
Three-Dimensional Quantification of Copepods Predictive Distributions in the Ross Sea: First Data Based on a Machine Learning Model Approach and Open Access (FAIR) Data


Abstract
:1. Introduction
2. Materials and Methods
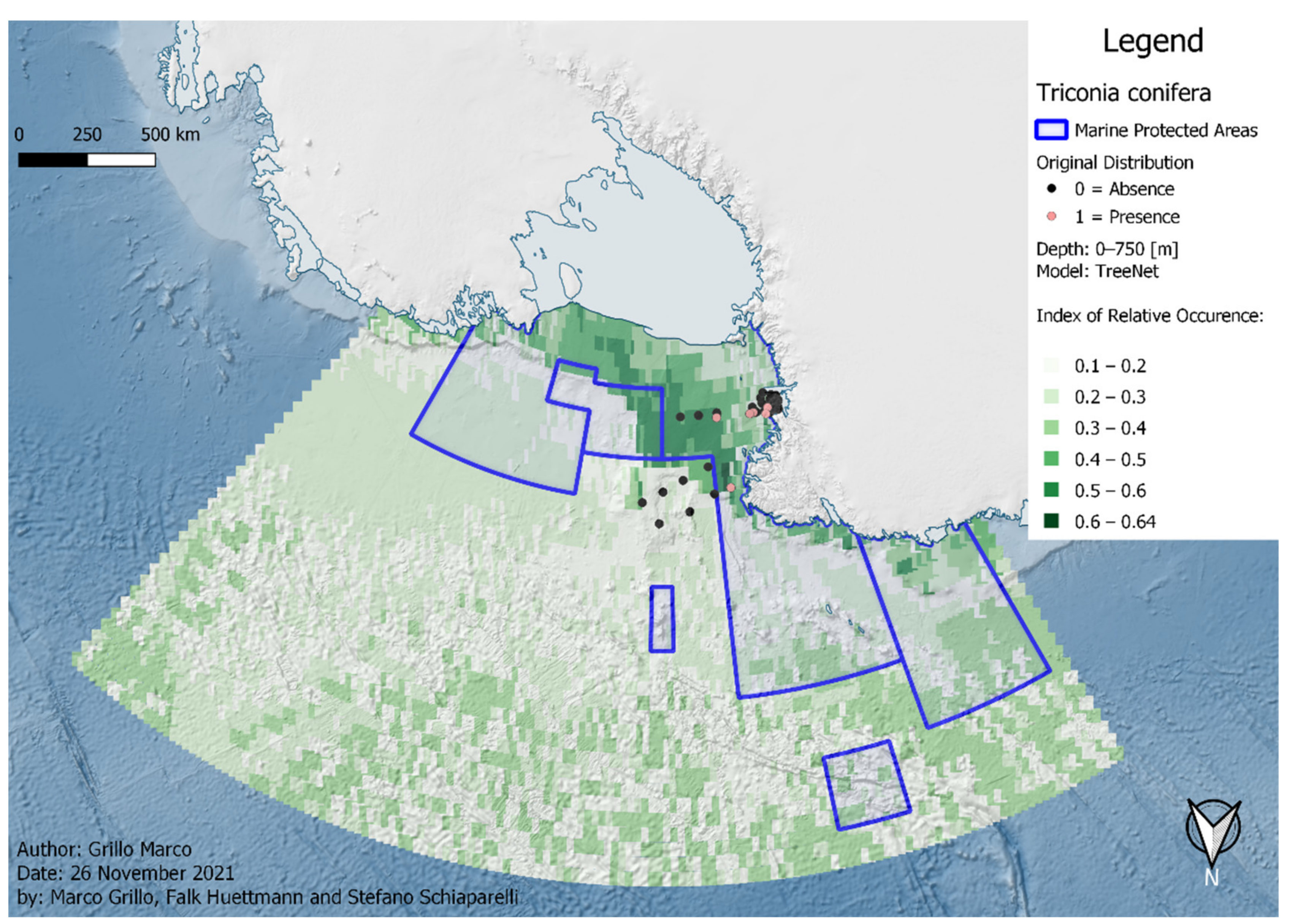
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ainley, D.G.; Brooks, C.M.; Eastman, J.T.; Massaro, M. Unnatural Selection of Antarctic Toothfish in the Ross Sea, Antarctica. In Protection of the Three Poles; Springer: Berlin/Heidelberg, Germany, 2012; pp. 53–75. [Google Scholar]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2016. In Contributing to Food Security and Nutrition for All; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, C.M.; Crowder, L.B.; Österblom, H.; Strong, A.L. Reaching Consensus for Conserving the Global Commons: The Case of the Ross Sea, Antarctica. Conserv. Lett. 2020, 13, e12676. [Google Scholar] [CrossRef]
- Ainley, D.G.; Ballard, G.; Jones, R.M.; Jongsomjit, D.; Pierce, S.D.; Smith, W.O.J.; Veloz, S. Trophic Cascades in the Western Ross Sea, Antarctica: Revisited. Mar. Ecol. Prog. Ser. 2015, 534, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Huettmann, F.; Schmid, M.S.; Humphries, G.R.W. A First Overview of Open Access Digital Data for the Ross Sea: Complexities, Ethics, and Management Opportunities. Hydrobiologia 2015, 761, 97–119. [Google Scholar] [CrossRef]
- Mormede, S.; Dunn, A.; Parker, S.; Hanchet, S. Using Spatial Population Models to Investigate the Potential Effects of the Ross Sea Region Marine Protected Area on the Antarctic Toothfish Population. Fish. Res. 2017, 190, 164–174. [Google Scholar] [CrossRef]
