
DNA Identification and Diversity of the Vector Mosquitoes Culex pipiens s.s. and Culex torrentium in Belgium (Diptera: Culicidae)


 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction and COI Amplification
2.3. Fragment-Size Analyses
2.4. COI Data Analyses
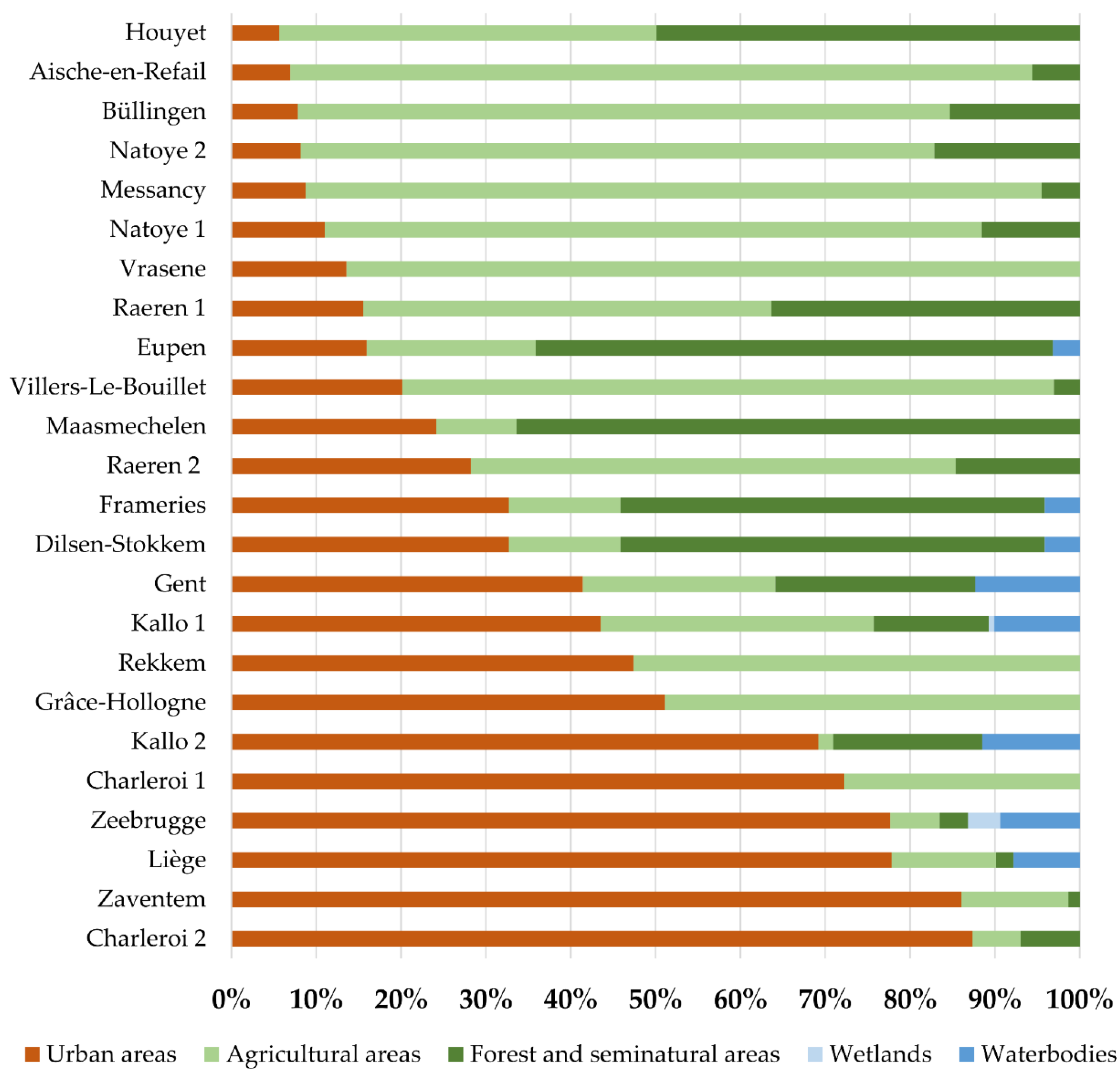
2.5. Habitat Characterization: Land-Cover Classes
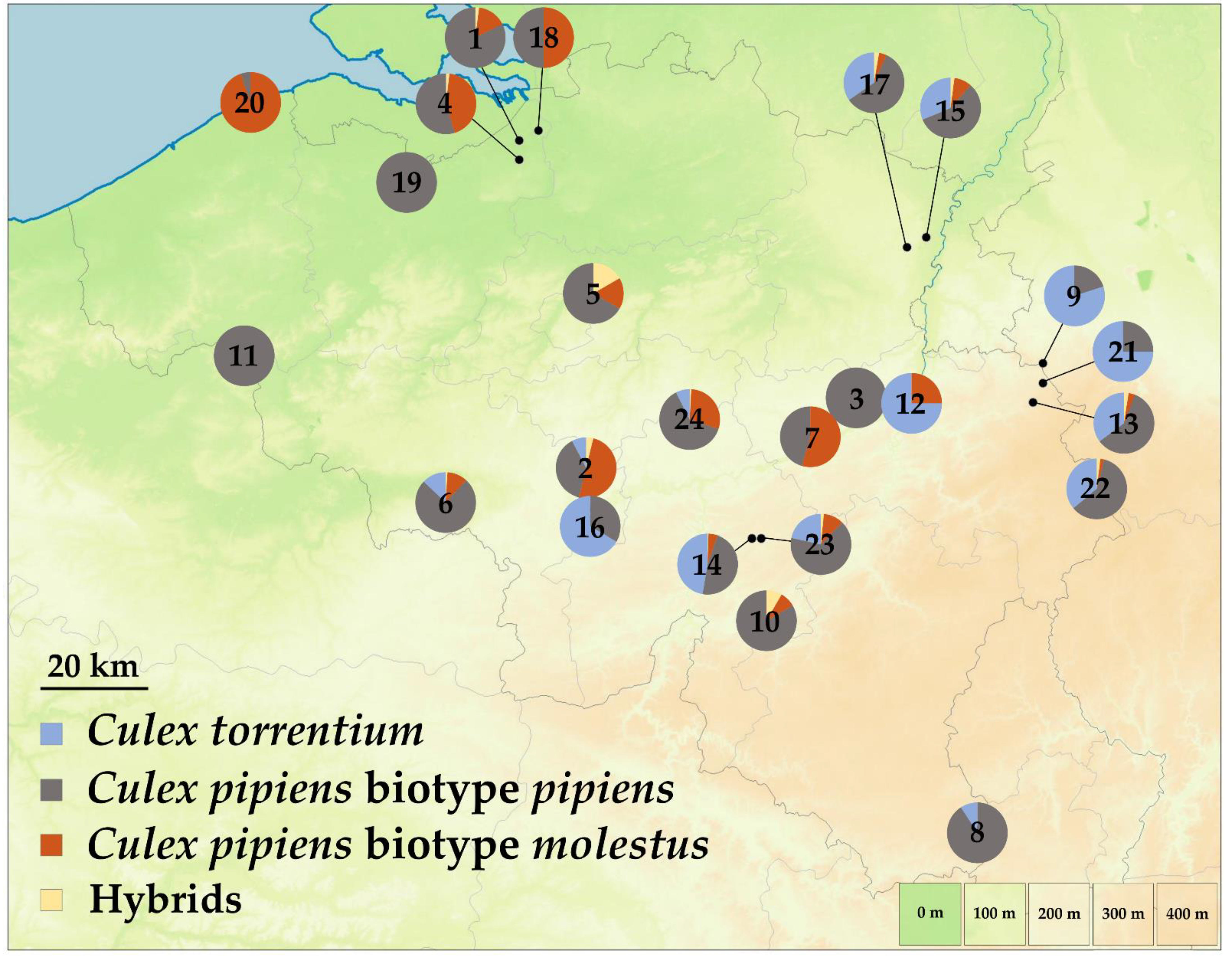
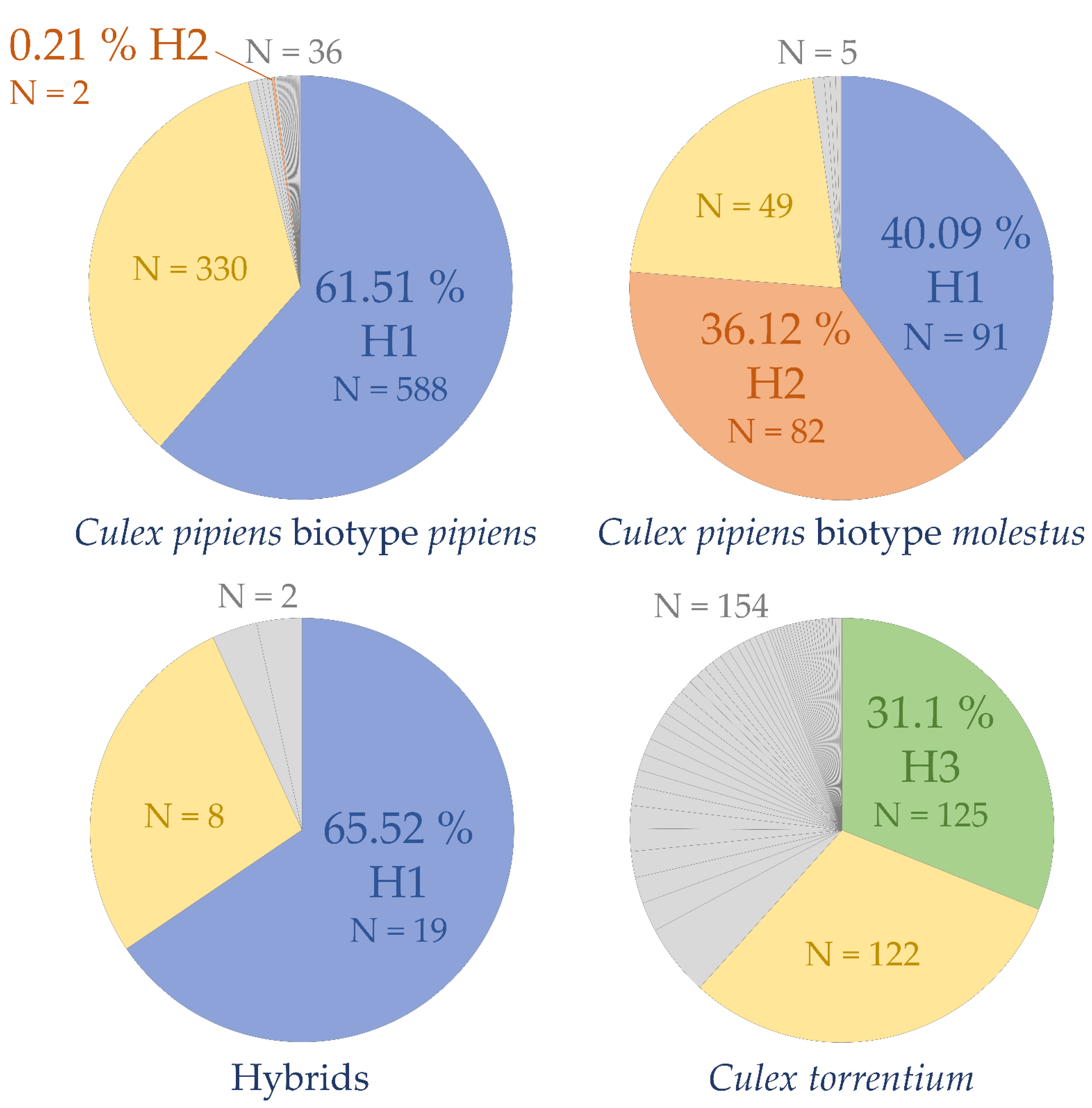
3. Results
4. Discussion
4.1. Species Occurrence in Belgium
4.2. COI Haplotype Composition and Genetic Variability
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ECDC. Culex pipiens Factsheet. 2020. Available online: https://www.ecdc.europa.eu/en/all-topics-z/disease-vectors/facts/mosquito-factsheets/culex-pipiens-factsheet-experts (accessed on 16 February 2022).
