
Participation of Rossiulus kessleri (Diplopoda, Julida) in the Formation of Algae Assemblages of Urbanized Territories


Abstract
:1. Introduction
2. Material and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kajimoto, T. Factors affecting seedling recruitment and survivorship of the Japanese subalpine stone pine, Pinus pumila, after seed dispersal by nutcrackers. Ecol. Res. 2002, 17, 481–491. [Google Scholar] [CrossRef]
- Boch, S.; Berlinger, M.; Fischer, M.; Knop, E.; Nentwig, W.; Turke, M.; Prati, D. Fern and bryophyte endozoochory by slugs. Oecologia 2013, 172, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liehrmann, O.; Jegoux, F.; Guilbert, M.A.; Isselin-Nondedeu, F.; Said, S.; Locatelli, Y.; Baltzinger, C. Epizoochorous dispersal by ungulates depends on fur, grooming and social interactions. Ecol. Evol. 2018, 8, 1582–1594. [Google Scholar] [CrossRef]
- Silva, G.G.; Green, A.J.; Hoffman, P.; Weber, V.; Stenert, C.; Lovas-Kiss, A.; Maltchik, L. Seed dispersal by neotropical waterfowl depends on bird species and seasonality. Freshw. Biol. 2021, 66, 78–88. [Google Scholar] [CrossRef]
- Ndiade-Bourobou, D.; Hardy, O.J.; Favreau, B.; Moussavou, H.; Nzengue, E.; Mignot, A.; Bouvet, J.-M. Long-distance seed and pollen dispersal inferred from spatial genetic structure in the very low-density rainforest tree, Baillonella toxisperma Pierre, in Central Africa. Mol. Ecol. 2010, 19, 4949–4962. [Google Scholar] [CrossRef] [PubMed]
- Anjos, D.V.; Leal, L.C.; Jordano, P.; Del-Claro, K. Ants as diaspore removers of non-myrmecochorous plants: A meta-analysis. Oikos 2020, 129, 775–786. [Google Scholar] [CrossRef]
- Campagnoli, M.L.; Christianini, A.V. Temporal consistency in interactions among birds, ants, and plants in a neotropical savanna. Oikos 2021, 2022. [Google Scholar] [CrossRef]
- Ramos, M.; Robles, A.; Castro, J. Efficiency of endozoochorous seed dispersal in six dry-fruited species (Cistaceae): From seed ingestion to early seedling establishment. Plant Ecol. 2006, 185, 97–106. [Google Scholar] [CrossRef]
- Kul’bachko, Y.L.; Didur, O.O.; Loza, I.M.; Pakhomov, O.E.; Bezrodnova, O.V. Environmental aspects of the effect of earthworm (Lumbricidae, Oligochaeta) tropho-metabolic activity on the pH buffering capacity of remediated soil (steppe zone, Ukraine). Biol. Bull. 2015, 42, 899–904. [Google Scholar] [CrossRef] [Green Version]
- Ambarish, C.N.; Sridhar, K.R. Chemical and Microbial Characterization of Feed and Faeces of Two Giant Pill-Millipedes from Forests in the Western Ghats of India. Pedosphere 2016, 26, 861–871. [Google Scholar] [CrossRef]
- Cordeiro, R.S.; Barbosa, J.E.L.; Lima, G.Q.; Barbosa, L.G. Periphytic algae dynamics in lentic ecosystems in the Brazilian semiarid. Braz. J. Biol. 2017, 77, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.L.; Duan, P.F.; Zhang, P.; Li, M. Variations in cyanobacterial and algal communities and soil characteristics under biocrust development under similar environmental conditions. Plant Soil 2018, 429, 241–251. [Google Scholar] [CrossRef]
- Hu, C.X.; Liu, Y.D. Primary succession of algal community structure in desert soil. Acta Bot. Sin. 2003, 45, 917–924. [Google Scholar]