- Behrens, E.; Pinkerton, M.; Parker, S.; Rickard, G.; Collins, C. The Impact of Sea-Ice Drift and Ocean Circulation on Dispersal of Toothfish Eggs and Juveniles in the Ross Gyre and Amundsen Sea. J. Geophys. Res. Ocean. 2021, 126, e2021JC017329. [Google Scholar] [CrossRef]
- Parker, S.J.; Sundby, S.; Stevens, D.; Di Blasi, D.; Schiaparelli, S.; Ghigliotti, L. Buoyancy of Post-Fertilised Dissostichus Mawsoni Eggs and Implications for Early Life History. Fish. Oceanogr. 2021, 30, 697–706. [Google Scholar] [CrossRef]
- Atkinson, A. Life Cycle Strategies of Epipelagic Copepods in the Southern Ocean. J. Mar. Syst. 1998, 15, 289–311. [Google Scholar] [CrossRef]
- Turner, J.T. The Importance of Small Planktonic Copepods and Their Roles in Pelagic Marine Food Webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Edwards, M.; Richardson, A.J. Impact of Climate Change on Marine Pelagic Phenology and Trophic Mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate Change and Marine Plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Marshall, G.J. Climate Change in the Polar Regions; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change). Climate Change 2013: Executive Summary; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Atkinson, A. Subantarctic Copepods in an Oceanic, Low Chlorophyll Environment: Ciliate Predation, Food Selectivity and Impact on Prey Populations. Mar. Ecol. Prog. Ser. 1996, 130, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Razouls, S.; Desreumaux, N.; Kouwenberg, J.; de Bovée, F. Diversity and Geographic Distribution of Marine Planktonic Copepods. Marine Planktonic Copepods. 2022. Available online: http://copepods.obs-banyuls.fr/en (accessed on 1 March 2022).