- Scholte, E.; Ibáñez-Justicia, A.; Stroo, A.; Zeeuw, J.D.; Hartog, W.D.; Reusken, C. Mosquito collections on incoming intercontinental flights at Schiphol International airport, the Netherlands, 2010–2011. J. Eur. Mosq. Control Assoc. 2014, 32, 17–21. [Google Scholar]
- Shaikevich, E.V.; Vinogradova, E.B.; Bouattour, A.; Gouveia de Almeida, A.P. Genetic diversity of Culex pipiens mosquitoes in distinct populations from Europe: Contribution of Cx. quinquefasciatus in Mediterranean populations. Parasites Vectors 2016, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer: Heidelberg, Germany; New York, NY, USA, 2010. [Google Scholar]
- Miller, B.R.; Crabtree, M.B.; Savage, H.M. Phylogeny of fourteen Culex mosquito species, including the Culex pipiens complex, inferred from the internal transcribed spacers of ribosomal DNA. Insect Mol. Biol. 1996, 5, 93–107. [Google Scholar] [CrossRef]
- Harbach, R.E. Culex pipiens: Species versus species complex taxonomic history and perspective. J. Am. Mosq. Control Assoc. 2012, 28, 10–23. [Google Scholar] [CrossRef]
- Hesson, J.C.; Rettich, F.; Merdić, E.; Vignjević, G.; Östman, Ö.; Schäfer, M.; Schaffner, F.; Foussadier, R.; Besnard, G.; Medlock, J.; et al. The arbovirus vector Culex torrentium is more prevalent than Culex pipiens in northern and central Europe. Med. Vet. Entomol. 2014, 28, 179–186. [Google Scholar] [CrossRef]
- Dekoninck, W.; Hendrickx, F.; Versteirt, V.; Coosemans, M.; De Clercq, E.M.; Hendrickx, G.; Hance, T.; Grootaert, P. Changes in species richness and spatial distribution of mosquitoes (Diptera: Culicidae) inferred from museum specimen records and a recent inventory: A case study from Belgium suggests recent expanded distribution of arbovirus and malaria vectors. J. Med. Entomol. 2013, 50, 237–243. [Google Scholar] [CrossRef]
- Versteirt, V.; Boyer, S.; Damiens, D.; De Clercq, E.; Dekoninck, W.; Ducheyne, E.; Grootaert, P.; Garros, C.; Hance, T.; Hendrickx, G. Nationwide inventory of mosquito biodiversity (Diptera: Culicidae) in Belgium, Europe. Bull. Èntomol. Res. 2013, 103, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deblauwe, I.; Sohier, C.; Schaffner, F.; Rakotoarivony, L.M.; Coosemans, M. Implementation of surveillance of invasive mosquitoes in Belgium according to the ECDC guidelines. Parasites Vectors 2014, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Deblauwe, I.; De Wolf, K.; Smitz, N.; Vanslembrouck, A.; Schneider, A.; De Witte, J.; Verlé, I.; Dekoninck, W.; De Meyer, M.; Backeljau, T.; et al. Monitoring of Exotic Mosquitoes in Belgium (MEMO): Final Report Phase 7 Part 1: MEMO Results; Institute of Tropical Medicine: Antwerp, Belgium, 2020. [Google Scholar]
- Boukraa, S.; Dekoninck, W.; Versteirt, V.; Schaffner, F.; Coosemans, M.; Haubruge, E.; Francis, F. Updated checklist of the mosquitoes (Diptera: Culicidae) of Belgium. J. Vector Ecol. 2015, 40, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Boukraa, S.; de La Grandiere, M.A.; Bawin, T.; Raharimalala, F.N.; Zimmer, J.Y.; Haubruge, E. Diversity and ecology survey of mosquitoes potential vectors in Belgian equestrian farms: A threat prevention of mosquito-borne equine arboviruses. Prev. Vet. Med. 2016, 124, 58–68. [Google Scholar] [CrossRef]
- Harbach, R.E.; Harrison, B.A.; Gad, A.M. Culex (Culex) molestus Forskål (Diptera: Culicidae): Neotype designation, description, variation, and taxonomic status. Proc. Entomol. Soc. Wash. 1984, 86, 521–542. [Google Scholar]
- Razygraev, A.V.; Sulesco, T.M. The Use of the Bayes Factor for identification of Culex pipiens and C. torrentium (Diptera: Culicidae) based on morphometric wing characters. Èntomol. Rev. 2020, 100, 220–227. [Google Scholar] [CrossRef]
- Byrne, K.; Nichols, R.A. Culex pipiens in London Underground tunnels: Differentiation between surface and subterranean populations. Heredity 1999, 82, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Lundström, J. Mosquito-borne viruses in western Europe: A review. J. Vector Ecol. 1999, 24, 1–39. [Google Scholar] [PubMed]
