Csathó, A.I. Előzetes javaslatok a mezsgyék mintavételezéséhez. In Magyar Ökológus Kongresszus, Előadások és poszterek összefoglalói; MÖK: Szeged, Hungary, 2009; Volume 8, p. 41. [Google Scholar]
- Csathó, A.I. Az elsődleges és másodlagos mezsgyék növényzetének összehasonlító vizsgálata a battonyai Gránic és Csárda-dűlő példáján. Tájökológiai Lapok 2011, 9, 345–356. [Google Scholar]
- Tóth, A. A kunhalom-kérdésről. In A kunhalmokról más Szemmel; Tóth, A., Ed.; Alföldkutatásért Alapítvány: Kisújszállás-Debrecen, Hungary, 2004; pp. 7–12. [Google Scholar]
- Deák, B.; Tóthmérész, B.; Valkó, O.; Sudnik-Wójcikowska, B.; Bragina, T.M.; Moysiyenko, I.; Bragina, T.M.; Apostolova, I.; Dembicz, I.; Bykov, N.I.; et al. Cultural monuments and nature conservation: The role of kurgans in maintaining steppe vegetation. Biodivers. Conserv. 2016, 25, 2473–2490. [Google Scholar] [CrossRef]
- Sudnik-Wójcikowska, B.; Moysiyenko, I.I.; Zachwatowicz, M.; Jabłońska, E. The value and need for protection of kurgan flora in the anthropogenic landscape of steppe zone in Ukraine. Plant Biosyst. 2011, 145, 638–653. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Tóthmérész, B. Factors threatening grassland specialist plants—A multi-proxy study on the vegetation of isolated grasslands. Biol. Conserv. 2016, 204, 255–262. [Google Scholar] [CrossRef]
- Deák, B.; Bede, Á.; Rádai, Z.; Tóthmérész, B.; Török, P.; Torma, A.; Lőrinczi, G.; Nagy, A.; Mizser, S.; Kelemen, A.; et al. Different extinction debts among plants and arthropods after loss of grassland amount and connectivity. Biol. Conserv. 2021, 264, 109372. [Google Scholar] [CrossRef]
- Deák, B.; Rádai, Z.; Bátori, Z.; Kelemen, A.; Lukács, K.; Kiss, R.; Mák, E.I.; Valkó, O. Ancient burial mounds provide safe havens for grassland specialist plants in transformed landscapes—A trait-based analysis. Front. Ecol. Evol. 2021, 9, 619812. [Google Scholar] [CrossRef]
- Kiss, I. Ősgyep-maradvány az orosházi Nagytatársáncon. Acta Acad. Paedagog. Szeged. 1968, 2, 39–61. [Google Scholar]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Pauchard, A.; García, A.; Zalba, S.; Sarasola, M.; Zenni, R.; Ziller, S.; Nuñez, M.A. Pine Invasions in South America: Reducing Their Ecological Impacts Through Active Management. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning Clode, J., Ed.; De Gruyter Open Ltd.: Warsaw, Poland, 2016; pp. 318–342. [Google Scholar]
- Bajor, Z.; Zimmermann, Z.; Szabó, G.; Fehér, Z.; Járdi, I.; Lisztes-Szabó, Z.; Penksza, K. Effect of conservation management practices on sand grassland vegetation in Budapest, Hungary. Appl. Ecol. Environ. Res. 2016, 14, 233–247. [Google Scholar] [CrossRef]
- Soó, R. A magyar flóra és vegetáció rendszertani-növényföldrajzi kézikönyve I. In Synopsis Systematico-Geobotanica Florae Vegetationisque Hungariae I; Akadémia Kiadó: Budapest, Hungary, 1964. [Google Scholar]
- Kiss, I. Az Adonis volgensis lelőhelyei és népies gyógyászati vonatkozásai. Magyarországon. Acta Academ. Pedagog. Szeged. 1964, 2, 25–51. [Google Scholar]
