
Diversity of Periphytic Chironomidae on Different Substrate Types in a Floodplain Aquatic Ecosystem

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
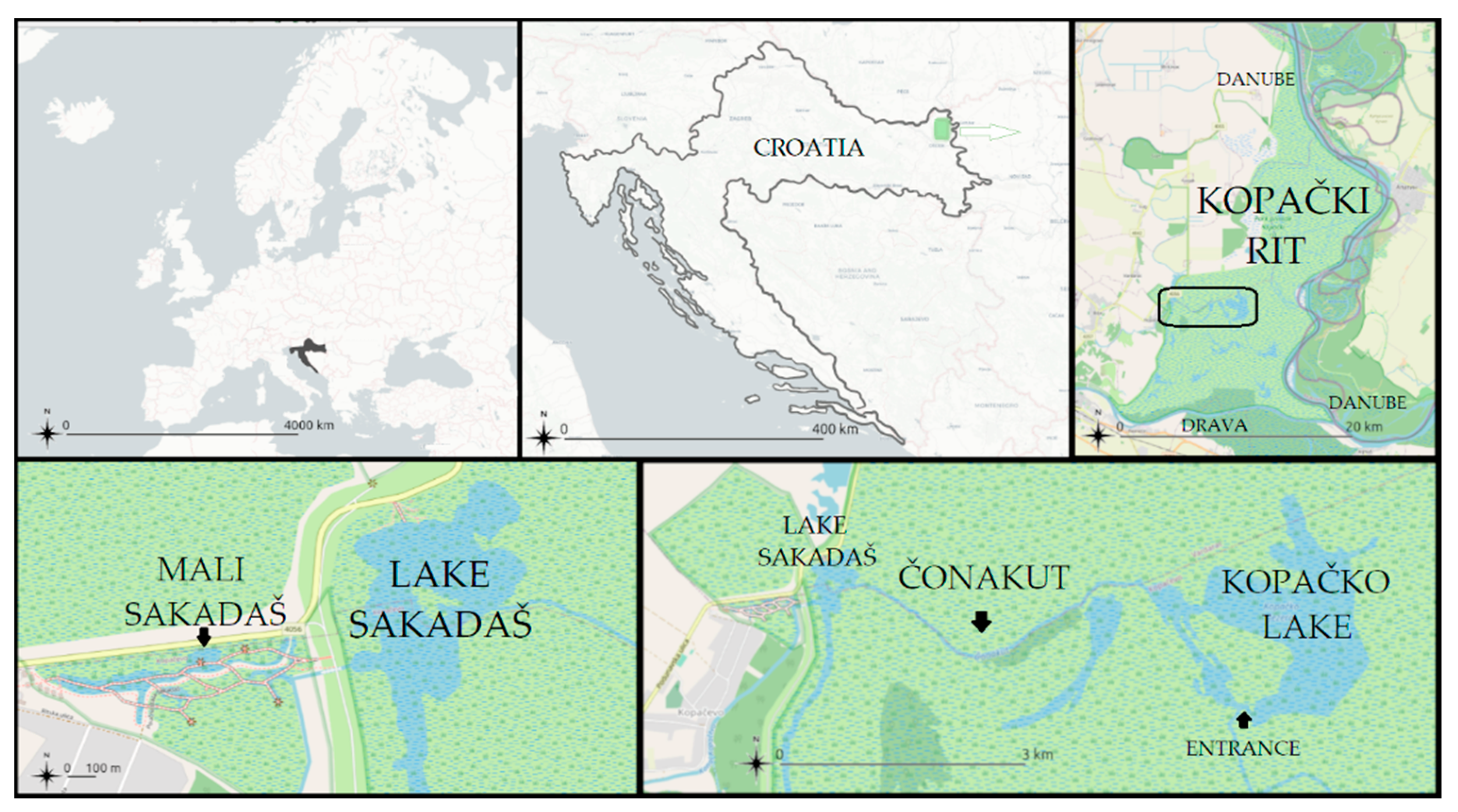
2.1. Study Area
2.2. Sampling Strategies
2.3. Periphyton
2.4. Statistical Analysis
3. Results
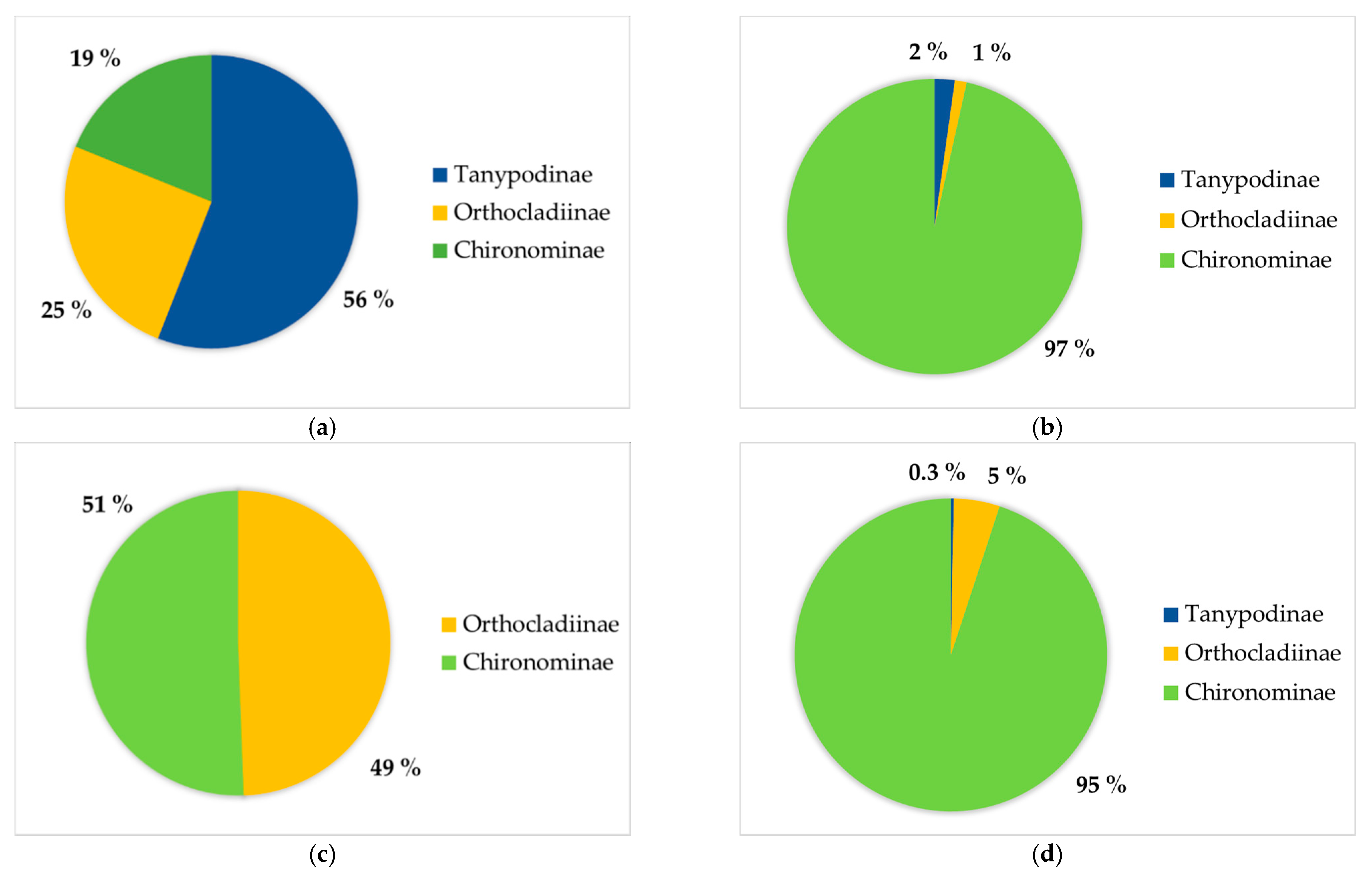
3.1. Diversity of Periphytic Chironomidae
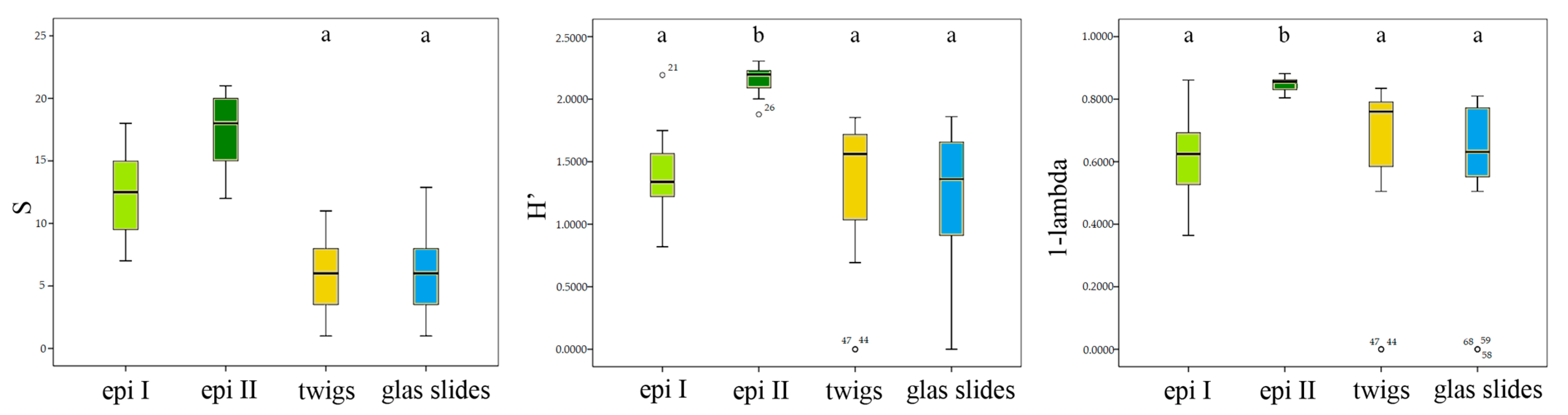
3.2. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Junk, W.; Bayley, P.; Sparks, R. The Food Pulse Concept in River-Foodplain Systems; Canadian Special Publication of Fisheries and Aquatic Science: Ottawa, ON, Canada, 1989; pp. 110–127. [Google Scholar]
- Tockner, K.; Malard, F.; Ward, J.V. An extension of the flood pulse concept. Hydrol. Process. 2000, 14, 2861–2883. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Agostinho, A.A.; Hahn, N.S. The Upper Paraná River and Its Floodplain: Physical Aspects, Ecology and Conservation; Backhuys Publishers: Leiden, The Netherlands, 2004; p. 393. [Google Scholar]
- Amoros, C.; Bornette, G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw. Biol. 2002, 47, 761–776. [Google Scholar] [CrossRef]
- Karaus, U.; Larsen, S.; Guillong, H.; Tockner, K. The contribution of lateral aquatic habitats to insect diversity along river corri-dors in the Alps. Landsc. Ecol. 2013, 28, 1755–1767. [Google Scholar] [CrossRef]
- Sommerwerk, N.; Bloesch, J.; Baumgartner, C.; Bittl, T.; Čerba, D.; Csányi, B.; Davideanu, G.; Dokulil, M.; Frank, G.; Grecu, I.; et al. The Danube River Basin. In Rivers of Europe, 2nd ed.; Tockner, K., Zarfl, C., Robinson, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 81–180. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J.A. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef] [Green Version]
- Opperman, J.J.; Galloway, G.E.; Duvail, S. The Multiple Benefits of River—Floodplain Connectivity for People and Biodiversity. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: London, UK, 2013; pp. 144–160. [Google Scholar] [CrossRef]
- Jakubínský, J.; Prokopova, M.; Raška, P.; Salvati, L.; Bezak, N.; Cudlín, O.; Cudlín, P.; Purkyt, J.; Vezza, P.; Camporeale, C.; et al. Managing Floodplains Using Nature-Based Solutions to Support Multiple Ecosystem Functions and Services; Wiley Interdisciplinary Reviews: Water; Wiley: New York, NY, USA, 2021; Volume 8, p. e1545. [Google Scholar] [CrossRef]
- Moss, B. Ecology of Freshwaters: A View for the Twenty-First Century, 4th ed.; John Wiley & Sons Ltd.: West Sussex, UK, 2010; p. 470. [Google Scholar]