- Vinogradova, E.B. Culex Pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and control; Pensoft: Sofia, Bulgaria, 2000. [Google Scholar]
- Rudolf, M.; Czajka, C.; Börstler, J.; Melaun, C.; Jöst, H.; von Thien, H.; Badusche, M.; Becker, N.; Schmidt-Chanasit, J.; Krüger, A.; et al. First nationwide surveillance of Culex pipiens Complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS ONE 2013, 8, e71832. [Google Scholar] [CrossRef]
- Haba, Y.; McBride, L. Origin and status of Culex pipiens mosquito ecotypes. Curr. Biol. 2022, 32, R237–R246. [Google Scholar] [CrossRef]
- Becker, N.; Jöst, A.; Weitzel, T. The Culex pipiens complex in Europe. J. Am. Mosq. Control Assoc. 2012, 28, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, A.A.; Masri, R.A.; Khrabrova, N.V.; Sibataev, A.K.; Fritz, M.L.; Sharakhova, M.V. Genomic differentiation and intercontinental population structure of mosquito vectors Culex pipiens pipiens and Culex pipiens molestus. Sci. Rep. 2020, 10, 7504. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.L.; Walker, E.D.; Miller, J.R.; Severson, D.W.; Dworkin, I. Divergent host preferences of above- and below-ground Culex pipiens mosquitoes and their hybrid offspring. Med. Vet. Entomol. 2015, 29, 115–123. [Google Scholar] [CrossRef]
- Osório, H.C.; Zé-Zé, L.; Amaro, F.; Nunes, A.; Alves, M.J. Sympatric occurrence of Culex pipiens (Diptera, Culicidae) biotypes pipiens, molestus and their hybrids in Portugal, Western Europe: Feeding patterns and habitat determinants. Med. Vet. Entomol. 2014, 28, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Sousa, C.A.; Vicente, J.L.; Pinho, L.; Calderón, I.; Arez, E.; Almeida, A.P.; Donnelly, M.J.; Pinto, J. Feeding patterns of molestus and pipiens forms of Culex pipiens (Diptera: Culicidae) in a region of high hybridization. Parasites Vectors 2013, 6, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, V.A.; Hernández-Triana, L.M.; Medlock, J.M.; Fooks, A.R.; Carpenter, S.; Johnson, N. The role of Culex pipiens L. (Diptera: Culicidae) in virus transmission in Europe. Int. J. Environ. Res. Public Health 2018, 15, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, D.M.; Keyghobadi, N.; Malcolm, C.A.; Mehmet, C.; Schaffner, F.; Mogi, M.; Fleischer, R.C.; Wilkerson, R.C. Emerging Vectors in the Culex pipiens Complex. Science 2004, 303, 1535–1538. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.; Kioulos, E.; Papa, A.; Almeida, A.P.G.; Vontas, J.; Pinto, J. Distribution and hybridization of Culex pipiens forms in Greece during the West Nile virus outbreak of 2010. Infect. Genet. Evol. 2013, 16, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Chin, P.A.; Kramer, L.D. The effect of hybridization of Culex pipiens complex mosquitoes on transmission of West Nile virus. Parasites Vectors 2013, 6, 305. [Google Scholar] [CrossRef] [Green Version]
- Hesson, J.C.; Verner-Carlsson, J.; Larsson, A.; Ahmed, R.; Lundkvist, Å.; Lundström, J.O. Culex torrentium mosquito role as major enzootic vector defined by rate of Sindbis virus infection, Sweden, 2009. Emerg. Infect. Dis. 2015, 21, 875. [Google Scholar] [CrossRef] [PubMed]
- Leggewie, M.; Badusche, M.; Rudolf, M.; Jansen, S.; Börstler, J.; Krumkamp, R.; Huber, K.; Krüger, A.; Schmidt-Chanasit, J.; Tannich, E.; et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health 2016, 2, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A potent vector for the transmission of west nile virus in central Europe. Viruses 2019, 11, 492. [Google Scholar] [CrossRef] [Green Version]
- ECDC. Surveillance and Disease Data for West Nile Virus Infections. 2020. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data (accessed on 21 February 2022).