- Broyer, C.; de Koubbi, P. Biogeographic Atlas of the Southern Ocean 2014; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 290–296. [Google Scholar]
- Barton, A.D.; Pershing, A.J.; Litchman, E.; Record, N.R.; Edwards, K.F.; Finkel, Z.V.; Kiørboe, T.; Ward, B.A. The Biogeography of Marine Plankton Traits. Ecol. Lett. 2013, 16, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T. Global Imprint of Climate Change on Marine Life. Nat. Clim. Chang. 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Humphries, G.R.; Magness, D.R.; Huettmann, F. Machine Learning for Ecology and Sustainable Natural Resource Management; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive Habitat Distribution Models in Ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Moilanen, A.; Leathwick, J.R.; Quinn, J.M. Spatial Prioritization of Conservation Management. Conserv. Lett. 2011, 4, 383–393. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Bonello, G.; Grillo, M.; Cecchetto, M.; Giallain, M.; Granata, A.; Guglielmo, L.; Pane, L.; Schiaparelli, S. Distributional Records of Ross Sea (Antarctica) Planktic Copepoda from Bibliographic Data and Samples Curated at the Italian National Antarctic Museum (MNA): Checklist of Species Collected in the Ross Sea Sector from 1987 to 1995. ZooKeys 2020, 969, 1. [Google Scholar] [CrossRef]
- Carli, A.; Feletti, M.; Pane, L. Zooplankton Biomass and Copepod Abundance of Terra Nova Bay, Ross Sea Antarctic Campaign 1994/1995). J. Mar. Sci. Eng. 2002, 8, 51–55. [Google Scholar]
- Carli, A.; Mariottini, G.L.; Pane, L. Contribution to the Study of Copepods Collected in Terra Nova Bay (Ross Sea). Nat. Sc. Com. Ant. Ocean. Camp. 1990, 88, 129–167. [Google Scholar]
- Carli, A.; Pane, L.; Stocchino, C. Planktonic Copepods in Terra Nova Bay (Ross Sea): Distribution and Relationship with Environmental Factors. In Ross Sea Ecology; Springer: Berlin/Heidelberg, Germany, 2000; pp. 309–321. [Google Scholar]
- Carli, A.; Feletti, M.; Mariottini, G.L.; Pane, L. Contribution to the Study of Copepods Collected during the Italian Oceanographic Campaign in Antarctica 1989–90. 1992, pp. 179–210. Available online: https://www.coldregions.org/vufind/Record/193046 (accessed on 1 March 2022).
- Pane, L.; Feletti, M.; Francomacaro, B.; Mariottini, G.L. Summer Coastal Zooplankton Biomass and Copepod Community Structure near the Italian Terra Nova Base (Terra Nova Bay, Ross Sea, Antarctica). J. Plankton Res. 2004, 26, 1479–1488. [Google Scholar] [CrossRef] [Green Version]
- Sertorio, T.Z.; Licandro, P.; Ossola, C.; Artegiani, A. Copepod Communities in the Pacific Sector of the Southern Ocean in Early Summer. In Ross Sea Ecology; Springer: Berlin/Heidelberg, Germany, 2000; pp. 291–307. [Google Scholar]
- Zunini Sertorio, T.; Salemi Picone, P.; Bernat, P.; Cattini, E.; Ossola, C. Copepods Collected in Sixteen Stations during the Italian Antarctic Expedition 1987–1988. Natl. Sci. Com. Antarct. Ocean. Camp. 1990, 1988, 67–125. [Google Scholar]
- Zunini Sertorio, T.; Licandro, P.; Ricci, F.; Giallain, M.; Antarctica, N.S.C. for A Study on Ross Sea Copepods. Nat. Sc. Com. Ant. Ocean. Camp. 1992, 88, 217–246. [Google Scholar]
- Hoshiai, T.; Tanimura, A.; Watanabe, K. Ice Algae as Food of an Antarctic Ice-Associated Copepod, Paralabidocera Antarctica (IC Thompson). In Proceedings of the NIPR Symposium on Polar Biology, Tokyo, Japan, 24–26 November 1987; Volume 1, p. 3. [Google Scholar]
- Arashkevich, Y.G. The Food and Feeding of Copepods in the Northwestern. Pacific. Oceanol. 1969, 9, 695–709. [Google Scholar]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; Ray Society: London, UK, 2004. [Google Scholar]
- Michels, J.; Schnack-Schiel, S.B. Feeding in Dominant Antarctic Copepods—Does the Morphology of the Mandibular Gnathobases Relate to Diet? Mar. Biol. 2005, 146, 483–495. [Google Scholar] [CrossRef]
- Yu, J.; Tian, J.; Yang, G. Ingestion, Fecundity and Population Growth of Harpacticus Sp.(Harpacticoida, Copepod) Fed on Five Species of Algae. Aquac. Res. 2017, 48, 2209–2220. [Google Scholar] [CrossRef]