- Armitage, P.D.; Cranston, P.S.; Pinder, L.C.V. The Chironomidae: Biology and Ecology of Non-Biting Midges; Chapman & Hall: London, UK, 1995; p. 572. [Google Scholar]
- Ferrington, L.C. Global diversity of non-biting midges (Chironomidae; Insecta-Diptera) in freshwater. In Freshwater Animal Diversity Assessment. Developments in Hydrobiology, Volume 198; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 447–455. [Google Scholar] [CrossRef]
- Vallenduuk, H.J. Chironomini Larvae of Western European Lowland (Diptera: Chironomidae). Keys with Notes to the Species; Eric Mauch Verlag: Dinkelscherben, Germany, 2017; p. 216. [Google Scholar]
- Andersen, T.; Baranov, V.; Hagenlund, L.K.; Ivković, M.; Kvifte, G.M.; Pavlek, M. Blind Flight? A New Troglobiotic Orthoclad (Diptera, Chironomidae) from the Lukina Jama-Trojama Cave in Croatia. PLoS ONE 2016, 11, e0152884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagna, M.; Urbanelli, S.; Rossaro, B. The species of the genus Diamesa (Diptera, Chironomidae) known to occur in Italian Alps and Apennines. Zootaxa 2016, 4193, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Čerba, D.; Hamerlík, L. Fountains—Overlooked Small Water Bodies in the Urban Areas. In Small Water Bodies of the Western Balkans; Pešić, V., Milošević, D., Miliša, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 73–91. [Google Scholar] [CrossRef]
- Rosenberg, D.M. Freshwater biomonitoring and Chironomidae. Neth. J. Aquat. Ecol. 1992, 26, 101–122. [Google Scholar] [CrossRef]
- Milošević, D.; Mančev, D.; Čerba, D.; Stojković Piperac, M.; Popović, N.; Atanacković, A.; Đuknić, J.; Simić, V.; Paunović, M. The potential of chironomid larvae-based metrics in the bioassessment of non-wadeable rivers. Sci. Total Environ. 2018, 616–617, 472–479. [Google Scholar] [CrossRef]
- Lencioni, V.; Rossaro, B. Microdistribution of chironomids (Diptera: Chironomidae) in Alpine streams: An autoecological perspective. Hydrobiologia 2005, 533, 61–76. [Google Scholar] [CrossRef]
- Tarkowska-Kukuryk, M. Environmental Drivers of Macroinvertebrate Assemblages within Peat Pool Habitat-Implication for Bioassessment. Water 2021, 13, 2369. [Google Scholar] [CrossRef]
- Čerba, D.; Mihaljević, Z.; Vidaković, J. Colonisation trends, community and trophic structure of chironomid larvae (Chironomidae: Diptera) in a temporal phytophylous assemblage. Fundam. Appl. Limnol. 2011, 179, 203–214. [Google Scholar] [CrossRef]