- Maltsev, Y.; Maltseva, I. The influence of forest-forming tree species on diversity and spatial distribution of algae in forest litter. Folia Oecol. 2018, 45, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Maltseva, I.A.; Maltsev, Y.I. Diversity of cyanobacteria and algae in dependence to forest-forming tree species and properties rocks of dump. Int. J. Environ. Sci. Technol. 2020, 545–560. [Google Scholar] [CrossRef]
- Maltsev, Y.I.; Didovich, S.V.; Maltseva, I.A. Seasonal changes in the communities of microorganisms and algae in the litters of tree plantations in the Steppe zone. Eurasian Soil Sci. 2017, 50, 935–942. [Google Scholar] [CrossRef]
- Maltsev, Y.I.; Pakhomov, A.Y.; Maltseva, I.A. Specific features of algal communities in forest litter of forest biogeocenoses of the Steppe zone. Contemp. Probl. Ecol. 2017, 10, 71–76. [Google Scholar] [CrossRef]
- Didur, O.A.; Kulbachko, Y.L.; Maltsev, Y.I.; Konovalenko, T.V. Ecology of soil algae cenoses in Norway maple plantation in the recultivated territory of the Western Donbas (Ukraine). Ukr. J. Ecol. 2018, 8, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, K.A.M.; Ritchie, M.E. Effects of macrophyte species richness on wetland ecosystem functioning and services. Nature 2001, 411, 6838. [Google Scholar] [CrossRef]
- Rossi, F.; Mallet, C.; Portelli, C.; Donnadieu, F.; Bonnemoy, F.; Artigas, J. Stimulation or inhibition: Leaf microbial decomposition in streams subjected to complex chemical contamination. Sci. Total Environ. 2019, 648, 1371–1383. [Google Scholar] [CrossRef]
- David, J.F.; Handa, I.T. The ecology of saprophagous macroarthropods (millipedes, woodlice) in the context of global change. Biol. Rev. 2010, 85, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, M.; Jiménez, J.J.; McKey, D.; Mathieu, J.; Velasquez, E.; Zangerlé, A. Ecosystem engineers in a self-organized soil: A review of concepts and future research questions. Soil Sci. 2016, 181, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Hagner, M.; Romantschuk, M.; Penttinen, O.P.; Egfors, A.; Marchand, C.; Augustsson, A. Assessing toxicity of metal contaminated soil from glassworks sites with a battery of biotests. Sci. Total Environ. 2018, 613–614, 30–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Butenschoen, O.; Barantal, S.; Handa, I.T.; Makkonen, M.; Vos, V.; Aerts, R.; Berg, M.P.; McKie, B.; Van Ruijven, J.; et al. Decomposition of leaf litter mixtures across biomes: The role of litter identity, diversity and soil fauna. J. Ecol. 2020, 108, 2283–2297. [Google Scholar] [CrossRef]
- Brygadyrenko, V.V. Influence of moisture conditions and mineralization of soil solution on structure of litter macrofauna of the deciduous forests of Ukraine steppe zone. Visnyk Dnipropetr. Univ. Biol. Ecol. 2015, 23, 50–65. [Google Scholar] [CrossRef]
- Brygadyrenko, V.V. Influence of tree crown density and density of the herbaceous layer on the structure of litter macrofauna of deciduous forests of Ukraine’s steppe zone. Visnyk Dnipropetr. Univ. Biol. Ecol. 2015, 23, 134–148. [Google Scholar] [CrossRef]
- Svyrydchenko, A.O.; Brygadyrenko, V.V. Trophic preferences of Rossiulus kessleri (Diplopoda, Julidae) for the litter of various tree species. Folia Oecol. 2014, 41, 202–212. [Google Scholar]
- Šustra, V.; Šimek, M.; Faktorová, L.; Macková, J.; Tajovský, K. Release of greenhouse gases from millipedes as related to food, body size, and other factors. Soil Biol. Biochem. 2020, 44, 107765. [Google Scholar] [CrossRef]
