
Nearctic Species in the Palearctic: Trans-Beringian Range, Phylogeny and Phylogeography of Pterostichus (Cryobius) mandibularoides (Coleoptera, Carabidae)

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic and Phylogeographic Analyses
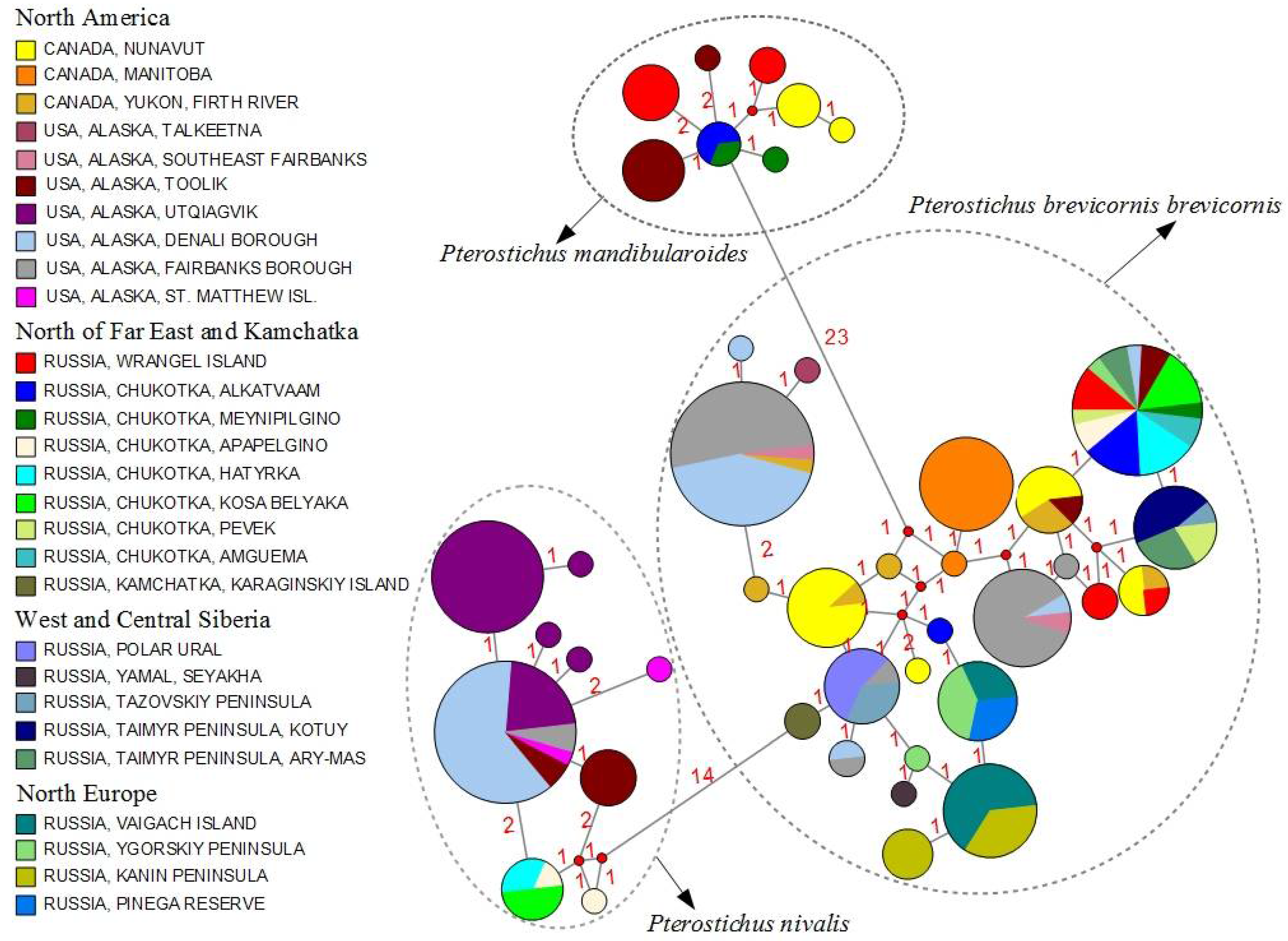
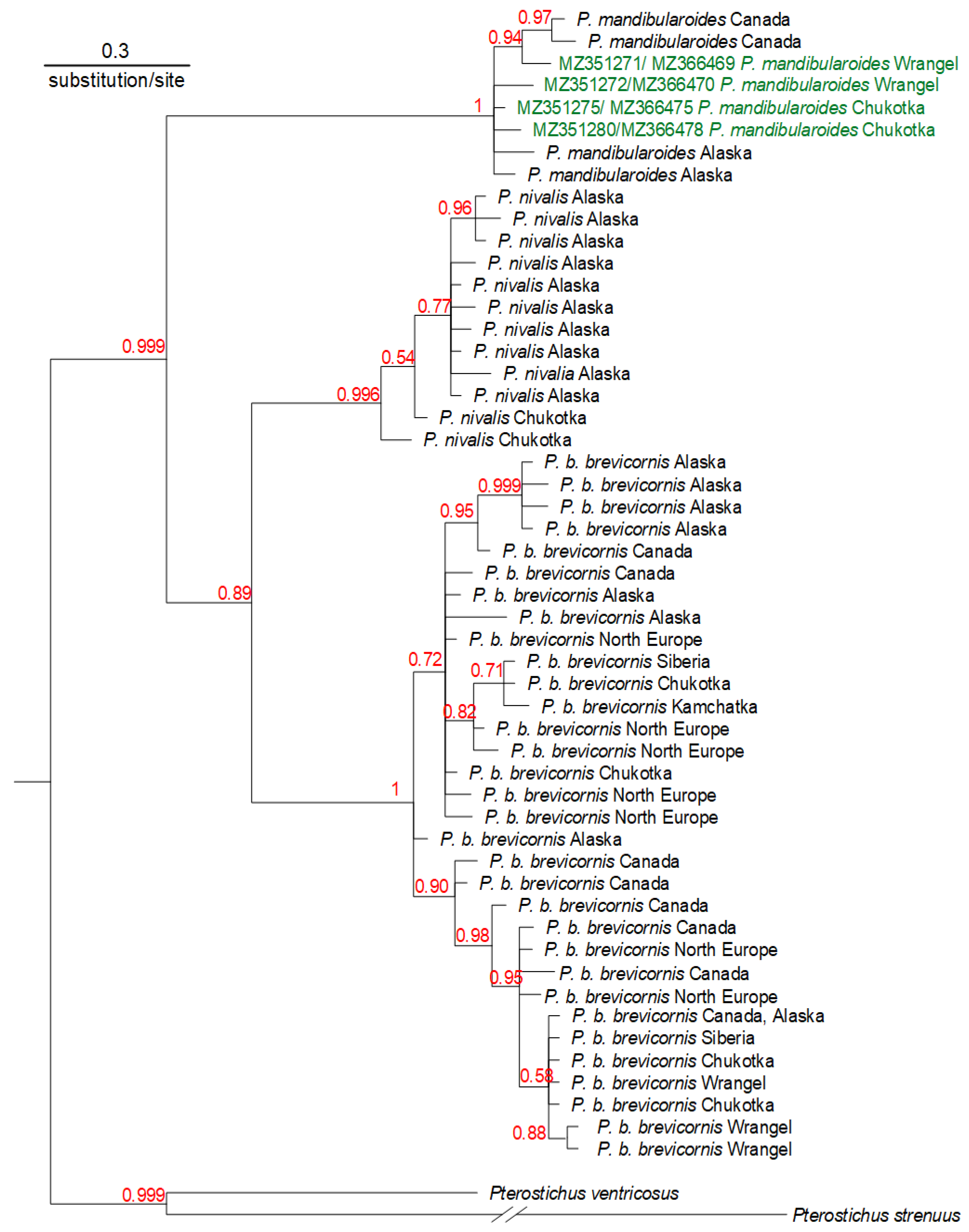
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeChaine, E.G. A bridge or a barrier? Beringia’s influence on the distribution and diversity of tundra plants. Plant Ecol. Divers. 2008, 1, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Elias, S.A.; Crocker, B. The Bering Land Bridge: A moisture barrier to the dispersal of steppe–tundra biota? Quat. Sci. Rev. 2008, 27, 2473–2483. [Google Scholar] [CrossRef]
- Elias, S.A. Advances in Quaternary Entomology. In Developments in Quaternary Sciences; Jaap, J.M., van der Meer, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 12, pp. 1–288. [Google Scholar]
- Jiang, D.; Li, J.-T.; Klaus, S.; Zhang, Y.-P.; Hillis, D.M. Asymmetric biotic interchange across the Bering land bridge between Eurasia and North America. Natl. Sci. Rev. 2019, 6, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Potapov, G.S.; Berezin, M.V.; Kolosova, Y.S.; Kondakov, A.V.; Tomilova, A.A.; Spitsyn, V.M.; Zheludkova, A.A.; Zubrii, N.A.; Filippov, B.Y.; Bolotov, I.N. The last refugia for a polar relict pollinator: Isolates of Bombus glacialis on Novaya Zemlya and Wrangel Island indicate its broader former range in the Pleistocene. Polar Biol. 2021, 44, 1691–1709. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Tatarinov, A.G.; Filippov, B.Y.; Gofarov, M.Y.; Kondakov, A.V.; Kulakova, O.I.; Potapov, G.S.; Zubryi, N.A.; Spitsyn, V.M. The distribution and biology of Pararctia subnebulosa (Dyar, 1899) (Lepidoptera: Erebidae: Arctiinae), the largest tiger moth species in the High Arctic. Polar Biol. 2015, 38, 905–911. [Google Scholar] [CrossRef]
