
Dam Construction Impacts Fish Biodiversity in a Subtropical River Network, China


Abstract
:1. Introduction
2. Material and Methods
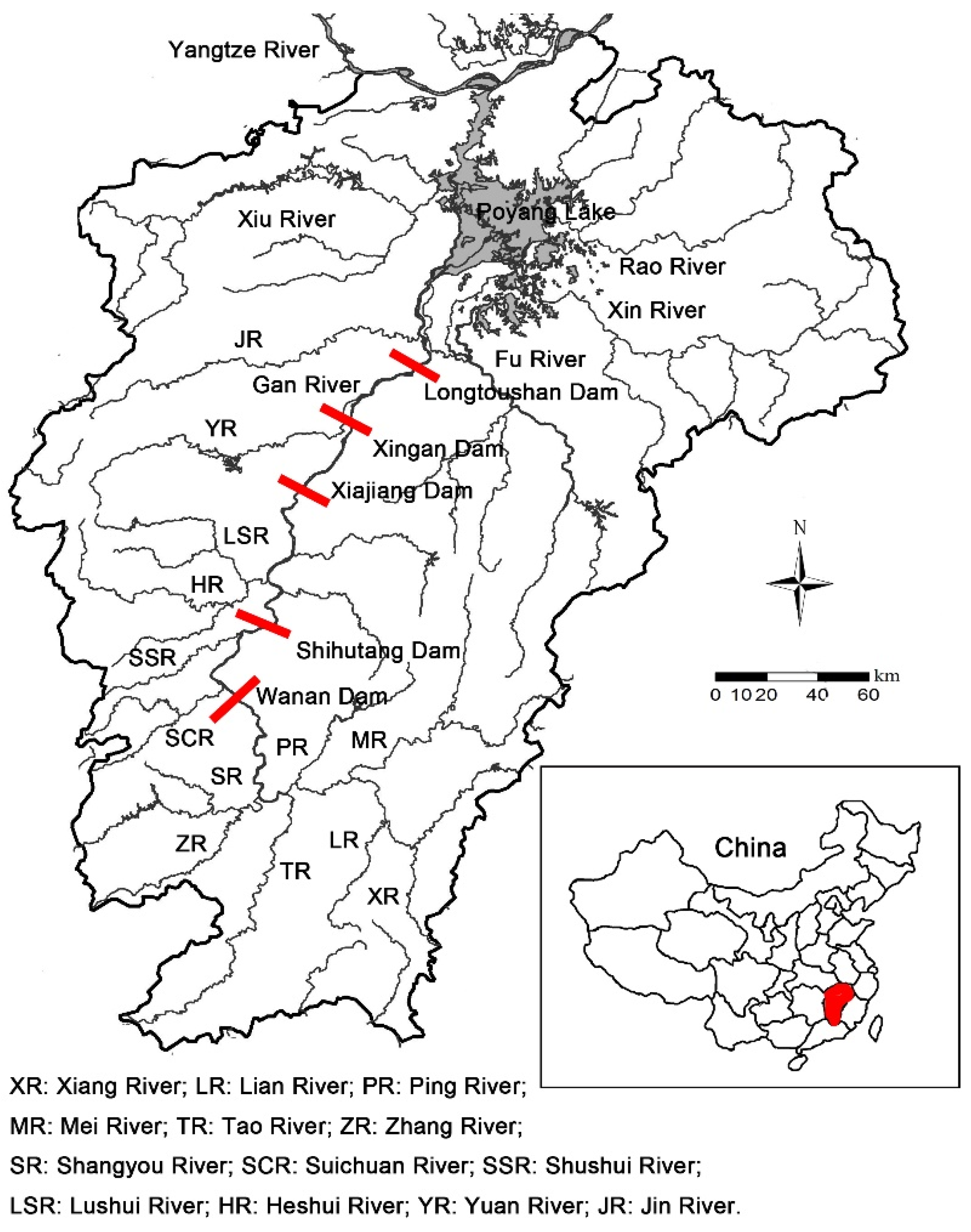
2.1. Study Area
2.2. Data Collection
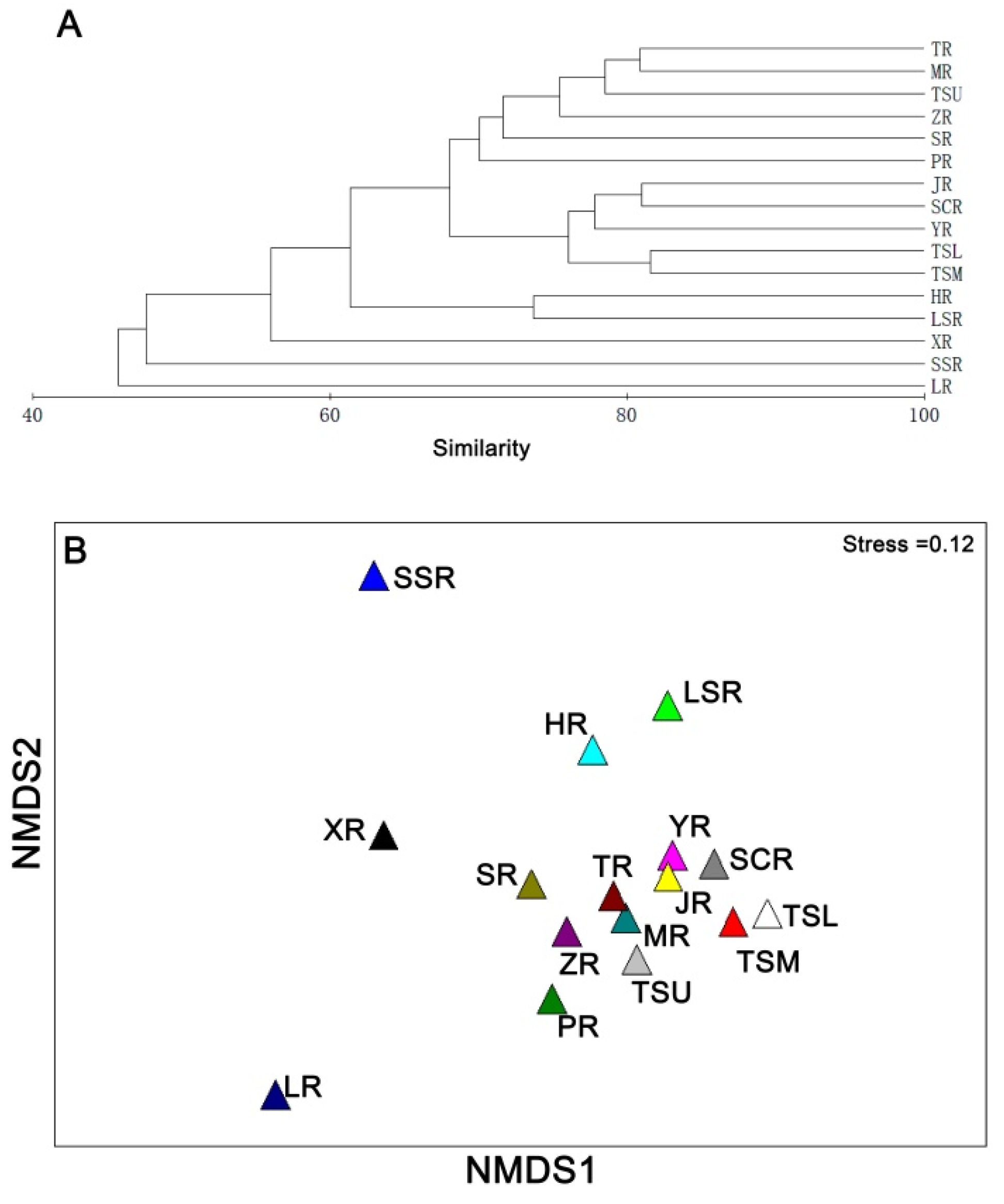
2.3. Data Analysis
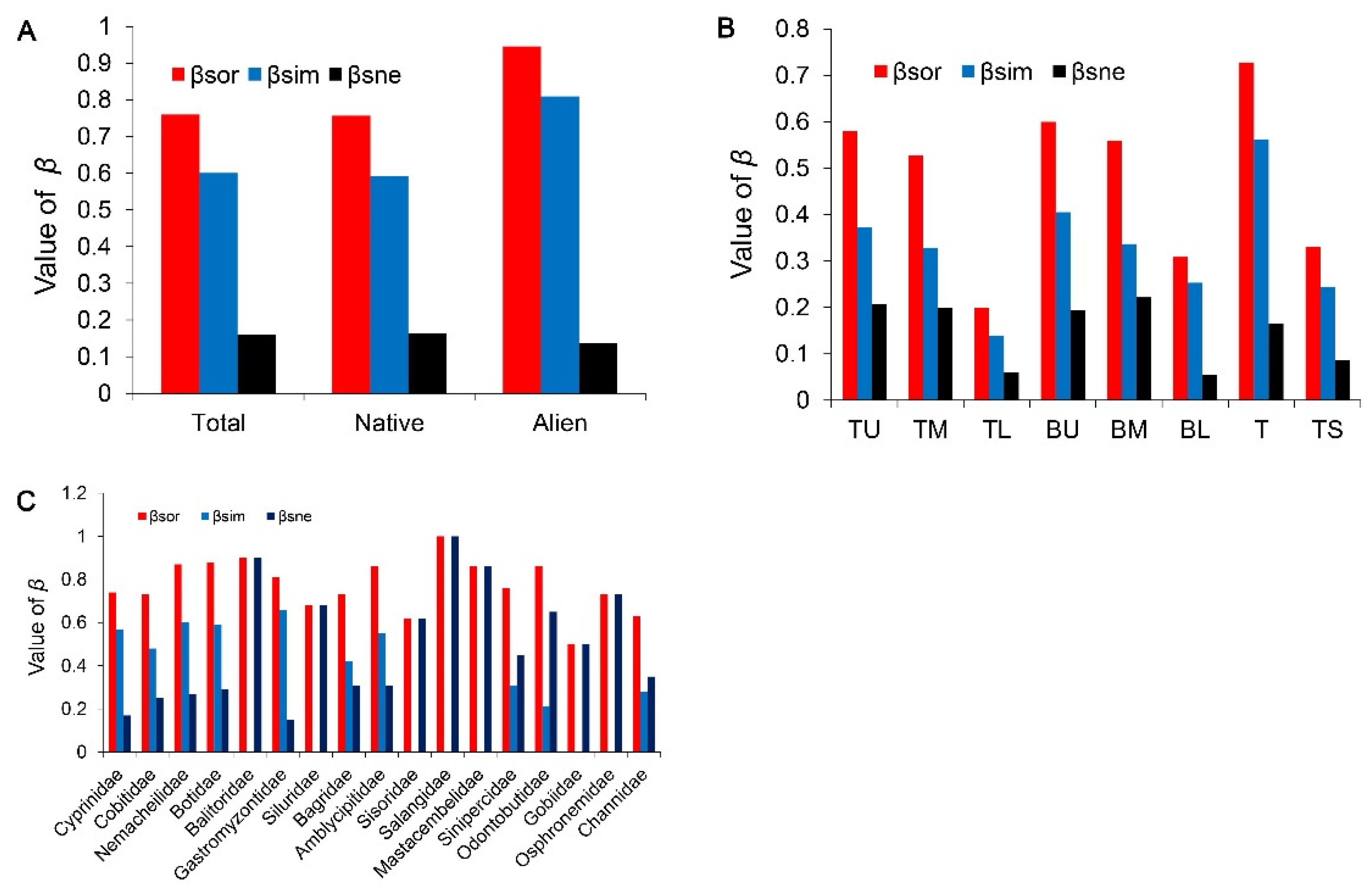
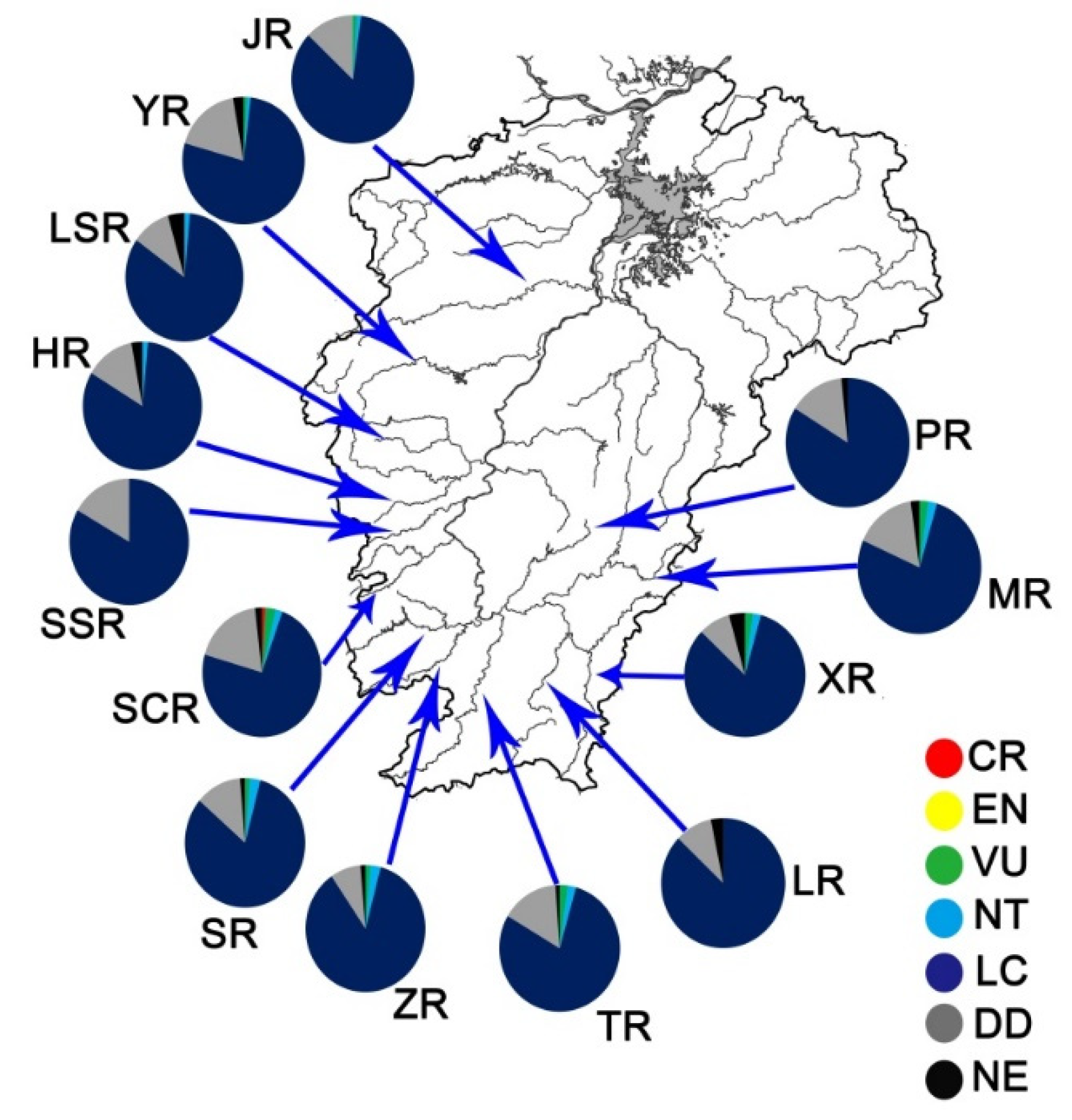
3. Results
3.1. Changes in Species Diversity
3.2. Threatened Status of Fish
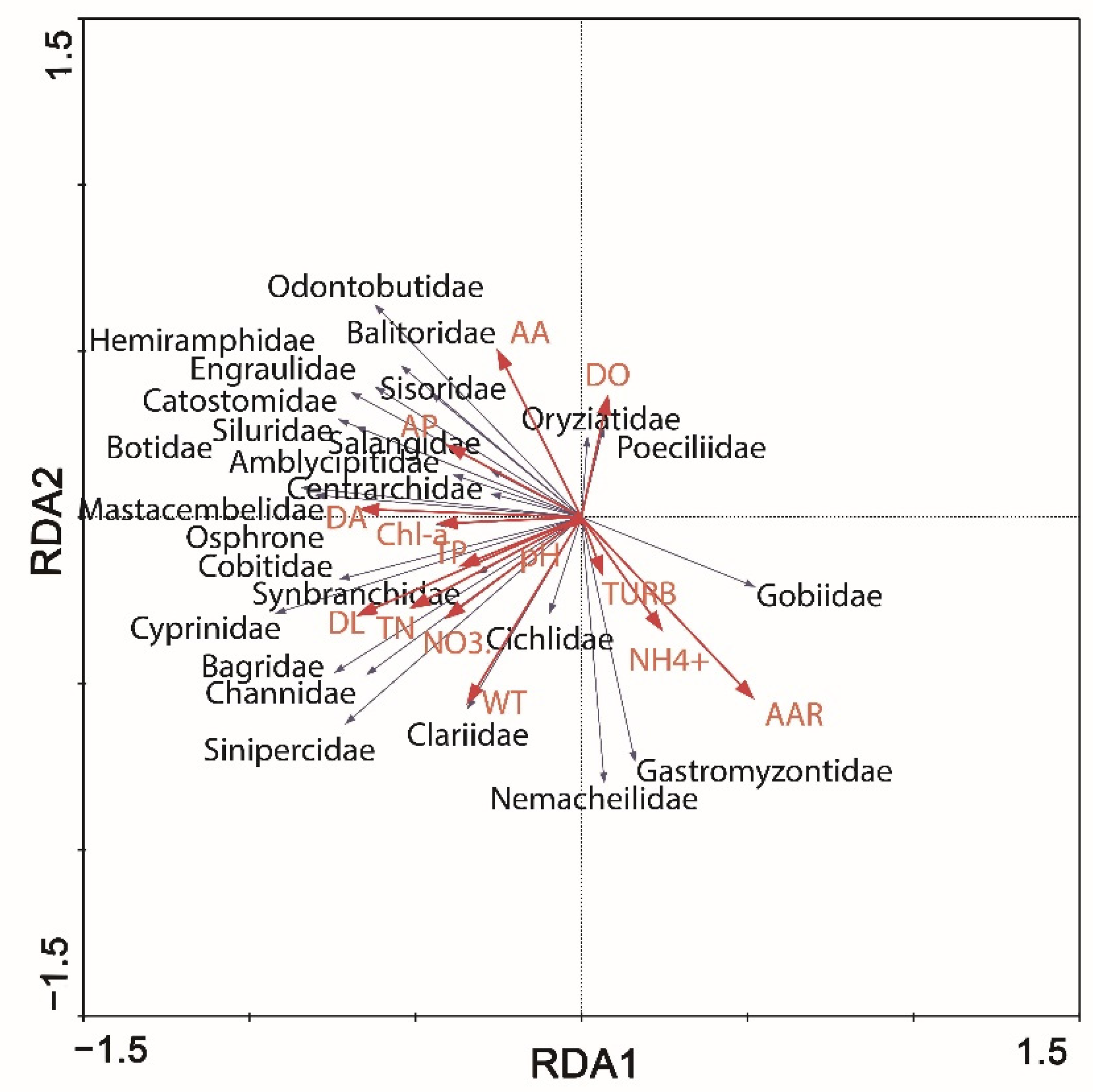
3.3. Environmental Drivers of Fish Diversity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Couto, T.B.; Olden, J.D. Global proliferation of small hydropower plants—science and policy. Front. Ecol. Environ. 2018, 16, 91–100. [Google Scholar] [CrossRef]
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the world’s free-flowing rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tickner, D.; Opperman, J.J.; Abell, R.; Acreman, M.; Arthington, A.H.; Bunn, S.E.; Cooke, S.J.; Dalton, J.; Darwall, W.; Edwards, G.; et al. Bending the Curve of Global Freshwater Biodiversity Loss: An Emergency Recovery Plan. BioScience 2020, 70, 330–342. [Google Scholar] [CrossRef]
- Bunn, S.E.; Arthington, A.H. Basic Principles and Ecological Consequences of Altered Flow Regimes for Aquatic Biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef] [Green Version]
- Olden, J.D.; Naiman, R.J. Incorporating thermal regimes into environmental flows assessments: Modifying dam operations to restore freshwater ecosystem integrity. Freshw. Biol. 2010, 55, 86–107. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Bonecker, C.C.; Gomes, L.C. Effects of water quantity on connectivity: The case of the upper Paraná River floodplain. Ecohydrol. Hydrobiol. 2009, 9, 99–113. [Google Scholar] [CrossRef]