- Didur, O.; Kulbachko, Y.; Ovchynnykova, Y.; Pokhylenko, A.; Lykholat, T. Zoogenic mechanisms of ecological rehabilitation of urban soils of the park zone of megapolis. Environ. Res. Eng. Manag. 2019, 75. [Google Scholar] [CrossRef] [Green Version]
- Potapov, A.M.; Beaulieu, F.; Birkhofer, K.; Bluhm, S.L.; Degtyarev, M.I.; Devetter, M.; Goncharov, A.A.; Gongalsky, K.B.; Klarner, B.; Korobushkin, D.I.; et al. Feeding habits and multifunctional classification of soil-associated consumers from protists to vertebrates. Biol. Rev. 2022. [Google Scholar] [CrossRef]
- Shtina, E.A.; Gollerbah, M.M. Ekologiya Pochvennyih Vodorosley; Nauka: Moscow, Russia, 1976; 143p. (In Russian) [Google Scholar]
- Urbasek, F.; Tajovsky, K. The influence of food and temperature on enzymatic-activities of the millipede Glomeris-hexasticha (Diplopoda). Rev. D Ecol. Et De Biol. Du Sol 1991, 28, 155–163. [Google Scholar]
- Sustr, V.; Semanova, S.; Rost-Roszkowska, M.M.; Tajovsky, K.; Sosinka, A.; Kaszuba, F. Enzymatic activities in the digestive tract of spirostreptid and spirobolid millipedes (Diplopoda: Spirostreptida and Spirobolida). Comp. Biochem. Physiol. B Biochem. 2020, 241, 110388. [Google Scholar] [CrossRef] [PubMed]
- Loranger-Merciris, G.; Imbert, D.; Bernhard-Reversat, F.; Lavelle, P.; Ponge, J.F. Litter N-content influences soil millipede abundance, species richness and feeding preferences in a semi-evergreen dry forest of Guadeloupe (Lesser Antilles). Biol. Fertil. Soils 2008, 45, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Seeber, J.; Langel, R.; Meyer, E.; Traugott, M. Dwarf shrub litter as a food source for macro-decomposers in alpine pastureland. Appl. Soil Ecol. 2009, 41, 178–184. [Google Scholar] [CrossRef]
- Gritsan, Y.I. Ekolohichni Osnovy Peretvoryuyuchoho Vplyvu Lisovoyi Roslynnosti na Stepove Seredovyshche; DNU: Dnipro, Ukraine, 2000; 300p. (In Ukranian) [Google Scholar]
- Hryhoriuk, I.P.; Yavorovskyi, P.P.; Lykholat, Y.V. Tekhnolohii Vyroshchuvannia i Biorehuliatsiia Stiikosti Hazonnykh Roslyn u Miskomu Urbanizovanomu Seredovyshchi: Monohrafiia; NULESU: Kyiv, Ukraine, 2014; 223p. (In Ukranian) [Google Scholar]
- Maltsev, Y.I.; Maltseva, I.A.; Solonenko, A.N.; Bren, A.G. Use of soil biota in the assessment of the ecological potential of urban soils. Biosyst. Divers. 2017, 25, 257–262. [Google Scholar] [CrossRef]
- Pokhylenko, A.P.; Didur, O.O.; Kulbachko, Y.L.; Bandura, L.P.; Chernykh, S.A. Influence of saprophages (Isopoda, Diplopoda) on leaf litter decomposition under different levels of humidification and chemical loading. Biosyst. Divers. 2020, 28, 384–389. [Google Scholar] [CrossRef]
- Lykholat, Y.V.; Khromykh, N.A.; Ivan’ko, I.A.; Matyukha, V.L.; Kravets, S.S.; Didur, O.O.; Alexeyeva, A.A.; Shupranova, L.V. Otsinka i prohnoz invaziinosti deiakykh adventyvnykh roslyn za vplyvu klimatychnykh zmin u Stepovomu Prydniprov’i. Biosyst. Divers. 2017, 25, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Stroganova, M.; Prokofieva, T. Urban soils classification for Russian cities of the taiga zone. Eur. Soil Bur.—Res. Rep. No 2001, 153–156. Available online: https://www.researchgate.net/publication/237430707_Urban_soils_classification_for_Russian_cities_of_the_taiga_zone (accessed on 28 April 2022).