- Kullberg, J.; Filippov, B.Y.; Spitsyn, V.M.; Zubrij, N.A.; Kozlov, M.V. Moths and butterflies (Insecta: Lepidoptera) of the Russian Arctic islands in the Barents Sea. Polar Biol. 2019, 42, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Kullberg, J.; Filippov, B.Y.; Zubrij, N.A.; Kozlov, M. Faunistic notes on Lepidoptera collected from arctic tundrain European Russia. Nota Lepidopterol. 2013, 36, 127–136. [Google Scholar]
- Khruleva, O.A.; Stekolshchikov, A.V. Additions to the aphid fauna of Wrangel Island due to climate change with redescription of the oviparous female of Pterocomma groenlandicum Hille Ris Lambers, 1952 (Hemiptera, Aphidoidea). Zootaxa 2019, 4615, 511–528. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Khruleva, O.A. Taxonomic Position and Status of Arctic Gynaephora and Dicallomera Moths (Lepidoptera, Erebidae, Lymantriinae). Folia Biol. 2015, 63, 257–261. [Google Scholar] [CrossRef]
- Shamshev, I.V.; Sinclair, B.J.; Khruleva, O.A. The empidoid flies (Diptera: Empidoidea, exclusive of Dolichopodidae) of the Russian Arctic islands and Svalbard Archipelago. Zootaxa 2020, 4848, 001–075. [Google Scholar] [CrossRef]
- Barkalov, A.V.; Khruleva, O.A. Hoverflies (Diptera, Syrphidae) of Wrangel Island (Chukotka Autonomous Okrug, Russia). Nat. Conserv. Res. 2021, 6, 78–87. [Google Scholar] [CrossRef]
- Nartshuk, E.P.; Khruleva, O.A. Plant-feeding Dipterans (Diptera, Chloropidae, Agromyzidae) from Wrangel Island (the Chukotsk Sea). Entomol. Rev. 2011, 91, 849–854. [Google Scholar] [CrossRef]
- Sorokina, V.S.; Khruleva, O.A. Details of species composition and distribution of house-flies (Diptera, Muscidae) of the Wrangel Island, Russia. Euroasian Entomol. J. 2012, 11, 553–564. (In Russian) [Google Scholar]
- Sorokina, V.S.; Tridrikh, N.N. An annotated checklist of the Muscidae (Diptera) of Chukotka (Russia), with new records. Ann. de la Société Entomol. de Fr. (N.S.) 2021, 57, 205–234. [Google Scholar] [CrossRef]