- Bonecker, C.C.; Simões, N.; Minte-Vera, C.V.; Lansac-Tôha, F.A.; Velho, L.F.M.; Agostinho, A. Temporal changes in zooplankton species diversity in response to environmental changes in an alluvial valley. Limnologica 2013, 43, 114–121. [Google Scholar] [CrossRef]
- Winemiller, K.O.; McIntyre, P.B.; Castello, L.; Fluet-Chouinard, E.; Giarrizzo, T.; Nam, S.; Baird, I.G.; Darwall, W.; Lujan, N.K.; Harrison, I.; et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 2016, 351, 128–129. [Google Scholar] [CrossRef] [Green Version]
- Oberdorff, T.; Pont, D.; Hugueny, B.; Porcher, J.-P. Development and validation of a fish-based index for the assessment of ‘river health’ in France. Freshw. Biol. 2002, 47, 1720–1734. [Google Scholar] [CrossRef]
- Arthington, A.H.; Dulvy, N.K.; Gladstone, W.; Winfield, I.J. Fish conservation in freshwater and marine realms: Status, threats and management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 838–857. [Google Scholar] [CrossRef] [Green Version]
- Olden, J.D. Challenges and opportunities for fish conservation in dam-impacted waters. In Conservation of Freshwater Fishes; Closs, G.P., Krkosek, M., Olden, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 107–148. [Google Scholar]
- Pereira, H.R.; Gomes, L.F.; Barbosa, H.D.O.; Pelicice, F.M.; Nabout, J.C.; Teresa, F.B.; Vieira, L.C.G. Research on dams and fishes: Determinants, directions, and gaps in the world scientific production. Hydrobiologia 2020, 847, 579–592. [Google Scholar] [CrossRef]