- Nishida, S.; Ohtsuka, S. Specialized Feeding Mechanism in the Pelagic Copepod Genus Heterorhabdus (Calanoida: Heterorhabdidae), with Special Reference to the Mandibular Tooth and Labral Glands. Mar. Biol. 1996, 126, 619–632. [Google Scholar] [CrossRef]
- Ohtsuka, S.; Soh, H.Y.; Nishida, S. Evolutionary Switching from Suspension Feeding to Carnivory in the Calanoid Family Heterorhabdidae (Copepoda). J. Crustacean Biol. 1997, 17, 577–595. [Google Scholar] [CrossRef]
- Tseng, L.-C.; Dahms, H.-U.; Chen, Q.-C.; Hwang, J.-S. Copepod Feeding Study in the Upper Layer of the Tropical South China Sea. Helgol. Mar. Res. 2009, 63, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, M.L.; Pájaro, M.; Martos, P.; Esnal, G.B.; Sabatini, M.; Capitanio, F.L. Potential Zooplankton Preys (Copepoda and Appendicularia) for Engraulis Anchoita in Relation to Early Larval and Spawning Distributions in the Patagonian Frontal System (SW Atlantic Ocean). Sci. Mar. 2012, 76, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Sameoto, D.D.; Jaroszynski, L.O.; Fraser, W.B. BIONESS, a New Design in Multiple Net Zooplankton Samplers. Can. J. Fish. Aquat. Sci. 1980, 37, 722–724. [Google Scholar] [CrossRef]
- Guglielmo, L.; Costanzo, G.; Zagami, G.; Manganaro, A.; Arena, G. Zooplankton Ecology in the Southern Ocean. Data Rep. Part II Natl. Sci. Comm. Antarct. Genova 1992, 969, 30–468. [Google Scholar]
- Guglielmo, L.; Costanzo, G.; Manganaro, A.; Zagami, G. Spatial and Vertical Distribution of Zooplanktonic Communities in the Terra Nova Bay (Ross Sea). Nat. Sci. Antarct. Ocean. Campaign. 1990, 1988, 257–398. [Google Scholar]
- Team: QGIS Geographic Information System; Open Source Geospat. Found Proj. 2020. Available online: https://qgis.org/en/site/ (accessed on 1 March 2022).
- R Core Team. Rf. R: A Language and Environment for Statistical Computing; R Foundation: Vienna, Austria, 2013. [Google Scholar]
- Roth, G.; Matsuoka, K.; Skoglund, A.; Melvær, Y.; Tronstad, S. Quantarctica: A Unique, Open, Standalone GIS Package for Antarctic Research and Education. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 8–13 April 2017; p. 1973. [Google Scholar]
- Brown, M. Ocean Data View 4.0. Oceanography 1998, 11, 19–21. [Google Scholar] [CrossRef] [Green Version]
- Hardy, S.M.; Lindgren, M.; Konakanchi, H.; Huettmann, F. Predicting the Distribution and Ecological Niche of Unexploited Snow Crab (Chionoecetes opilio) Populations in Alaskan Waters: A First Open-Access Ensemble Model. Integr. Comp. Biol. 2011, 51, 608–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meißner, K.; Fiorentino, D.; Schnurr, S.; Arbizu, P.M.; Huettmann, F.; Holst, S.; Brix, S.; Svavarsson, J. Distribution of Benthic Marine Invertebrates at Northern Latitudes―An Evaluation Applying Multi-Algorithm Species Distribution Models. J. Sea Res. 2013, 85, 241–254. [Google Scholar] [CrossRef]
- Huettmann, F.; Schmid, M. 9.1. Climate Change and Predictions of Pelagic Biodiversity Components. Biogeogr. Atlas South. Ocean. Sci. Comm. Antarct. Res. Camb. 2014, 16, 470–475. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Mi, C.; Huettmann, F.; Sun, R.; Guo, Y. Combining Occurrence and Abundance Distribution Models for the Conservation of the Great Bustard. PeerJ 2017, 5, e4160. [Google Scholar] [CrossRef] [Green Version]
- Pearce, J.; Ferrier, S. Evaluating the Predictive Performance of Habitat Models Developed Using Logistic Regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef] [Green Version]
- Swets, J.A. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffman, H.S. CCAMLR Fisheries: Challenges to Effective Conservation and Management. J. Int. Wildl. Law Policy 2009, 12, 180–189. [Google Scholar] [CrossRef]
- C-CAMLR-XXXV, Report of the thirty-fifth meeting of the Scientific Committee, Hobart, Australia, 17–21 October, Annex 6, 3.2, 3.7-3.9. 2016. CCAMLR CONSERVATION MEASURE 91-05 (2016) for the Ross Sea Region Marine Protected Area, Specifically, Addressing the Priorities of Annex 91-05/C. 2016. Available online: https://www.ccamlr.org/en/system/files/e-sc-xxxv.pdf (accessed on 1 March 2022).