- Chernov, Y.I.; Makarova, O.L.; Penev, L.D.; Khruleva, O.A. Beetles (Insecta, Coleoptera) in the Arctic Fauna: Communication 1. Faunal Composition. Entomol. Rev. 2014, 94, 438–478. [Google Scholar] [CrossRef]
- Ball, G.E. Descriptions of eleven new species of the beetle subgenus Cryobius (Pterostichus, Coleoptera, Carabidae) from Alaska and northwestern Canada. Opusc. Entomol. 1962, 28, 1–26. [Google Scholar]
- Ball, G.E. The distribution of the species of the subgenus Cryobius (Pterostichus, Coleoptera, Carabidae) with special reference to the Bering land bridge and Pleistocene refugia. In Pacific Basin Biogeography–A Symposium; Bishop Museum Press: Honolulu, HI, USA, 1963; pp. 133–149. [Google Scholar]
- Ball, G.E. A Revision of the North American Species of the subgenus Cryobius Chaudoir (Pterostichus, Carabidae, Coleoptera). Opusc. Entomol. 1966, 28, 1–165. [Google Scholar]
- Lindroth, C.H. The ground-beetles (Carabidae, excl. Cicindelinae) of Canada and Alaska. Opusc. Entomol. Suppl. 1966, 29, 409–648. [Google Scholar]
- Lindroth, C.H. Ground beetles (Carabidae) of Fennoscandia. A zoogeographic Study. In Part 3: General Analysis with a Discussion on Biogeographic Principles; Adis, J., Erwin, T.L., Eds.; Amerind Publishing Co. Pvt. Ltd.: New Delhi, India, 1992; p. 814. [Google Scholar]
- Erjiomin, P.K. Palaearctic Species of the Pterostichus (Cryobius) brevicornis Group (Coleoptera, Carabidae). Zool. Zh. 1998, 77, 295–302. (In Russian) [Google Scholar]
- Kryzhanovskij, O.L.; Belousov, I.A.; Kabak, I.I.; Kataev, B.M.; Makarov, K.V.; Shilinkov, V.G. A Check-List of the Ground-Beetles of Russia and Adjacent Lands (Insecta, Coleoptera, Carabidae); Pensoft Publishers: Sofia, Bulgaria, 1995; p. 271. [Google Scholar]
- Morita, S. Pterostichine carabid beetles of the subgenus Cryobius (Coleoptera, Carabidae) from north Japan. Elytra 2002, 30, 73–89. [Google Scholar]