- Díaz, G.; Górski, K.; Heino, J.; Arriagada, P.; Link, O.; Habit, E. The longest fragment drives fish beta diversity in fragmented river networks: Implications for river management and conservation. Sci. Total Environ. 2021, 766, 144323. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.; Olden, J.D.; Zanden, M.J.V. Dam invaders: Impoundments facilitate biological invasions into freshwaters. Front. Ecol. Environ. 2008, 6, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, K.; Turpin, C.; Gregory-Eaves, I. Dams have varying impacts on fish communities across latitudes: A quantitative synthesis. Ecol. Lett. 2019, 22, 1501–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Qin, J.; Xu, Y.; Ouyang, S.; Wu, X. Biodiversity decline of fish assemblages after the impoundment of the Three Gorges Dam in the Yangtze River Basin, China. Rev. Fish Biol. Fish. 2019, 29, 177–195. [Google Scholar] [CrossRef]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proc. Natl. Acad. Sci. USA 2020, 117, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.W.; Olden, J.D. Response diversity, nonnative species, and disassembly rules buffer freshwater ecosystem processes from anthropogenic change. Glob. Chang. Biol. 2017, 23, 1871–1880. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Past progress and future directions. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Heino, J.; Tolonen, K.T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol. Oceanogr. 2017, 62, 2431–2444. [Google Scholar] [CrossRef]