- Prejs, A.; Koperski, P.; Prejs, K. Food-web manipulation in a small, eutrophic Lake Wirbel, Poland: The effect of replacement of key predators on epiphytic fauna. Hydrobiologia 1997, 342, 377–381. [Google Scholar] [CrossRef]
- Moller Pillot, H.K.M. Chironomidae Larvae. Biology and Ecology of the Chironomini; KNNV Publishing: Zeist, The Netherlands, 2009; p. 270. [Google Scholar]
- Moller Pilot, H.K.M. Chironomidae Larvae. Vol. 3. Orthocladiinae: Biology and Ecology of the Aquatic Orthocladiinae; KNNV Publishing: Zeist, The Netherlands, 2013; p. 312. [Google Scholar] [CrossRef]
- Toth, M.; Móra, A.; Kiss, A.; Dévai, G.Y. Chironomid Communities in Different Vegetation Types in a Backwater Nagy-Morotva of the Active Floodplain of River Tisza, Hungary; Boletim Municipal: Margem, Portugal, 2008; pp. 169–175. [Google Scholar]
- Keiper, J.B.; Espeland, E.M. Spatial distribution and larval behavior of Glyptotendipes lobiferus (Diptera: Chironomidae). Hydrobiologia 2000, 427, 129–133. [Google Scholar] [CrossRef]
- Coe, H.J.; Kiffney, P.M.; Pess, G.R. A Comparison of Methods to Evaluate the Response of Periphyton and Invertebrates to Wood Placement in Large Pacific Coastal Rivers. Northwest Sci. 2006, 80, 298–307. [Google Scholar]
- Braccia, A.; Batzer, D.P. Invertebrates associated with woody debris in a southeastern U.S. forested floodplain wetland. Wetlands 2001, 21, 18–31. [Google Scholar]
- Schneider, K.N.; Winemiller, K.O. Structural complexity of woody debris patches influences fish and macroinvertebrate species richness in a temperate floodplain-river system. Hydrobiologia 2008, 610, 235–244. [Google Scholar] [CrossRef]
- Bazzanti, M.; Coccia, C.; Dowgiallo, M.G. Microdistribution of macroinvertebrates in a temporary pond of Central Italy: Taxonomic and functional analyses. Limnologica 2010, 40, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J. An example of relationships between macrophytes, macroinvertebrates and their food resources in a shallow euthrophic lake. Hydrobiologia 1996, 339, 27–36. [Google Scholar] [CrossRef]
- Bogut, I.; Vidaković, J.; Palijan, G.; Čerba, D. Bentic macroinvertebrates associated with four species of macrophytes. Biologia 2007, 62, 600–606. [Google Scholar] [CrossRef]
- De Camargo, N.S.J.; Lehun, A.L.; Rosa, J.; de Deus Bueno-Krawczyk, A.C. Colonization of benthic invertebrates on artificial and natural substrate in a Neotropical lotic environment in Southern Brazil. Acta. Sci. Biol. Sci. 2019, 41, e45872. [Google Scholar] [CrossRef]