- Senô, J.; Komaki, Y.; Takeda, A. Reports on the Biology of the” Umitaka-Maru” Expedition Plankton Collected by the” Umitaka-Maru” in the Antarctic and Adjacent Waters by Larva Net, with Special References to Copepoda. Mem. Natl. Inst. Polar Res. Spec. 1963, 40, 212–220. [Google Scholar]
- Hopkins, T.L. Zooplankton Standing Crop in the Pacific Sector of the Antarctic. Biol. Antarct. Seas 1971, 4, 347–362. [Google Scholar]
- Maruyama, T.; Toyoda, H.; Suzuki, S. Preliminary Report on the Biomass of Macroplankton and Micronekton Collected with a Bongo Net during the Umitaka Maru FIBEX Cruise. Trans. Tokyo Univ. Fish. Tokyo Suisan Daigaku Tokubetsu Kenkyu Hokoku 1982, 5, 125. [Google Scholar]
- Hutchinson, G.E. Concluding Remarks Cold Spring Harbor Symposia on Quantitative Biology. GS Search 1957, 22, 415–427. [Google Scholar]
- Drew, C.A.; Wiersma, Y.F.; Huettmann, F. Predictive Species and Habitat Modeling in Landscape Ecology: Concepts and Applications; Springer Science & Business Media: Berlin, Germany, 2010. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Leathwick, J.R.; Elith, J.; Francis, M.P.; Hastie, T.; Taylor, P. Variation in Demersal Fish Species Richness in the Oceans Surrounding New Zealand: An Analysis Using Boosted Regression Trees. Mar. Ecol. Prog. Ser. 2006, 321, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Huettmann, F.; Schmid, M. Publicly Available Open Access Data and Machine Learning Model-Predictions Applied with Open Source Gis for the Entire Antarctic Ocean: A first Meta-Analysis and Synthesis from 53 Charismatic Species. Horiz. Earth Sci. Res. 2014, 23, 23–33. [Google Scholar]
- Pinkerton, M.H.; Smith, A.N.; Raymond, B.; Hosie, G.W.; Sharp, B.; Leathwick, J.R.; Bradford-Grieve, J.M. Spatial and Seasonal Distribution of Adult Oithona Similis in the Southern Ocean: Predictions Using Boosted Regression Trees. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 469–485. [Google Scholar] [CrossRef]
- Voronina, N.; Kolosova, E.; Melnikov, I. Zooplankton Life under the Perennial Antarctic Sea Ice. Polar Biol. 2001, 24, 401–407. [Google Scholar]
- Hureau, J.-C. The Significance of Fish in the Marine Antarctic Ecosystems. Polar Biol. 1994, 14, 307–313. [Google Scholar] [CrossRef]
- Pinkerton, M.H.; Cummings, V.; Forman, J.; Brown, J.; Bury, S.J. Trophic Connections in the Ross Sea: Information from Stomach Contents Analysis and Stable Isotopes of Carbon and Nitrogen; Final Research Report for Ministry of Fisheries, Project IPY200701 Obj10; NIWA, Wellington, 2011, 18.