- Bagaria, G.; Helm, A.; Rod`a, F.; Pino, J. Assessing coexisting plant extinction debt and colonization credit in a grassland–forest change gradient. Oecologia 2015, 179, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Biro, M.; Bölöni, J.; Molnár, Z. Use of long-term data to evaluate loss and endangerment status of Natura 2000 habitats and effects of protected areas. Conserv. Biol. 2018, 3, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Deák, B.; Tölgyesi, C.; Kelemen, A.; Bátori, Z.; Gallé, R.; Bragina, T.M.; Abil, Y.A.; Valkó, O. The effects of micro-habitats and grazing intensity on the vegetation of burial mounds in the kazakh steppes. Plant Ecol. Divers. 2017, 10, 509–520. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Risto, K.; Heikkinen, R.; Helm, A.; Krauss, J.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; et al. Extinction debt: A challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef]
- Joanne Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2004, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Archer, S.; Bouton, T.W.; Hibbard, K.A. Trees in Grasslands: Biogeochemical Consequences of Woody Plant Expansion. In Global Biogeochemical Cycles and Their Interrelationship with Climate; Schulze, E.A., Harrison, S.P., Heimann, M., Holland, E.A., Lloyd, J., Prentice, I.C., Schimel, D.S., Eds.; Academic Press: London, UK, 2001; pp. 115–130. [Google Scholar]
- Yezzi, A.; Nebbia, A.; Zalba, S. Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa. Diversity 2021, 13, 637. [Google Scholar] [CrossRef]
- Penksza, K.; Loksa, G.; Barczi, A.; Joó, K.; Malatinszky, Á. Effects of extrazonal and climatic conditions on the vegetation of kurgans. A pilot study from the Hortobágy (Csípő-halom). In Kurgan studies: An Environmental and Archaeological Multiproxy Study of Burial Mounds in the Eurasian Steppe Zone; Pető, Á., Barczi, A., Eds.; BAR International Series: Oxford, UK, 2011; pp. 347–350. [Google Scholar]
- Deák, B.; Valkó, O.; Török, P.; Kelemen, A.; Miglécz, T.; Szabó, S.; Szabó, G.; Tóthmérész, B. Micro-topographic heterogeneity increases plant diversity in old stages of restored grasslands. Basic Appl. Ecol. 2015, 16, 291–299. [Google Scholar] [CrossRef]
- Rákóczi, A.; Barczi, A. Protected landscape elements in the European Union, the influence of EC decree 73/2009 on the condition of Hungarian mounds. Tájökológiai Lapok 2014, 12, 95–105. [Google Scholar]
- Tóth, C.A.; Deák, B.; Nyilas, I.; Bertalan, L.; Valkó, O.; Novák, T. Iron age burial mounds as refugia for steppe specialist plants and invertebrates—Case study from the Zsolca mounds (NE Hungary). Hacquetia 2019, 18, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Vona, M.; Penksza, K. A szentesi Kántor-halom vegetációjának változása és ennek összefüggése a talaj vízháztartásával. Tájökológiai Lapok 2004, 2, 341–348. [Google Scholar]
- Kiss, T.; Penksza, K. A legeltetés hosszú távú hatása kiskunsági füves pusztákon. Természetvédelmi Közlemények 2018, 24, 104–113. [Google Scholar] [CrossRef]
- Kiss, T.; Penksza, K.; Tasi, J.; Szentes, S. Juh-és marhalegelő cönológia és gyepgazdálkodási vizsgálata kiskunsági területeken. Gyepgazdálkodási Közlemények 2008, 6, 39–45. [Google Scholar]
- Kiss, T.; Lévai, P.; Ferencz, Á.; Szentes, S.; Hufnagel, L.; Nagy, A.; Balogh, Á.; Pintér, O.; Saláta, D.; Házi, J.; et al. Change of composition and diversity of species and grassland management between different grazing intensity—In Pannonian dry and wet grasslands. Appl. Ecol. Environ. Res. 2011, 9, 197–230. [Google Scholar] [CrossRef]
- Herczeg, E. Botanikai vizsgálatok kunhalmok Dél-tiszántúli löszgyepein. Kanitzia 2005, 13, 45–54. [Google Scholar]
- Herczeg, E.; Pottyondy, Á.; Penksza, K. Cönológiai vizsgálatok eltérő gazdálkodású dél–tiszántúli löszgyepekben. Tájökológiai Lapok 2005, 3, 259–265. [Google Scholar]