- Preiner, S.; Weigelhofer, G.; Funk, A.; Hohensinner, S.; Reckendorfer, W.; Schiemer, F.; Hein, T. Danube Floodplain Lobau. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer: Cham, Switzerland, 2018; Volume 8, pp. 491–506. [Google Scholar] [CrossRef] [Green Version]
- Vidaković, J.; Turković Čakalić, I.; Stević, F.; Čerba, D. The influence of different hydrological conditions on periphytic invertebrate communities in a Danubian floodplain. Fundam. Appl. Limnol. 2012, 181, 59–72. [Google Scholar] [CrossRef]
- Mihaljević, M.; Žuna Pfeiffer, T.; Vidaković, J.; Špoljarić, D.; Stević, F. The importance of microphytic composition on coarse woody debris for nematode colonization: A case study in a fluvial floodplain environment. Biodivers. Conserv. 2015, 24, 1711–1727. [Google Scholar] [CrossRef]
- Schmid, P.E. A Key to the Larval Chironomidae and Their Instars from Austrian Danube Region Streams and Rivers. Part I: Diamesinae, Prodiamesinae and Orthocladiinae; Wasser und Abwasser, Suppl. 3; Fed. Inst. for Water Quality: Vienna, Austria, 1993; p. 514. [Google Scholar]
- Vallenduuk, H.J.; Moller Pillot, H.K.M. Chirnomidae Larvae, Vol. 1: Tanypodinae; KNNV Publishing Company: Zeist, The Netherlands, 2007. [Google Scholar]
- Bitušík, P. Priručka na Určovanie Lariev Pakomarov (Diptera: Chironomidae) Slovenska. Čast’ I. Buconomyinae, Diamesinae, Prodiamesinae a Orthocladiinae; Techn. Univ. vo Zvolene, Fak. Ekol. Environm, Katedra Biologie: Zvolen, Slovakia, 2000; pp. 1–133. [Google Scholar]
- Bitušik, P.; Hamerlik, L. Príručka na Určovanie Lariev Pakomárov (Diptera: Chironomidae) Slovenska. Časť 2. Tanypodinae. (Identification Key for Chironomidae of Slovakia. Part 2. Tanypodinae); Belianum, Vydavatelstvo Univerzity Matej Bela v Banskej Bystrici: Banská Bystrica, Slovakia, 2014; p. 96. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Pinder, L.C.V. The habitats of chironomid larvae. In The Chironomidae: Biology and Ecology of Non-Biting Midges; Armitage, P.D., Cranston, P.S., Pinder, L.C.V., Eds.; Chapman & Hall: London, UK, 1995; pp. 107–135. [Google Scholar]
- Tan, C.; Sheng, T.; Wang, L.; Mbao, E.; Gao, J.; Wang, B. Water-level fluctuations affect the alpha and beta diversity of macroinvertebrates in Poyang Lake, China. Fundam. Appl. Limnol. 2021, 194, 321–334. [Google Scholar]
- Larsen, S.; Karaus, U.; Claret, C.; Sporka, F.; Hamerlík, L.; Tockner, K. Flooding and hydrologic connectivity modulate community assembly in a dynamic river-floodplain ecosystem. PLoS ONE 2019, 14, e0213227. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, B.; Dolédec, S.; Paillex, A.; Arscott, D.B.; Sheldon, F.; Zilli, F.; Mérigoux, S.; Castella, E.; Comín, F.A. Response of benthic macroinvertebrates to gradients in hydrological connectivity: A comparison of temperate, subtropical, Mediterranean and semiarid river floodplains. Freshw. Biol. 2014, 59, 630–648. [Google Scholar] [CrossRef] [Green Version]