- Stevens, D.W.; Dunn, M.R.; Pinkerton, M.H.; Forman, J.S. Diet of Antarctic Toothfish (Dissostichus Mawsoni) from the Continental Slope and Oceanic Features of the Ross Sea Region, Antarctica. Antarct. Sci. 2014, 26, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Granata, A.; Zagami, G.; Vacchi, M.; Guglielmo, L. Summer and Spring Trophic Niche of Larval and Juvenile Pleuragramma Antarcticum in the Western Ross Sea, Antarctica. Polar Biol. 2009, 32, 369–382. [Google Scholar] [CrossRef]
- Williams, R. Trophic Relationships between Pelagic Fish and Euphausiids in Antarctic Waters. In Antarctic Nutrient Cycles and Food Webs; Springer: Berlin/Heidelberg, Germany, 1985; pp. 452–459. [Google Scholar]
- Culina, A.; Baglioni, M.; Crowther, T.W.; Visser, M.E.; Woutersen-Windhouwer, S.; Manghi, P. Navigating the Unfolding Open Data Landscape in Ecology and Evolution. Nat. Ecol. Evol. 2018, 2, 420–426. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Worms Aphia ID | Trophic Guild |
|---|---|---|---|
| Acartiidae | Paralabidocera antarctica (Thompson I.C., 1898) | 345234 | Phytoplankton feeder [34] |
| Aetideidae | Aetideopsis antarctica (Wolfenden, 1908) | 341236 | Phyto- and zooplankton feeder [35] |
| Aetideopsis minor (Wolfenden, 1911) | 254600 | Phyto- and zooplankton feeder [35] | |
| Gaetanus tenuispinus (Sars G.O., 1900) | 237965 | Phyto- and zooplankton feeder [35] | |
| Augaptilidae | Haloptilus ocellatus Wolfenden, 1905 | 342624 | Zooplankton feeder [36] |
| Calanidae | Calanoides acutus (Giesbrecht, 1902) | 342434 | Phytoplankton feeder [18,37] |
| Calanus propinquus Brady, 1883 | 342435 | Phytoplankton feeder [18,37] | |
| Clausocalanidae | Microcalanus pygmaeus (Sars G.O., 1900) | 104513 | Phyto- and zooplankton feeder [37] |
| Ctenocalanus vanus Giesbrecht, 1888 | 104510 | Phytoplankton feeder [37] | |
| Euchaetidae | Paraeuchaeta antarctica (Giesbrecht, 1902) | 344974 | Zooplankton feeder [37] |
| Paraeuchaeta exigua (Wolfenden, 1911) | 345051 | Zooplankton feeder [36] | |
| Paraeuchaeta similis (Wolfenden, 1908) | 345055 | Zooplankton feeder [36] | |
| Harpacticidae | Harpacticus furcifer Giesbrecht, 1902 | 293136 | Phytoplankton feeder [38] |
| Heterorhabdidae | Heterorhabdus austrinus Giesbrecht, 1902 | 343728 | Zooplankton feeder [39,40] |
| Lucicutiidae | Lucicutia ovalis (Giesbrecht, 1889) | 104609 | Phytoplankton feeder [41] |
| Metridinidae | Metridia gerlachei Giesbrecht, 1902 | 344689 | Phyto-, zooplankton [37] and suspension feeder [36] |
| Metridia curticauda Giesbrecht, 1889 | 104628 | Suspension feeder [36] | |
| Oithonidae | Oithona frigida Giesbrecht, 1902 | 344712 | Phytoplankton feeder [42] |
| Oithona similis Claus, 1866 | 106656 | Phytoplankton feeder [42] | |
| Oncaeidae | Oncaea curvata Giesbrecht, 1902 | 344719 | Zooplankton feeder [36] |
| Triconia conifera (Giesbrecht, 1891) | 128957 | Zooplankton feeder [36] | |
| Rhincalanidae | Rhincalanus gigas Brady, 1883 | 220837 | Phyto- and zooplankton feeder [37] |
| Scolecitrichidae | Scolecithricella minor (Brady, 1883) | 104811 | Phyto- and zooplankton feeder [36] |