- Löbl, I.; Löbl, D. Catalogue of Palaearctic Coleoptera. Volume 1. Revised and Updated Edition. In Archostemata—Myxophaga—Adephaga; Brill: Leiden The Netherlands; Boston, MA, USA, 2017; Volume XXXIV, p. 1443. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; p. 1659. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Foighil, D.O. Sphaeriid and corbiculid clams represent separate heterodont bivalve radiations into freshwater environments. Mol. Phylogenetics Evol. 2000, 14, 75–88. [Google Scholar] [CrossRef]
- Jovelin, R.; Justine, J.L. Phylogenetic relationships within the polyopisthocotylean monogeneans (Platyhelminthes) inferred from partial 28S rDNA sequences. Int. J. Parasitol. 2001, 31, 393–401. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT/T. A. Hall. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Villesen, P. FaBox: An online toolbox for FASTA sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigThree v1.4.2. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 25 July 2021).
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Khobrakova, L.T.; Shilenkov, V.G.; Dudko, R.Y. The Ground beetles (Coleoptera, Carabidae) of Buryatia; Buryat Scientific Center SB RAS Press: Ulan-Ude, Russia, 2014; p. 380. (In Russian) [Google Scholar]
- Elias, S.A.; Matthews, J.V. Arctic North American seasonal temperatures from the latest Miocene to the Early Pleistocene, based on mutual climatic range analysis of fossil beetle assemblages. Can. J. Earth Sci. 2002, 39, 911–920. [Google Scholar] [CrossRef]
- Kienast, F.; Wetterich, S.; Kuzmina, S.; Schirrmeister, L.; Andreev, A.; Tarasov, P.; Nazarova, L.; Kossler, A.; Frolova, L.; Kunitsky, V. Paleontological records indicate the occurrence of open woodlands in a dry inland climate at the present-day Arctic coast in western Beringia during the Last Interglacial Quaternary. Sci. Rev. 2011, 30, 2134–2159. [Google Scholar] [CrossRef] [Green Version]