- Heino, J.; Alahuhta, J.; Fattorini, S.; Schmera, D. Predicting beta diversity of terrestrial and aquatic beetles using ecogeographical variables: Insights from the replacement and richness difference components. J. Biogeogr. 2018, 46, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Soininen, J.; Heino, J.; Wang, J. A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosystems. Glob. Ecol. Biogeogr. 2018, 27, 96–109. [Google Scholar] [CrossRef] [Green Version]
- López-Delgado, E.O.; Winemiller, K.O.; Villa-Navarro, F.A. Local environmental factors influence beta-diversity patterns of tropical fish assemblages more than spatial factors. Ecology 2020, 101, e02940. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, J.D.; Altermatt, F.; Finn, D.S.; Heino, J.; Olden, J.D.; Pauls, S.U.; Lytle, D.A. The role of dispersal in river network metacommunities: Patterns, processes, and pathways. Freshw. Biol. 2018, 63, 141–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Comte, L.; Xian, W.; Chen, Y.; Olden, J.D. Current and projected future risks of freshwater fish invasions in China. Ecography 2019, 42, 2074–2083. [Google Scholar] [CrossRef]
- Rogosch, J.S.; Olden, J.D. Dynamic contributions of intermittent and perennial streams to fish beta diversity in dryland rivers. J. Biogeogr. 2019, 46, 2311–2322. [Google Scholar] [CrossRef]
- Leprieur, F.; Olden, J.D.; Lek, S.; Brosse, S. Contrasting patterns and mechanisms of spatial turnover for native and exotic freshwater fish in Europe. J. Biogeogr. 2009, 36, 1899–1912. [Google Scholar] [CrossRef]