- Weitzel, T.; Braun, K.; Collado, A.; Jöst, A.; Becker, N. Distribution and frequency of Culex pipiens and Culex torrentium (Culicidae) in Europe and diagnostic allozyme markers. Eur. Mosq. Bull. 2011, 29, 22–37. [Google Scholar]
- Calistri, P.; Giovannini, A.; Hubalek, Z.; Ionescu, A.; Monaco, F.; Savini, G.; Lelli, R. Epidemiology of West Nile in Europe and in the Mediterranean Basin. Open Virol. J. 2010, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- García-Carrasco, J.M.; Muñoz, A.R.; Olivero, J.; Segura, M.; Real, R. Predicting the spatio-temporal spread of West Nile virus in Europe. PLoS Negl. Trop. Dis. 2021, 15, e0009022. [Google Scholar] [CrossRef] [PubMed]
- Gunay, F.; Picard, M.; Robert, V. MosKeyTool, An Interactive Identification Key for Mosquitoes of Euro-Mediterranean. 2018. Available online: http://medilabsecure.com/moskeytool (accessed on 5 August 2020).
- Wickham, H.; François, R.; Henry, L.; Müller, K. A Grammar of Data Manipulation dplyr 1.0.8. 2020. Available online: https://dplyr.tidyverse.org/index.html (accessed on 1 April 2022).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hebert, P.; Cywinska, A.; Ball, S.; DeWaard, J. Biological identifications through DNA barcodes. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenback, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Èntomol. Soc. Am. 1994, 87, 653–701. [Google Scholar] [CrossRef]
- Ibáñez-Justicia, A.; Smitz, N.; den Hartog, W.; van de Vossenberg, B.; De Wolf, K.; Deblauwe, I.; Van Bortel, W.; Jacobs, F.; Vaux, A.G.C.; Medlock, J.M.; et al. Detection of exotic mosquito species (Diptera: Culicidae) at international airports in Europe. Int. J. Environ. Res. Public Health 2020, 17, 3450. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Fonseca, D.M. Rapid assays for identification of members of the Culex (Culex) pipiens complex, their hybrids, and other sibling species (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 2004, 70, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahnck, C.M.; Fonseca, D.M. Rapid assay to identify the two genetic forms of Culex (Culex) pipiens L. (Diptera: Culicidae) and hybrid populations. Am. J. Trop. Med. Hyg. 2006, 75, 251–255. [Google Scholar] [CrossRef] [Green Version]
- De Wolf, K.; Vanderheyden, A.; Deblauwe, I.; Smitz, N.; Gombeer, S.; Vanslembrouck, A.; Meganck, K.; Dekoninck, W.; DE Meyer, M.; Backeljau, T.; et al. First record of the West Nile virus bridge vector Culex modestus Ficalbi (Diptera: Culicidae) in Belgium, validated by DNA barcoding. Zootaxa 2021, 4920, 131–139. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Gomes, B.; Sousa, C.A.; Novo, M.T.; Freitas, F.B.; Alves, R.; Côrte-Real, A.R.; Salgueiro, P.; Donnelly, M.J.; Almeida, A.P.; Pinto, J. Asymmetric introgression between sympatric molestus and pipiens forms of Culex pipiens (Diptera: Culicidae) in the Comporta region, Portugal. BMC Evol. Biol. 2009, 9, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versteirt, V.; De Clercq, E.; Dekoninck, W.; Damiens, D.; Ayrinhac, A.; Jacobs, F.; Bortel, W.V. MODIRISK: Mosquito Vectors of Disease: Spatial Biodiversity, Drivers of Change, and Risk. Antwerp, Belgium. 2011. Available online: https://www.belspo.be/belspo/SSD/science/Reports/MODIRISK_Phase 1 Summary.pdf (accessed on 31 January 2017).