- Čerba, D.; Mihaljević, Z.; Vidaković, J. Colonisation of temporary macrophyte substratum by midges (Chironomidae: Diptera). Ann. Limnol. Int. J. Lim. 2010, 46, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.J.; Ricciardi, A. Epiphytic macroinvertebrate communities on Eurasian watermilfoil (Myriophyllum spicatum) and native milfoils Myriophyllum sibericum and Myriophyllum alterniflorum in eastern North America. Can. J. Fish. Aquat. Sci. 2009, 66, 18–30. [Google Scholar]
- Čerba, D.; Milošević, D.J.; Turković Čakalić, I.; Ergović, V.; Koh, M.; Vuković, A. Functional role of chironomid larvae (Chironomidae, Diptera) within a Danube floodplain. In Proceedings of the 8th Central European Dipterological Conference, Kežmarské Žľaby, High Tatra Mountains, Slovakia, 28–30 September 2015. [Google Scholar]
- Tank, J.L.; Winterbourn, M.J. Microbial activity and invertebrate colonization of wood in a New Zealand forest stream. N. Z. J. Mar. Freshwat. Res. 1996, 30, 271–280. [Google Scholar]
- Moog, O. Fauna Aquatica Austriaca. Wasserwirtschaftskataster, Bundesministerium fur Land-und Forstwirtschaft, Umwelt und Wasserwirtsdhaft; FAA: Vienna, Austria, 2002. [Google Scholar]
- Spänhoff, B.; Kaschek, N.; Meyer, E.I. Laboratory investigation on community composition, emergence patterns and biomass of wood-inhabiting Chironomidae (Diptera) from a sandy lowland stream in Central Europe (Germany). Aquat. Ecol. 2004, 38, 547–560. [Google Scholar]
- Wotton, R.S.; Armitage, P.D.; Aston, K.; Blackburn, J.H.; Hamburger, M.; Woodward, C.A. Colonization and emergence of midges (Chironomidae: Diptera) in slow sand filter beds. Neth. J. Aquat. Ecol. 1992, 26, 331–339. [Google Scholar]
- Stević, F.; Čerba, D.; Turković Čakalić, I.; Žuna Pfeiffer, T.; Vidaković, J.; Mihaljević, M. Interrelations between Dreissena polymorpha colonization and autotrophic periphyton development—A field study in a temperate floodplain lake. Fundam. Appl. Limnol. 2013, 183, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Tarkowska-Kukuryk, M. Periphytic algae as food source for grazing chironomids in a shallow phytoplankton-dominated lake. Limnologica 2013, 43, 254–264. [Google Scholar] [CrossRef]