- Liu, X.; Hu, X.; Ao, X.; Wu, X.; Ouyang, S. Community characteristics of aquatic organisms and management implications after construction of Shihutang Dam in the Gangjiang River, China. Lake Reserv. Manag. 2017, 34, 1–16. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, X.; Ao, X.; Qin, J.; Wu, X.; Ouyang, S. Fish diversity in the middle and lower reaches of the Ganjiang River of China: Threats and conservation. PLoS ONE 2018, 13, e205116. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.L.; Guo, C.B.; Wang, R.; Zeng, Z.G.; Zhang, E.; Chen, Y.S. Research Status of Water Environment and Fish Resources Under Multiple Human Disturbances in the Gan-Jiang River Basin: A Review. Resour. Environ. Yangtze Basin 2019, 28, 2879–2892. [Google Scholar] [CrossRef]
- Tian, J.L. Fish resources in Ganjiang River before the construction of Wan’an Dam and its fishery utilization. Freshw. Fish. 1989, 19, 33–39. [Google Scholar]
- Guo, Z.Z.; Liu, R.L. The fishes of Jiangxi Province. J. Nanchang Univ. 1995, 19, 222–232. [Google Scholar]
- Zou, S.Z. Effects of Large Hydraulic Project on Fish and Ecological Environment in the Middle Reach of Gan River. PhD Thesis, Nanchang University, Nanchang, China, 2011. [Google Scholar]
- Su, N.; Li, L.; Xu, Z.Q.; Xiao, D. Status of fish resources in Xiajiang to Nanchang section of Ganjiang River. J. Huazhong Agric. Univ. 2012, 31, 756–764. [Google Scholar] [CrossRef]
- Wang, C.Y.; Huang, D.C.; Hu, M.L.; Liu, J.; Miao, C.; Wang, T.; Jian, S.Q. Comparative analysis of fish community diversity in two primary tributaries of the lower Ganjiang River. J. Hydroecol. 2019, 40, 99–106. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Wu, H.; Yang, J.; Hu, M. Taxonomic diversity pattern and composition of fish species in the upper reaches of Ganjiang River, Jiangxi, China. PLoS ONE 2020, 15, e0241762. [Google Scholar] [CrossRef] [PubMed]