- Oliveira, M.P.A.; Bastos-Pereira, R.; Torres, S.H.S.; Pereira, T.S.; Batista, F.M.; Alves, J.P.; Iniesta, L.F.M.; Bouzan, R.S.; Chagas-Jr, A.; Prous, X.; et al. Choosing sampling methods for Chilopoda, Diplopoda and Isopoda (Oniscidea): A case study for ferruginous landscapes in Brazilian Amazonia. Appl. Soil Ecol. 2019, 143, 181–191. [Google Scholar] [CrossRef]
- Martinez, J.M.; Tarazona, R.; Lohr, B.L.; Narvaez, C.A. Measuring the effect of long-term pitfall trapping on the prevalence of epigeal arthropods: A case study in the Pacific Coast of Colombia. Sociobiology 2021, 68, e5928. [Google Scholar] [CrossRef]
- Chernyy, N.G.; Golovach, S.I. Dvuparnonogiye Mnogonozhki Ravninnykh Territoriy Ukrainy; Naukova dumka: Kiev, Russia, 1993; 55p. (In Russian) [Google Scholar]
- Wytwer, J.; Golovatch, S.I.; Penev, L. Variation in millipede (Diplopoda) assemblages in oak woodlands of the Eastern European Plain. Soil Org. 2009, 81, 791–813. [Google Scholar]
- Reip, H.S.; Voigtländer, K. Diplopoda and Chilopoda of Thuringia, Germany. Soil Org. 2009, 81, 635–645. [Google Scholar]
- Gaysina, L.A.; Fazlutdinova, A.I.; Kabirov, R.R. Sovremennyye Metody Vydeleniya i Kultivirovaniya Vodorosley: Uchebnoye Posobiye; BNPU: Ufa, Russia, 2008; 152p. (In Russian) [Google Scholar]
- Neustupa, J.; Eliás, M.; Skaloud, P.; Němcová, Y.; Šejnohová, L. Xylochloris irregularis gen. et sp. nov. (Trebouxiophyceae, Chlorophyta), a novel subaerial coccoid green alga. Phycologia 2011, 50, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J. Cyanoprokaryota. 3rd Part, Heterocytous Genera; Springer Spectrum: Berlin/Heidelberg, Germany, 2013; 1130p. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier GmbH: München, Germany, 2005; 759p. [Google Scholar]
- Frey, W. Syllabus of Plant Families—A. Engler’s Syllabus der Pflanzenfamilien Part 2/1: Photoautotrophic Eukaryotic Algae; Schweizerbart Science Publishers: Stuttgart, Germany, 2015; 324p. [Google Scholar]
- Ettl, H.; Gärtner, G. Syllabus der Boden-, Luft und Flechtenalgen; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1995; 699p. [Google Scholar]
- Ettl, H.; Gärtner, G. Syllabus der Boden-, Luft und Flechtenalgen; Springer Spektrum: Berlin/Heidelberg, Germany, 2014; 733p. [Google Scholar]
- Ettl, H. Xanthophyceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1978; 530p. [Google Scholar]
- Ettl, H. Chlorophyta I (Phytomonadina). In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1983; 807p. [Google Scholar]
- Ettl, H.; Gärtner, G. Chlorophyta II, Tetrasporales, Chlorococcales, Gloeodendrales. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1988; 436p. [Google Scholar]