- Nagy, A.; Penksza, K. Élőhely–értékelési lehetőségek dél–tiszántúli és veresegyházi területeken természetességi mutatók alapján. Tájökológiai Lapok 2006, 4, 115–125. [Google Scholar]
- Nagy, A.; Penksza, K. A Vésztő–Mágor Természetvédelmi Terület élőhelytérképe, és környezetgazdálkodási–természetvédelmi értékelési lehetősége. Tájökológiai Lapok 2007, 5, 103–116. [Google Scholar]
- Nagy, A.; Penksza, K.; Laborczi, A.; Kiss, T. Habitat mapping of Vésztő–Mágorpuszta (South–East Hungary) protected natural area. Lucr. Ştitinţifice 2007, 9, 125–132. [Google Scholar]
- Penksza, K. Die Koerrektur der histologischen Beschreibung von Festuca javorkae von Májovszky im Jahre 1962, und Angaben zum Vorkommnis der Art in Ungarn. Ber. Inst. Landsch. Pflanz. Univ. Hohenh. 2000, 10, 49–54. [Google Scholar]
- Penksza, K. Poaceae—Pázsitfüvek nemzetségeinek határozókulcsa. Festuca—Csenkeszek, Lolium—Vadóc, Festulolium—Korcsvadóc. In Új Magyar Fűvészkönyv; Király, G., Ed.; Aggteleki Nemzeti Park: Aggtelek, Hungary, 2009; pp. 498–509. [Google Scholar]
- Penksza, K.; Csík, A.; Filep, A.F.; Saláta, D.; Pápay, G.; Kovács, L.; Varga, K.; Pauk, J.; Lantos, C.; Lisztes-Szabó, Z. Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa. Forests 2020, 11, 1325. [Google Scholar] [CrossRef]
- Penksza, K.; Saláta, D.; Pápay, G.; Péter, N.; Bajor, Z.; Lisztes-Szabó, Z.; Fűrész, A.; Fuchs, M.; Michéli, E. Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes? Forests 2021, 12, 114. [Google Scholar] [CrossRef]
- Galli, Z.; Penksza, K.; Kiss, E.; Sági, L.; Heszky, L.E. Low variability of Internal Transcribed Spacer rDNA and trnL (UAA) intron sequences of several taxa in the Festuca ovina aggregate (Poaceae). Acta Biol. Hung. 2006, 57, 57–69. [Google Scholar] [CrossRef]
- Bauer, L.; Galli, Z.; Penksza, K.; Engloner, A.; Szerdahelyi, T.; Kiss, E.; Heszky, L. Morfológiai és molekuláris taxonómiai vizsgálatok kárpát–medencei Festuca fajokon. In Kárpát–Medencei Biológiai Szimpózium; MBT: Budapest, Hungary, 2003; Volume III, pp. 33–37. [Google Scholar]
- Markgraf-Dannenberg, I. Festuca L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; Volume 5, pp. 125–153. [Google Scholar]
- Barczi, A.; Penksza, K.; Joó, K. Reseach of soil–plant connections on Kurgans in Hungary. Ekológia 2004, 23, 15–22. [Google Scholar]
- Dövényi, Z. Magyarország Kistájainak Katasztere; Magyar Tudományos Akadémia, Földrajztudományi Kutatóintézet: Budapest, Hungary, 2010; pp. 34–56. [Google Scholar]
- Available online: https://maps.arcanum.com/hu/map/firstsurvey-lower-austria/?bbox=1859421.4186977376%2C6117715.954432676%2C1962534.9698544405%2C6155399.409377268&map-list=1&layers=151 (accessed on 2 May 2022).
- Térképek. Magyar Királyság (1819–1869)—Második Katonai Felmérés. Available online: https://maps.arcanum.com/hu/map/secondsurvey-hungary/?layers=5&bbox=2108467.652103985%2C6019361.428761189%2C2134246.039893161%2C6027960.594443272 (accessed on 2 May 2022).
- Térképek. Habsburg Birodalom (1869–1887)—Harmadik Katonai Felmérés (1:75000). Available online: https://maps.arcanum.com/hu/map/thirdsurvey75000/?layers=43&bbox=1900697.477281965%2C6130610.723300188%2C1913586.671176553%2C6134910.306141228 (accessed on 2 May 2022).
- Arcanum. IV. A Negyedik Katonai Felmérés (1896–1914). Available online: https://www.arcanum.com/hu/online-kiadvanyok/Janko-janko-annamaria-magyarorszag-katonai-felmeresei-1/iv-a-negyedik-katonai-felmeres-1896-1914-27A/ (accessed on 2 May 2022).