- Dornelas, M.; Gotelli, N.J.; McGill, B.; Shimadzu, H.; Moyes, F.; Sievers, C.; Magurran, A.E. Assemblage Time Series Reveal Biodiversity Change but Not Systematic Loss. Science 2014, 344, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; He, D.; Chen, Y.; Olden, J.D. Species invasions threaten the antiquity of China’s freshwater fish fauna. Divers. Distrib. 2017, 23, 556–566. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Watson, M. Guidance on Conducting a Systematic Literature Review. J. Plan. Educ. Res. 2019, 39, 93–112. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: http://www.fishbase.org (accessed on 7 September 2021).
- Liu, X.; Wu, R.; Chen, X.; Zhou, Y.; Yang, L.; Ouyang, S.; Wu, X. Effects of dams and their environmental impacts on the genetic diversity and connectivity of freshwater mussel populations in Poyang Lake Basin, China. Freshw. Biol. 2019, 65, 264–277. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.; Pusey, B.J. Species invasions and the changing biogeography of Australian freshwater fishes. Glob. Ecol. Biogeogr. 2008, 17, 25–37. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. 2014. Available online: https://Rproject.org (accessed on 23 December 2016).
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-3. 2018. Available online: https://CRAN.R-project.org/package¼vegan (accessed on 5 March 2018).