- Ettl, H.; Gärtner, G. Tetrasporales, Chlorococcales, Gloeodendrales, Chlorophyta II. In Süswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1988; 453p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 2/1, Naviculaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1986; 576p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae. In Subwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany; Stuttgart, Germany, 1988; 654p. [Google Scholar]
- Lewis, L.A.; Flechtner, V.R. Green algae (Chlorophyta) of desert Microbiotic Crusts: Diversity of North American Taxa. Taxon 2002, 51, 443–451. [Google Scholar] [CrossRef]
- Czerwik-Marcinkowska, J.; Mrozińska, T. Algae and Cyanobacteria in caves of the polish. Pol. Bot. J. 2011, 56, 203–243. [Google Scholar]
- Sanmartín, P.; Méndez, A.; Carballeira, R.; López, E. New insights into the growth and diversity of subaerial biofilms colonizing granite-built heritage exposed to UV-A or UV-B radiation plus red LED light. Int. Biodeterior. Biodegrad. 2021, 161, 105225. [Google Scholar] [CrossRef]
- Safiullin, S.Y.; Mansurova, A.R. Ustoychivost Bracteacoccus minor var. desertorum (Friedmann & Ocampo-Paus) k vozdeystviyu vysokikh temperature. Izv. Samar. Nauchnogo Tsentra Ross. Akad. Nauk. 2011, 13, 5. (In Russian) [Google Scholar]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Milovanovic, I.; Misan, A.; Saric, B.; Kos, J.; Mandic, A.; Simeunovic, J.; Kovac, D. Evaluation of protein and lipid content and determination of fatty acid profile in selected species of cyanobacteria. In Proceedings of the 6th Central Europen Congress on Food (CEFood), Novi Sad, Serbia, 23–26 May 2012. [Google Scholar]
- Mata, T.M.; Martinsa, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sust. Energ. Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
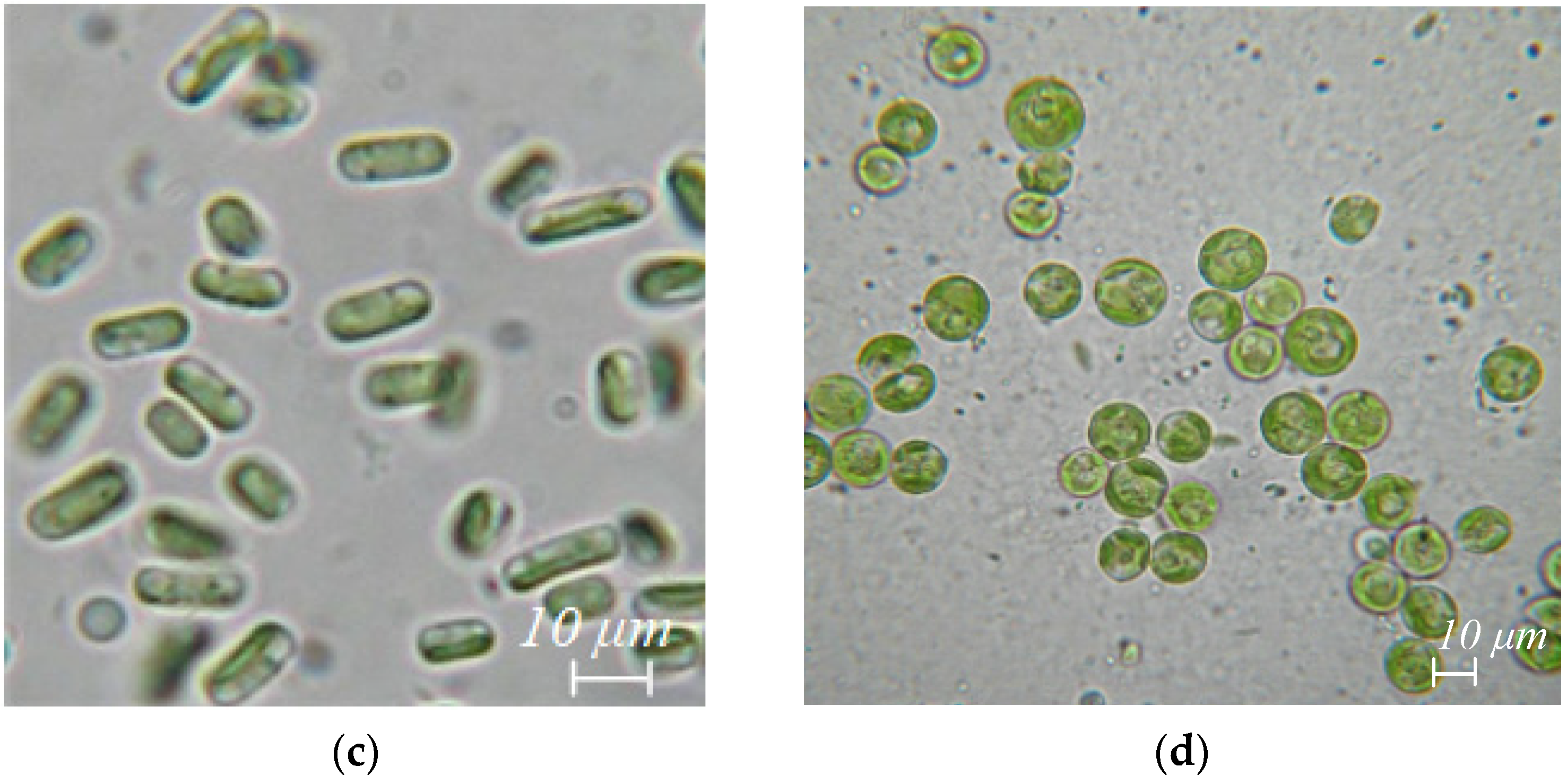
| Taxon | Species | Ecological Life-Form | A. platanoides | A. negundo + R. pseudoacacia + P. deltoides | A. negundo | U. laevis |
|---|---|---|---|---|---|---|
| Cyanobacteria Cyanophyceae | Nostoc punctiforme (Kütz. ex Hariot) Hariot | Cf | + | – | – | – |
| Chlorophyta Chlorophyceae | Bracteacoccus minor (Chodat) Petrová | Ch | + | – | + | – |
| Chlorophyta Chlorophyceae | Mychonastes homosphaera (Skuja) Kalina et Punčoch | Cf | + | + | + | + |
| Chlorophyta Chlorophyceae | Neospongiococcum sp. | – | + | – | – | – |
| Chlorophyta Chlorophyceae | Chlamydomonas sp. | – | + | – | – | – |
| Chlorophyta Trebouxiophyceae | Chlorella vulgaris Beij. | Ch | + | + | + | + |