- Kuzmina, S.; Froese, D.G.; Jensen, B.J.L.; Hall, E.; Zazula, G.D. Middle Pleistocene (MIS 7) to Holocene fossil insect assemblages from the Old Crow basin, northern Yukon, Canada. Quat. Int. 2014, 341, 216–242. [Google Scholar] [CrossRef]
- Kuzmina, S.A.; Elias, S.A.; Kotov, A.A. Late Quaternary insects and freshwater invertebrates of the Alaskan North Slope and paleoenvironmental reconstructions in Arctic Alaska. Invert. Zool. 2019, 16, 89–125. [Google Scholar] [CrossRef]
- Kuzmina, S.A. Macroentomology analysis: Methods, opportunities, and examples of reconstructions of paleoclimatic and paleoenvironmental conditions in the Quaternary of the northeastern. Sib. Contemp. Probl. Ecol. 2017, 10, 336–349. [Google Scholar] [CrossRef]
- Filippov, B.; Shuvalov, E. Ground beetles of the Kanin Peninsula southern tundras. Arct. Environ. Res. 2006, 1, 99–109. (In Russian) [Google Scholar]
- Filippov, B. Fauna and ground beetles composition (Coleoptera, Carabidae) of bog habitats in Kanin Peninsula. Arct. Environ. Res. 2008, 1, 45–53. (In Russian) [Google Scholar]
- Mokhnatkin, A.; Zezin, I.; Filippov, B. Ground beetles (Coleoptera, Carabidae) of different karst landscape biocenoses of the south-eastern part of Belomor-Kuloi Plateau. Arct. Environ. Res. 2010, 4, 59–64. (In Russian) [Google Scholar]
- Zenkova, I.; Pozharskaya, V.; Filippov, B.; Kolesnikova, A.; Seredyuk, S. AltitudinaL zonality and beetles in soils of the Khibiny mountains. Tr. Karelian Res. Cent. Russ. Acad. Sci. 2011, 2, 107–118. (In Russian) [Google Scholar]
- Zubriy, N.A.; Filippov, B.Y. Carabid beetles local fauna (Coleoptera, Carabidae) of the typical tundra of the Yugor Peninsula. Arct. Environ. Res. 2015, 2, 46–55. (In Russian) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | GPS Code | Number of Specimens | Site |
|---|---|---|---|
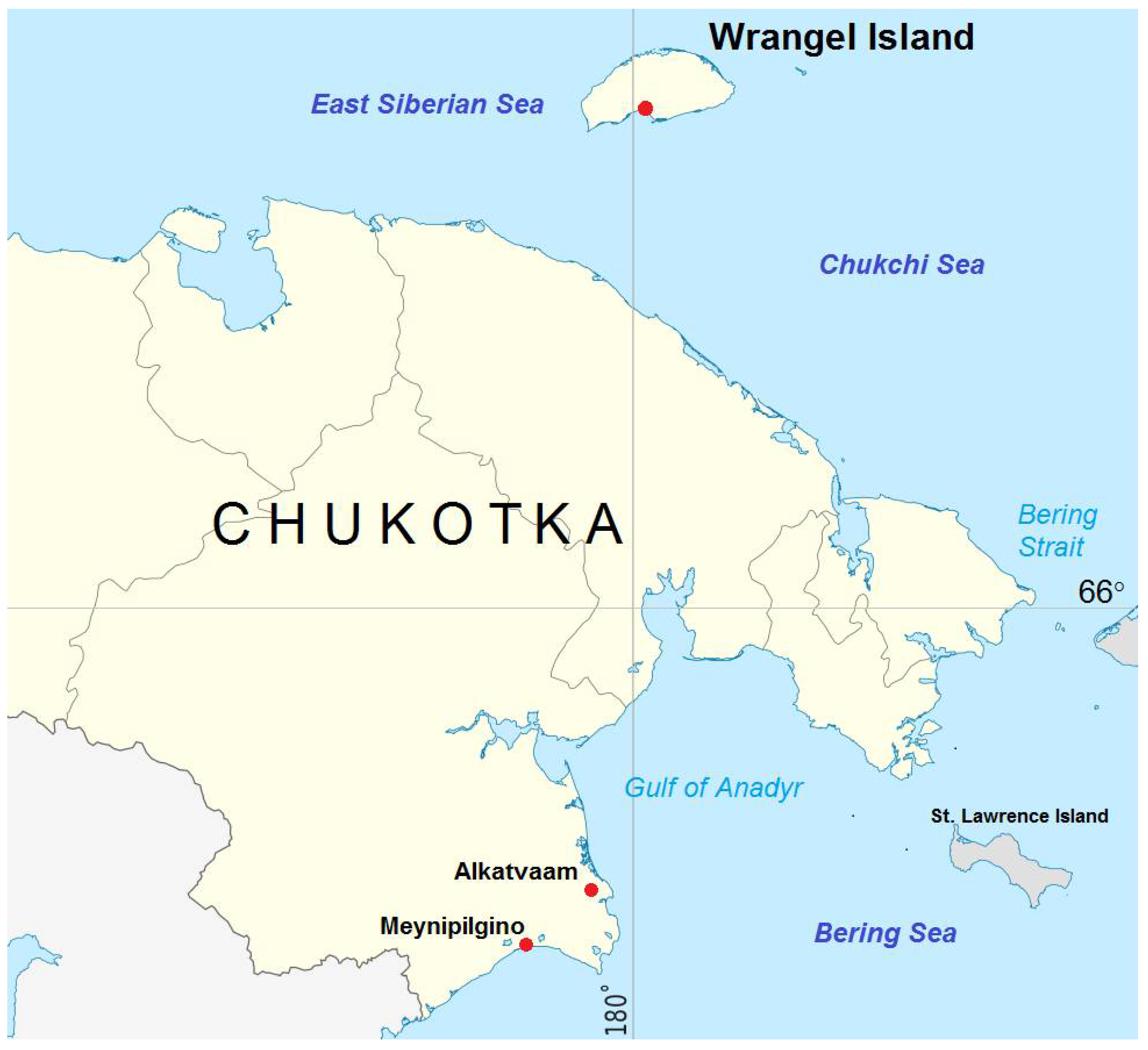
| Chukotka, suburbs of Alkatvaam settlement, 2019 | 63°8′9.6″ N, 179°2′9.6″ E | 49 | Well-drained slope with mossy–shrub–rhododendron tundra |