- Di Luca, M.; Toma, L.; Boccolini, D.; Severini, F.; La Rosa, G.; Minelli, G.; Bongiorno, G.; Montarsi, F.; Arnoldi, D.; Capelli, G.; et al. Ecological Distribution and CQ11 Genetic Structure of Culex pipiens Complex (Diptera: Culicidae) in Italy. PLoS ONE 2016, 11, e0146476. [Google Scholar] [CrossRef]
- Zittra, C.; Flechl, E.; Kothmayer, M.; Vitecek, S.; Rossiter, H.; Zechmeister, T.; Fuehrer, H.-P. Ecological characterization and molecular differentiation of Culex pipiens complex taxa and Culex torrentium in eastern Austria. Parasites Vectors 2016, 9, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Hamer, G.L.; Molaei, G.; Walker, E.D.; Goldberg, T.L.; Kitron, U.D.; Andreadis, T.G. Genetic variation associated with mammalian feeding in Culex pipiens from a West Nile virus epidemic region in Chicago, Illinois. Vector-Borne Zoonotic Dis. 2009, 9, 637–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werblow, A.; Klimpel, S.; Bolius, S.; Dorresteijn, A.W.C.; Sauer, J.; Melaun, C. Population structure and distribution patterns of the sibling mosquito species Culex pipiens and Culex torrentium (Diptera: Culicidae) reveal different evolutionary paths. PLoS ONE 2014, 9, e102158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werblow, A.; Bolius, S.; Dorresteijn, A.W.C.; Melaun, C.; Klimpel, S. Diversity of Culex torrentium Martini, 1925—A potential vector of arboviruses and filaria in Europe. Parasitol. Res. 2013, 112, 2495–2501. [Google Scholar] [CrossRef]
- Fedorova, M.V.; Shaikevich, E.V. Morphological and molecular-genetic distinctions between adult mosquitoes Culex torrentium Martini and C. pipiens Linnaeus (Diptera, Culicidae) from Moscow Province. Èntomol. Rev. 2007, 87, 127–135. [Google Scholar] [CrossRef]
- Guillemaud, T.; Pasteur, N.; Rousset, F. Contrasting levels of variability between cytoplasmic genomes and incompatibility types in the mosquito Culex pipiens. Proc. R. Soc. London Ser. B Biol. Sci. 1997, 264, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Koosha, M.; Oshaghi, M.A.; Sedaghat, M.M.; Vatandoost, H.; Azari-Hamidian, S.; Abai, M.R.; Hanafi-Bojd, A.A.; Mohtarami, F. Sequence analysis of mtDNA COI barcode region revealed three haplotypes within Culex pipiens assemblage. Exp. Parasitol. 2017, 181, 102–110. [Google Scholar] [CrossRef]
- Cariou, M.; Duret, L.; Charlat, S. The global impact of Wolbachia on mitochondrial diversity and evolution. J. Evol. Biol. 2017, 30, 2204–2210. [Google Scholar] [CrossRef] [Green Version]
- Hurst, G.D.D.; Jiggins, F.M. Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: The effects of inherited symbionts. Proc. R. Soc. B Boil. Sci. 2005, 272, 1525–1534. [Google Scholar] [CrossRef]
- Bergman, A.; Hesson, J.C. Wolbachia prevalence in the vector species Culex pipiens and Culex torrentium in a Sindbis virus-endemic region of Sweden. Parasites Vectors 2021, 14, 428. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Lagnel, J.; Raymond, M.; Bourtzis, K.; Fort, P.; Weill, M. Transposable element polymorphism of Wolbachia in the mosquito Culex pipiens: Evidence of genetic diversity, superinfection and recombination. Mol. Ecol. 2005, 14, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E.B.; Fedorova, M.V.; Shaikevich, E.V. Endosymbiotic bacterium Wolbachia pipientis in synanthropic populations of the mosquito Culex pipiens pipiens L. (Diptera, Culicidae). Dokl. Biol. Sci. 2003, 389, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Leggewie, M.; Krumkamp, R.; Badusche, M.; Heitmann, A.; Jansen, S.; Schmidt-Chanasit, J.; Tannich, E.; Becker, S.C. Culex torrentium mosquitoes from Germany are negative for Wolbachia. Med. Vet. Entomol. 