- Tokeshi, M. On the evolution of commensalism in the Chironomidae. Freshw. Biol. 1993, 29, 481–489. [Google Scholar] [CrossRef]
- Ricciardi, A. Occurrence of chironomid larvae (Paratanytarsus sp.) as commensals of dreissenid mussels (Dreissena polymorpha and D. bugensis). Can. J. Zool. 1994, 72, 1159–1162. [Google Scholar] [CrossRef]
- Coffman, W.P.; Ferrington, L.C. Chironomidae. In An Introduction of Aquatic Insects of North America; Merrit, K.W., Cummins, R.W., Eds.; Kendall Hunt Publishing: Dubuque, Iowa, 1996; pp. 551–652. [Google Scholar]
- Rossaro, B.; Lencioni, V.; Boggero, A.; Marziali, L. Chironomids from Southern Alpine running waters: Ecology, biogeography. Hydrobiologia 2006, 562, 231–246. [Google Scholar] [CrossRef] [Green Version]
- Milošević, D.; Simić, V.; Stojković, M.; Čerba, D.; Mančev, D.; Petrović, A.; Paunović, M. Spatio-Temporal Pattern of the Chironomidae Community: Toward the Use of Non-Biting Midges in Bioassessment Programs. Aquat. Ecol. 2013, 47, 37–55. [Google Scholar] [CrossRef]
- European Water Framework Directive. 2000/60/EC; WFD: London, UK, 2000. [Google Scholar]
- Karr, J.R.; Chu, E.W. Restoring Life in Running Waters: Better Biological Monitoring; Island Press: Washington, DC, USA, 1999; p. 200. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa/Substrate | Epiphyton I | Epiphyton II | Twigs | Glass Slides | ||||
|---|---|---|---|---|---|---|---|---|
| Range | Average (N = 25) | Range | Average (N = 9) | Range | Average (N = 24) | Range | Average (N = 19) | |
| Tanypodinae | ||||||||
| Ablabesmyia (Ablabesmyia) longistyla Fittkau, 1962 | 0–1.96 | 0.29 | 0–0.46 | 0.05 | ||||
| Ablabesmyia (Ablabesmyia) monilis agg. | 0–1.39 | 0.06 | 0–2.16 | 0.32 | ||||
| Ablabesmyia spp. | 0–0.32 | 0.01 | 0–0.46 | 0.05 | ||||
| Conchapelopia agg. | 0–0.01 | 0.001 | ||||||
| Monopelopia tenuicalcar (Kieffer, 1918) | 27.81–79.49 | 55.96 | ||||||
| Tanypodinae non det. | 0–11.70 | 1.78 | 0–5.59 | 0.29 | ||||
| Orthocladiinae | ||||||||
| Chaetocladius spp. | 0–3.97 | 0.58 | ||||||
| Corynoneura gr. scutellata | 0–45.77 | 19.49 | 0–0.74 | 0.04 | ||||
| Corynoneura spp. | 0–1.49 | 0.23 | ||||||
| Cricotopus (Cricotopus) bicinctus (Meigen, 1818) | 0–0.20 | 0.01 | ||||||
| Cricotopus (Isocladius) intersectus agg. | 0–14.81 | 0.99 | 0–100 | 25.05 | 0–5.90 | 0.50 | ||
| Cricotopus (Isocladius) gr. sylvestris | 0–18.17 | 4.20 | 0–3.68 | 1.01 | 0–100 | 11.36 | 0–8.07 | 1.60 |
| Cricotopus spp. | 0–0.65 | 0.03 | 0–14.29 | 1.92 | 0–0.74 | 0.08 | ||
| Cricotopus/Orthocladius spp. | 0–58.33 | 4.63 | ||||||
| Nanocladius gr. dichromus | 0–0.65 | 0.03 | ||||||