- Simon, T. A Magyarországi Edényes Flóra Határozója; Tankönyvkiadó: Budapest, Hungary, 2000; pp. 34–56. [Google Scholar]
- Horváth, F.; Dobolyi, Z.K.; Morschhauser, T.; Lőkös, L.; Karas, L.; Szerdahelyi, T. FLÓRA adatbázis 1.2—Taxonlista és attribútum-állomány. In FLÓRA Munkacsoport; MTA-ÖBKI; MTM Növénytára: Vácrátót, Hungary, 1995. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie; Wien: New York, NY, USA, 1964; pp. 2–865. [Google Scholar]
- Király, G. Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei. Határozókulcsok; Aggteleki Nemzeti Park Igazgatóság: Jósvafő, Hungary, 2009; pp. 3–456. [Google Scholar]
- Hammer, Ø. PAST—PAleontological STatictics Version 3.06 Reference Manual; Natural History Museum: Oslo, Norway; University of Oslo: Oslo, Norway, 1999–2015; pp. 3–225. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST—Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2009, 4, 1–9. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tóthmérész, B. Comparison of different methods for diversity ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Penksza, K.; Szabó, G.; Zimmermann, Z.; Lisztes-Szabó, Z.; Pápay, G.; Járdi, I.; Fűrész, A.; Falusi, E.S. The taxonomic problems of the Festuca vaginata agg. and their coenosystematic aspects. A Festuca vaginata alakkör taxonómiai problematikája és ennek cönoszisztematikai vonatkozásai. Georg. Agric. 2019, 23, 63–76. [Google Scholar]
- Deák, B.; Valkó, O.; Nagy, D.D.; Török, P.; Torma, A.; Lőrinczi, G.; Kelemen, A.; Nagy, A.; Bede, A.; Mizser, S.; et al. Habitat islands outside nature reserves—Threatened biodiversity hotspots of grassland specialist plant and arthropod species. Biol. Conserv. 2020, 241, 108254. [Google Scholar] [CrossRef]
- Dembicz, I.; Szczeparska, L.; Moysiyenko, I.I.; Wódkiewicz, M. High genetic diversity in fragmented Iris pumila L. populations in Ukrainian steppe enclaves. Basic Appl. Ecol. 2018, 28, 37–47. [Google Scholar] [CrossRef]
- Dembicz, I.; Moysiyenko, I.I.; Kozub, L.; Dengler, J.; Zakharova, M.; Sudnik-Wójcikowska, B. Steppe islands in a sea of fields: Where island biogeography meets the reality of a severely transformed landscape. J. Veg. Sci. 2020, 21, e12930. [Google Scholar] [CrossRef]
- Helm, A.; Hanski, I.; Pärtel, M. Slow response of plant species richness to habitat loss and fragmentation. Ecol. Lett. 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Brady, M.; Kellermann, K.; Sahrbacher, C.; Jelinek, L. Impacts of decoupled agricultural support on farm structure, biodiversity and landscape mosaic: Some EU results. J. Agric. Econ. 2009, 60, 563–585. [Google Scholar] [CrossRef]
- Valkó, O.; Tóth, K.; Kelemen, A.; Miglécz, T.; Sonkoly, J.; Tóthmérész, B.; Török, P.; Deák, B. Cultural heritage and biodiversity conservation—Plant introduction and practical restoration on ancient burial mounds. Nat. Conserv. 2018, 24, 65–80. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Török, P.; Hábenczyus, A.A.; Bátori, Z.; Valkó, O.; Deák, B.; Tóthmérész, B.; Erdős, L.; Kelemen, A. Underground deserts below fertility islands?—Woody species desiccate lower soil layers in sandy drylands. Ecography 2020, 43, 848–859. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.C.; Chen, X.; Corlett, R.T.; Didham, R.K.; Ding, P.; Holt, R.D.; Holyoak, M.; Hu, G.; Hughes, A.C.; Jiang, L. Habitat fragmentation and biodiversity conservation: Key findings and future challenges. Landsc. Ecol. 2016, 31, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Šmarda, P.; Šmerda, J.; Knoll, A.; Bureš, P.; Danihelka, J. Revision of Central European taxa of Festuca ser. Psammophilae Pawlus: Morphometrical, karyological and AFLP analysis. Plant Syst. Evol. 2007, 266, 197–232. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I3 | I2 | A5 | I1 | H3 | F2 | B2 | D1 | D2 | J3 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Dƒ | 9.2 | 12.3 | 8.4 | 11.6 | 2.23 | 3.37 | 4.6 | 4.1 | 5.1 | 6.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szentes, S.; Sutyinszki, Z.; Kiss, T.; Fűrész, A.; Saláta, D.; Harkányiné Székely, Z.; Penksza, K. Verges as Fragments of Loess Grasslands in the Carpathian Basin and Their Festuca Species. Diversity 2022, 14, 510. https://doi.org/10.3390/d14070510
Szentes S, Sutyinszki Z, Kiss T, Fűrész A, Saláta D, Harkányiné Székely Z, Penksza K. Verges as Fragments of Loess Grasslands in the Carpathian Basin and Their Festuca Species. Diversity. 2022; 14(7):510. https://doi.org/10.3390/d14070510
Chicago/Turabian StyleSzentes, Szilárd, Zsuzsanna Sutyinszki, Tímea Kiss, Attila Fűrész, Dénes Saláta, Zsuzsanna Harkányiné Székely, and Károly Penksza. 2022. "Verges as Fragments of Loess Grasslands in the Carpathian Basin and Their Festuca Species" Diversity 14, no. 7: 510. https://doi.org/10.3390/d14070510