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Lep, S.J.; Smilauer, P. Ultivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, L. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Hu, M.; Hua, Q.; Zhou, H.; Wu, Z.; Wu, X. The effect of dams on the larval abundance and composition of four carp species in key river systems in China. Environ. Bio. Fish 2015, 98, 1201–1205. [Google Scholar] [CrossRef] [Green Version]
- Vitule, J.R.S.; Skóra, F.; Abilhoa, V. Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers. Distrib. 2012, 18, 111–120. [Google Scholar] [CrossRef]
- Xie, P. Biodiversity crisis in the Yangtze River: The culprit was dams, followed by overfishing. J. Lake Sci. 2017, 29, 1279–1299. [Google Scholar] [CrossRef]
- Xiong, F.; Olden, J.D.; Lu, Y.; Liu, H.; Qu, X.; Xia, W.; Guo, C.; Wu, X.; Infante, D.M.; Wang, L.; et al. Riparian land use and in-channel stressors drive fish community structure in the Yangtze River. Landsc. Ecol. 2021, 36, 3079–3095. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Tang, W.; Heino, J.; Jiang, X. Effects of dam construction and fish invasion on the species, functional and phylogenetic diversity of fish assemblages in the Yellow River Basin. J. Environ. Manag. 2021, 29, 112863. [Google Scholar] [CrossRef]
- Kang, B.; He, D.; Perrett, L.; Wang, H.; Hu, W.; Deng, W.; Wu, Y. Fish and fisheries in the Upper Mekong: Current assessment of the fish community, threats and conservation. Rev. Fish Biol. Fish. 2009, 19, 465–480. [Google Scholar] [CrossRef]
- Sá-Oliveira, J.C.; Hawes, J.E.; Isaac-Nahum, V.J.; Peres, C. Upstream and downstream responses of fish assemblages to an eastern Amazonian hydroelectric dam. Freshw. Biol. 2015, 60, 2037–2050. [Google Scholar] [CrossRef]
- Harper, M.; Rytwinski, T.; Taylor, J.J.; Bennett, J.R.; Smokorowski, K.E.; Olden, J.D.; Clarke, K.D.; Pratt, T.; Fisher, N.; Leake, A.; et al. How do changes in flow magnitude due to hydropower operations affect fish abundance and biomass in temperate regions? A systematic review. Environ. Évid. 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Belmar, O.; Maire, A.; Morel, A.; Dumont, B.; Datry, T. Structural and functional responses of invertebrate communities to climate change and flow regulation in alpine catchments. Glob. Chang. Biol. 2019, 25, 1612–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattorini, S. The influence of geographical and ecological factors on island beta diversity patterns. J. Biogeogr. 2010, 37, 1061–1070. [Google Scholar] [CrossRef]
- Hirao, T.; Kubota, Y.; Murakami, M. Geographical patterns of butterfly species diversity in the subtropical Ryukyu Islands: The importance of a unidirectional filter between two source islands. J. Biogeogr. 2015, 42, 1418–1430. [Google Scholar] [CrossRef]
- Fu, C.; Wu, J.; Chen, J.; Wu, Q.; Lei, G. Freshwater fish biodiversity in the Yangtze River basin of China: Patterns, threats and conservation. Biodivers. Conserv. 2003, 12, 1649–1685. [Google Scholar] [CrossRef]
- Huang, L.L.; Wu, Z.Q.; Li, J.H. Fish fauna, biogeography and conservation of freshwater fish in Poyang Lake Basin, China. Environ. Bio. Fish 2011, 96, 1229–1243. [Google Scholar] [CrossRef]
- Wang, K.; Shi, X.Z.; Yu, D.S.; Shi, D.M.; Chen, J.M.; Xu, B.B.; Liang, Y.; Li, D.C. Environmental factors affecting temporal and spatial dynamics of soil erosion in Xingguo County, South China. Pedosphere 2005, 15, 620–627. [Google Scholar]
- Zhang, Z.Y.; Liu, M.; Peng, A.C.; Hu, L.; Wang, C.Y.; Chen, W.X. Main problems and restoration measures of fish habitat in Poyang Lake. J. Hydroecol. 2011, 32, 134–136. [Google Scholar] [CrossRef]