| Psectrocladius (Psectrocladius) limbatellus (Holmgren, 1869) | 0–0.27 | 0.04 | ||||||
| Psectrocladius (Psectrocladius) gr. sordidellus | 0–1.59 | 0.07 | ||||||
| Orthocladiinae non det. | 0–100 | 6.39 | 0–50 | 3.00 | ||||
| Chironominae | ||||||||
| Chironomus (Chironomus) annularis agg. | 0–0.42 | 0.05 | ||||||
| Chironomus (Chironomus) luridus Strenzke, 1959 | 0–1.60 | 0.57 | ||||||
| Chironomus (Chironomus) plumosus agg. | 0–2.93 | 0.44 | 0–2.63 | 0.14 | ||||
| Chironomus (Chironomus) tentans Fabricius, 1805 | 0–0.91 | 0.10 | ||||||
| Chironomus (Lobochironomus) dorsalis Meigen, 1818 | 0–17.07 | 3.73 | ||||||
| Chironomus spp. | 0–2.47 | 0.44 | 0–29.07 | 13.56 | ||||
| Dicrotendipes lobiger (Kieffer, 1921) | 0–2.30 | 0.12 | 3.25–21.64 | 10.33 | 0–3.13 | 0.25 | 0–10.81 | 0.57 |
| Dicrotendipes modestus (Say, 1823) | 0–7.33 | 1.27 | ||||||
| Dicrotendipes nervosus (Staeger, 1839) | 0–1.85 | 0.14 | 0–50 | 6.07 | 0–56.72 | 9.76 | ||
| Dicrotendipes notatus (Meigen, 1818) | 0–0.21 | 0.02 | ||||||
| Dicrotendipes pulsus (Walker, 1856) | 0–0.95 | 0.06 | 0–20 | 0.97 | 0–0.57 | 0.03 | ||
| Dicrotendipes spp. | 0–5.56 | 0.22 | 0–2.71 | 1.33 | 0–2.38 | 0.10 | 0–2.63 | 0.14 |
| Endochironomus albipennis (Meigen, 1830) | 0–15.82 | 1.33 | 0–2 | 0.61 | 0–4 | 0.53 | 0–59.86 | 6.36 |
| Endochironomus tendens (Fabricius, 1775) | 0–5.37 | 0.77 | 1.22–9.81 | 4.73 | ||||
| Glyptotendipes (Glyptotendipes) barbipes (Staeger, 1839) | 0–4.18 | 0.22 | ||||||
| Glyptotendipes (Glyptotendipes) pallens agg. | 0–10.54 | 2.33 | 4.27–30.96 | 15.10 | 0–43.75 | 13.82 | 0–61.20 | 24.74 |
| Glyptotendipes (Glyptotendipes) paripes (Edwards, 1929) | 0–0.54 | 0.06 | ||||||
| Glyptotendipes spp. | 0–11.24 | 2.47 | 0–5.26 | 1.28 | ||||
| Kiefferulus (Kiefferulus) tendipediformis (Goetghebuer, 1921) | 0–10.74 | 3.54 | 0–2.92 | 1.43 | ||||
| Parachironomus gr. arcuatus | 0–1.03 | 0.05 | 0–3.24 | 0.36 | 0–60 | 15.20 | ||
| Parachironomus gr. frequens | 0–10.69 | 0.56 | ||||||
| Parachironomus varus (Goetghebuer, 1921) | 0–100 | 15.56 | ||||||
| Parachironomus spp. | 0–31.10 | 8.92 | ||||||
| Paratendipes nudisquama (Edwards, 1929) | 0–0.32 | 0.01 | ||||||
| Paratendipes spp. | 0–0.17 | 0.01 | ||||||
| Polypedilum (Pentapedilum) sordens (van der Wulp, 1875) | 0–7.39 | 1.98 | 0–4.50 | 1.09 | 0–20 | 2.96 | 0–21.63 | 4.76 |
| Polypedilum (Pentapedilum) uncinatum agg. | 0–3.75 | 0.62 | 0–0.54 | 0.06 | ||||
| Polypedilum (Polypedilum) nubeculosum (Meigen, 1804) | 0–2.17 | 0.16 | 0–2.63 | 0.18 | ||||