| Chlorophyta Trebouxiophyceae | Stichococcus bacillaris Nägeli | Cf | + | – | + | + |
| Chlorophyta Trebouxiophyceae | Pseudococcomyxa simplex (Mainx) Fott | Ch | + | – | – | – |
| Chlorophyta Trebouxiophyceae | Desmococcus olivaceus (Pers. ex Ach.) I.R. Laundon | Ch | + | + | + | – |
| Chlorophyta Trebouxiophyceae | Trebouxia spp. | – | + | – | – | – |
| Streptophyta Klebsormidiophyceae | Klebsormidium flaccidum (Kütz.) P.C. Silva et al. | H | + | – | + | – |
| Heterokontophyta (Ochrophyta) Xanthophyceae | Nephrodiella phaseolus Pascher | Ch | + | – | – | – |
| Heterokontophyta (Ochrophyta) Bacillariophyceae | Navicula pelliculosa (Breb.) Hilse | B | + | – | – | – |
| Heterokontophyta (Ochrophyta) Eustigmatophyceae | Vischeria magna (J.B. Petersen) Hibberd | Ch | + | + | + | – |
| Algae Species | A. platanoides | A. negundo + R. pseudoacacia + P. deltoides | A. negundo | U. laevis |
|---|---|---|---|---|
| Chlorella vulgaris | – | + | – | + |
| Mychonastes homosphaera | – | + | + | – |
| Desmococcus olivaceus | – | + | – | – |
| Vischeria magna | – | + | – | – |
| Stichococcus bacillaris | + | – | – | – |
| Variant of Experiment | A. platanoides | A. negundo + R. pseudoacacia + P. deltoides | A. negundo | U. laevis |
|---|---|---|---|---|
| Gut wash | ||||
| Bracteacoccusminor | + | – | – | + |
| Chlorella vulgaris | + | – | + | + |
| Mychonasteshomosphaera | – | – | – | + |
| Desmococcus olivaceus | – | – | + | – |
| Stichococcus bacillaris | – | – | + | – |
| Klebsormidium flaccidum | – | + | + | – |
| Vischeriamagna | + | + | – | – |
| Occurrence from excretion | ||||
| Chlorella vulgaris | + | + | + | + |
| Mychonasteshomosphaera | – | – | – | + |
| Desmococcusolivaceus | – | – | + | – |
| Vischeriamagna | – | + | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakhomov, O.; Pokhylenko, A.; Maltseva, I.; Kulbachko, Y. Participation of Rossiulus kessleri (Diplopoda, Julida) in the Formation of Algae Assemblages of Urbanized Territories. Diversity 2022, 14, 508. https://doi.org/10.3390/d14070508
Pakhomov O, Pokhylenko A, Maltseva I, Kulbachko Y. Participation of Rossiulus kessleri (Diplopoda, Julida) in the Formation of Algae Assemblages of Urbanized Territories. Diversity. 2022; 14(7):508. https://doi.org/10.3390/d14070508
Chicago/Turabian StylePakhomov, Olexandr, Alevtyna Pokhylenko, Iryna Maltseva, and Yurii Kulbachko. 2022. "Participation of Rossiulus kessleri (Diplopoda, Julida) in the Formation of Algae Assemblages of Urbanized Territories" Diversity 14, no. 7: 508. https://doi.org/10.3390/d14070508