| Chukotka, suburbs of Meynipilgino settlement, 2020 | 62°33′3.6″ N, 177°4′4.2″ E | 9 | Damp depression with mossy–Salix vegetation |
| Wrangel Island, Somnitelnye Mountain, 2015 | 70°58′1.2″ N, 179°34′58.8″ E | 2 | Wet loam soil at the foot of the hill with Salix–dryad–sedge and mossy spotted vegetation |
| 70°59′11.4″ N, 179°35′14.2″ E | 2 | Dry loam–rubble soil on a raised terrace of the southern expose with Salix and grass–mossy vegetation | |
| Wrangel Island, Somnitelnaya Bay, 2015 | 70°57′7.6″ N, 179°32′26.5″ E | 3 | Dry loam–rubble soil on plain with grassy–dryad–sedge spotted vegetation |
| Species | P. mandibularoides | P. b. brevicornis | P. nivalis |
|---|---|---|---|
| Pterostichus mandibularoides | 5.45 ± 1.11 | 5.42 ± 1.12 | |
| Pterostichus b. brevicornis | 5.25 ± 0.87 | 3.98 ± 0.88 | |
| Pterostichus nivalis | 5.60 ± 0.98 | 3.37 ± 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubrii, N.A.; Filippov, B.Y.; Khruleva, O.A.; Kondakov, A.V.; Rybalov, L.B. Nearctic Species in the Palearctic: Trans-Beringian Range, Phylogeny and Phylogeography of Pterostichus (Cryobius) mandibularoides (Coleoptera, Carabidae). Diversity 2022, 14, 415. https://doi.org/10.3390/d14060415
Zubrii NA, Filippov BY, Khruleva OA, Kondakov AV, Rybalov LB. Nearctic Species in the Palearctic: Trans-Beringian Range, Phylogeny and Phylogeography of Pterostichus (Cryobius) mandibularoides (Coleoptera, Carabidae). Diversity. 2022; 14(6):415. https://doi.org/10.3390/d14060415
Chicago/Turabian StyleZubrii, Natalia Andreevna, Boris Yurevich Filippov, Olga Arturovna Khruleva, Alexander Vasilevich Kondakov, and Leonid Borisovich Rybalov. 2022. "Nearctic Species in the Palearctic: Trans-Beringian Range, Phylogeny and Phylogeography of Pterostichus (Cryobius) mandibularoides (Coleoptera, Carabidae)" Diversity 14, no. 6: 415. https://doi.org/10.3390/d14060415