| Polypedilum (Polypedilum) pedestre (Meigen, 1830) | 0–18.07 | 3.13 | ||||||
| Polypedilum (Tripodura) scalaenum (Schrank, 1803) | 0–0.26 | 0.01 | ||||||
| Polypedilum (Uresipedilum) cultellatum Goetghebuer, 1931 | 0–2.25 | 0.16 | ||||||
| Polypedilum uncinatum agg./cultellatum | 0–0.54 | 0.06 | ||||||
| Polypedilum spp. | 0–14.46 | 2.65 | 0–3.73 | 0.70 | 0–50 | 2.51 | 0–50 | 3.20 |
| Paratanytarsus spp. | 0–5.37 | 0.99 | 2.26–23.13 | 8.81 | 0–50 | 5.15 | ||
| Tanytarsus spp. | 0–1.05 | 0.06 | 0–12.68 | 5.18 | ||||
| Chironominae non det. | 2.40–36.57 | 16.71 | 0–42.37 | 8.17 | 0–100 | 20.21 | ||
| Twigs | Glass Slides | epi I | epi II | |
|---|---|---|---|---|
| Twigs | ||||
| Glass slides | 0.392 | |||
| epi I | 0.848 | 0.844 | ||
| epi II | 0.602 | 0.360 | 0.999 |
| Contribution (%) | |
|---|---|
| tw and gs | Average dissimilarity = 78.04 |
| Cricotopus intersectus agg. | 12.98 |
| Chironominae non det. | 10.37 |
| Glyptotendipes pallens agg. | 9.91 |
| Monopelopia tenuicalcar | 18.45 |
| Corynoneura gr. scutellata | 10.14 |
| Cricotopus intersectus agg. | 9.83 |
| gs and epi I | Average dissimilarity = 88.53 |
| Monopelopia tenuicalcar | 18.79 |
| Corynoneura gr. scutellata | 10.24 |
| Chironominae non det. | 8.96 |
| tw and epi II | Average dissimilarity = 82.64 |
| Cricotopus intersectus agg. | 9.39 |
| Chironomus spp. | 7.32 |
| Parachironomus gr. arcuatus | 6.7 |
| gs and epi II | Average dissimilarity = 75.37 |
| Chironomus spp. | 8.13 |
| Dicrotendipes lobiger | 7.39 |
| Glyptotendipes pallens agg. | 7.13 |
| epi I and epi II | Average dissimilarity = 79.28 |
| Monopelopia tenuicalcar | 16.03 |
| Corynoneura gr. scutellata | 8.82 |
| Chironominae non det. | 8.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čerba, D.; Koh, M.; Vlaičević, B.; Turković Čakalić, I.; Milošević, D.; Stojković Piperac, M. Diversity of Periphytic Chironomidae on Different Substrate Types in a Floodplain Aquatic Ecosystem. Diversity 2022, 14, 264. https://doi.org/10.3390/d14040264
Čerba D, Koh M, Vlaičević B, Turković Čakalić I, Milošević D, Stojković Piperac M. Diversity of Periphytic Chironomidae on Different Substrate Types in a Floodplain Aquatic Ecosystem. Diversity. 2022; 14(4):264. https://doi.org/10.3390/d14040264
Chicago/Turabian StyleČerba, Dubravka, Miran Koh, Barbara Vlaičević, Ivana Turković Čakalić, Djuradj Milošević, and Milica Stojković Piperac. 2022. "Diversity of Periphytic Chironomidae on Different Substrate Types in a Floodplain Aquatic Ecosystem" Diversity 14, no. 4: 264. https://doi.org/10.3390/d14040264