| Racovitzanus antarcticus Giesbrecht, 1902 | 104784 | Phyto- and zooplankton feeder [36] | |
| Scaphocalanus subbrevicornis (Wolfenden, 1911) | 104802 | Phyto- and zooplankton feeder [36] | |
| Stephidae | Stephos longipes Giesbrecht, 1902 | 345365 | Phytoplankton feeder [37] |
| Algorithm | ROC (%) Range | Class |
|---|---|---|
| TreeNet | 0–49 | Poor |
| RandomForest | 50–70 | Good |
| Ensemble | 71–100 | Very good |
| Sample Size (# of Occurrences) | TreeNet ROC (%) | RandomForest ROC (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | 0–750 [m] | 0–10 [m] | 11–70 [m] | 71–750 [m] | 0–750 [m] | 0–10 [m] | 11–70 [m] | 71–750 [m] | 0–750 [m] | 0–10 [m] | 11–70 [m] | 71–750 [m] |
| A. antarctica | 30 | 0 | 0 | 30 | 83 | 47 | ||||||
| A. minor | 1 | 0 | 0 | 1 | ||||||||
| C. acutus | 534 | 71 | 144 | 319 | 66 | 70 | 69 | 67 | 64 | 60 | 64 | 59 |
| C. propinquus | 242 | 28 | 55 | 159 | 66 | 73 | 73 | 60 | 63 | 70 | 69 | 55 |
| C. vanus | 125 | 8 | 22 | 95 | 82 | 83 | 84 | 76 | 75 | 58 | 76 | 71 |
| G. tenuispinus | 2 | 0 | 0 | 2 | ||||||||
| H. ocellatus | 5 | 0 | 2 | 3 | 73 | 87 | ||||||
| H. furcifer | 1 | 1 | 0 | 0 | ||||||||
| H. austrinus | 6 | 3 | 0 | 3 | 78 | 88 | ||||||
| L. ovalis | 3 | 2 | 0 | 1 | 49 | 38 | ||||||
| M. curticauda | 7 | 1 | 1 | 5 | 83 | 94 | ||||||
| M. gerlachei | 579 | 58 | 104 | 417 | 67 | 65 | 58 | 66 | 66 | 60 | 53 | 65 |
| M. pygmaeus | 1 | 0 | 0 | 1 | ||||||||
| O. frigida | 68 | 1 | 3 | 64 | 81 | 79 | ||||||
| O. similis | 231 | 55 | 52 | 124 | 84 | 84 | 87 | 79 | 79 | 78 | 80 | 73 |
| O. curvata | 214 | 55 | 43 | 116 | 83 | 84 | 85 | 76 | 77 | 82 | 83 | 68 |
| P. antarctica | 185 | 8 | 11 | 166 | 72 | 52 | 89 | 63 | 65 | 8 | 52 | 59 |
| P. exigua | 119 | 8 | 5 | 106 | 80 | 75 | 73 | 68 | 53 | 66 | ||
| P. similis | 3 | 0 | 0 | 3 | 60 | 0 | ||||||
| P. antarctica | 24 | 19 | 1 | 4 | 91 | 62 | ||||||
| R. antarcticus | 53 | 13 | 1 | 39 | 93 | 91 | ||||||
| R. gigas | 32 | 8 | 12 | 12 | 94 | 81 | 89 | 96 | 93 | 82 | 84 | 95 |
| S. subbrevicornis | 9 | 4 | 0 | 5 | 91 | 93 | ||||||
| S. minor | 4 | 0 | 0 | 4 | ||||||||
| S. longipes | 37 | 3 | 1 | 33 | 85 | 77 | ||||||
| T. conifera | 19 | 0 | 0 | 19 | 96 | 80 | ||||||
| Other copepods | 9 | 1 | 0 | 8 | 84 | 42 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grillo, M.; Huettmann, F.; Guglielmo, L.; Schiaparelli, S. Three-Dimensional Quantification of Copepods Predictive Distributions in the Ross Sea: First Data Based on a Machine Learning Model Approach and Open Access (FAIR) Data. Diversity 2022, 14, 355. https://doi.org/10.3390/d14050355
Grillo M, Huettmann F, Guglielmo L, Schiaparelli S. Three-Dimensional Quantification of Copepods Predictive Distributions in the Ross Sea: First Data Based on a Machine Learning Model Approach and Open Access (FAIR) Data. Diversity. 2022; 14(5):355. https://doi.org/10.3390/d14050355
Chicago/Turabian StyleGrillo, Marco, Falk Huettmann, Letterio Guglielmo, and Stefano Schiaparelli. 2022. "Three-Dimensional Quantification of Copepods Predictive Distributions in the Ross Sea: First Data Based on a Machine Learning Model Approach and Open Access (FAIR) Data" Diversity 14, no. 5: 355. https://doi.org/10.3390/d14050355