2018, 32, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaikevich, E. PCR-RFLP of the COI gene reliably differentiates Cx. pipiens, Cx. pipiens f. molestus and Cx. torrentium of the Pipiens Complex. Eur. Mosq. Bull. 2007, 23, 25–30. [Google Scholar]
- Gomes, B.; Wilding, C.S.; Weetman, D.; Sousa, C.A.; Novo, M.T.; Savage, H.M.; Almeida, A.P.G.; Pinto, J.; Donnelly, M.J. Limited genomic divergence between intraspecific forms of Culex pipiens under different ecological pressures. BMC Evol. Biol. 2015, 15, 197. [Google Scholar] [CrossRef] [Green Version]
- Kothera, L.; Godsey, M.; Mutebi, J.P.; Savage, H.M. A comparison of aboveground and belowground populations of Culex pipiens (Diptera: Culicidae) mosquitoes in Chicago, Illinois, and New York City, New York, using microsatellites. J. Med. Entomol. 2010, 47, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Trocke, S.; Sim, C. Comparative studies of stenogamous behaviour in the mosquito Culex pipiens complex. Med. Vet. Entomol. 2018, 32, 427–435. [Google Scholar] [CrossRef]
- Bourguet, D.; Fonseca, D.; Vourch, G.; Dubois, M.; Chandre, F.; Severini, C.; Raymond, M. The acetylcholinesterase gene Ace: A diagnostic marker for the pipiens and quinquefasciatus forms of the Culex pipiens complex. J. Am. Mosq. Control Assoc. 1998, 14, 390–396. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| FST | Hybrids | Cx. pipiens Biotype molestus | Cx. pipiens Biotype pipiens |
|---|---|---|---|
| Hybrids | 0 | - | - |
| Cx. pipiens biotype molestus | 0.2172 * | 0 | - |
| Cx. pipiens biotype pipiens | 0.0164 | 0.4737 * | 0 |
| Hybrids | Cx. pipiens Biotype molestus | Cx. pipiens Biotype pipiens | Cx. torrentium | |
|---|---|---|---|---|
| NCOI | 29 | 227 | 956 | 401 |
| NHap | 3 | 6 | 24 | 50 |
| NUnsharedHap | 1 | 3 | 20 | 50 |
| S | 3 | 8 | 19 | 31 |
| k | 0.2069 ± 0.2606 | 0.5458 ± 0.4522 | 0.0703 ± 0.1418 | 0.8990 ± 0.6275 |
| Pi | 0.0006 ± 0.0008 | 0.0014 ± 0.0012 | 0.0002 ± 0.0004 | 0.0024 ± 0.0018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanderheyden, A.; Smitz, N.; De Wolf, K.; Deblauwe, I.; Dekoninck, W.; Meganck, K.; Gombeer, S.; Vanslembrouck, A.; De Witte, J.; Schneider, A.; et al. DNA Identification and Diversity of the Vector Mosquitoes Culex pipiens s.s. and Culex torrentium in Belgium (Diptera: Culicidae). Diversity 2022, 14, 486. https://doi.org/10.3390/d14060486
Vanderheyden A, Smitz N, De Wolf K, Deblauwe I, Dekoninck W, Meganck K, Gombeer S, Vanslembrouck A, De Witte J, Schneider A, et al. DNA Identification and Diversity of the Vector Mosquitoes Culex pipiens s.s. and Culex torrentium in Belgium (Diptera: Culicidae). Diversity. 2022; 14(6):486. https://doi.org/10.3390/d14060486
Chicago/Turabian StyleVanderheyden, Ann, Nathalie Smitz, Katrien De Wolf, Isra Deblauwe, Wouter Dekoninck, Kenny Meganck, Sophie Gombeer, Adwine Vanslembrouck, Jacobus De Witte, Anna Schneider, and et al. 2022. "DNA Identification and Diversity of the Vector Mosquitoes Culex pipiens s.s. and Culex torrentium in Belgium (Diptera: Culicidae)" Diversity 14, no. 6: 486. https://doi.org/10.3390/d14060486