- Craig, L.S.; Olden, J.D.; Arthington, A.H.; Entrekin, S.; Hawkins, C.P.; Kelly, J.J.; Kennedy, T.A.; Maitland, B.M.; Rosi, E.J.; Roy, A.H.; et al. Meeting the challenge of interacting threats in freshwater ecosystems: A call to scientists and managers. Elementa Sci. Anthr. 2017, 5, 72. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, C.; Fan, E.; Zhao, Y. Freshwater fishes of China: Species richness, endemism, threatened species and conservation. Divers. Distrib. 2015, 22, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wei, H.; Vilizzi, L.; Zhan, A.; Olden, J.D.; Preston, D.L.; Clarke, S.A.; Cudmore, B.; Davies, G.D.; Wang, X.; et al. The Future of Legislation, Policy, Risk Analysis, and Management of Non-Native Freshwater Fishes in China. Rev. Fish. Sci. Aquac. 2020, 29, 149–166. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River | Code | Total Species | Native Species | Alien Species |
|---|---|---|---|---|
| Xiang River | XR | 47 | 46 | 1 |
| Lian River | LR | 31 | 31 | 0 |
| Ping River | PR | 62 | 62 | 0 |
| Mei River | MR | 87 | 86 | 1 |
| Tao River | TR | 91 | 91 | 0 |
| Zhang River | ZR | 73 | 73 | 0 |
| Shangyou River | SR | 73 | 73 | 0 |
| Yuan River | YR | 107 | 104 | 3 |
| Jin River | JR | 93 | 92 | 1 |
| Suichuan River | SCR | 107 | 107 | 0 |
| Shushui River | SSR | 42 | 42 | 0 |
| Lushui River | LSR | 66 | 65 | 1 |
| Heshui River | HR | 67 | 66 | 1 |
| Mainstem of middle reach of the Gan River | TSM | 117 | 117 | 0 |
| Mainstem of upper of the Gan River | TSU | 88 | 86 | 2 |
| Mainstem of lower reach of the Gan River | TSL | 111 | 111 | 0 |
| Tributary of upper reach of the Gan River | TU | 120 | 119 | 1 |
| Tributary of middle reach of the Gan River | TM | 126 | 125 | 1 |
| Tributary of lower reach of the Gan River | TL | 120 | 116 | 4 |
| Upper reach of the Gan River | BU | 124 | 121 | 3 |
| Middle reach of the Gan River | BM | 147 | 146 | 1 |
| Lower reach of the Gan River | BL | 143 | 139 | 4 |
| All tributaries | T | 158 | 153 | 5 |
| All mainstems | TS | 146 | 144 | 2 |
| α Diversity | β Diversity | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| All Species | Native Species | All Species | Native Species | ||||||
| βsor | βsim | βsne | βsor | βsim | βsne | ||||
| Geographical factors | DL | 0.729 ** | 0.725 ** | 0.562 ** | −0.306 ** | 0.713 ** | 0.563 ** | −0.286 ** | 0.708 ** |
| DA | 0.737 ** | 0.736 ** | 0.625 ** | −0.205 | 0.698 ** | 0.626 ** | −0.190 * | 0.696 ** | |
| AP | 0.033 | 0.030 | −0.060 | −0.152 | 0.055 | −0.061 | −0.151 * | 0.053 | |
| AAT | 0.170 * | 0.167 * | 0.327 ** | 0.224 ** | 0.130 * | 0.327 * | 0.230 * | 0.128 | |
| AA | 0.018 | 0.021 | 0.090 | 0.111 | 0.001 | 0.095 | 0.111 | 0.006 | |
| Environmental factors | WT | 0.260 * | 0.259 * | 0.456 * | 0.286 * | 0.200 * | 0.461 * | 0.301 * | 0.197 |
| pH | −0.218 * | −0.220 * | −0.142 | 0.136 | −0.222 * | −0.147 | 0.128 | −0.223 * | |
| DO | −0.025 | −0.023 | −0.031 | 0.014 | −0.037 | −0.025 | 0.018 | −0.035 | |
| TURB | −0.106 | −0.113 | −0.001 | 0.166 | −0.118 | 0.008 | 0.192 | −0.129 | |
| Chl-a | −0.003 | 0.001 | −0.071 | −0.104 | 0.011 | −0.069 | −0.105 | 0.013 | |
| NH4+ | 0.053 | 0.053 | 0.150 | 0.151 | 0.026 | 0.146 | 0.146 | 0.027 | |
| NO3- | 0.176 | 0.180 | 0.019 | −0.267 ** | 0.206 | 0.024 | −0.262 ** | 0.209 | |
| TP | −0.018 | −0.021 | 0.040 | 0.075 | -0.018 | 0.046 | 0.091 | −0.024 | |
| TN | 0.249 * | 0.250 * | 0.152 | −0.192 | 0.271 ** | 0.152 | −0.191 | 0.272 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Olden, J.D.; Wu, R.; Ouyang, S.; Wu, X. Dam Construction Impacts Fish Biodiversity in a Subtropical River Network, China. Diversity 2022, 14, 476. https://doi.org/10.3390/d14060476
Liu X, Olden JD, Wu R, Ouyang S, Wu X. Dam Construction Impacts Fish Biodiversity in a Subtropical River Network, China. Diversity. 2022; 14(6):476. https://doi.org/10.3390/d14060476
Chicago/Turabian StyleLiu, Xiongjun, Julian D. Olden, Ruiwen Wu, Shan Ouyang, and Xiaoping Wu. 2022. "Dam Construction Impacts Fish Biodiversity in a Subtropical River Network, China" Diversity 14, no. 6: 476. https://doi.org/10.3390/d14060476