
Genotyping-by-Sequencing Analysis Shows That Siberian Lindens Are Nested within Tilia cordata Mill


Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
4.1. Phylogeny of the T. cordata Clade
4.2. Natural History of Siberian Lindens
4.3. The Linden Specimen from Novosibirsk
4.4. Taxonomic Implications
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaripov, R.G.; Pashina, M.V. Tilia cordata Mill. In The Red Book of the Omsk Oblast, 2nd ed.; Sidorov, G.N., Ed.; Omsk Pedagogical State University: Omsk, Russia, 2015; p. 427. [Google Scholar]
- Krasnikov, A.A. Tilia cordata Mill. (1768). In The Red Book of the Novosibirsk Oblast. Animals, Plants, and Fungi, 3rd ed.; Marchenko, Y.Y., Ed.; Ministry of Natural Resources and Ecology of Novosibirsk Oblast: Novosibirsk, Russia, 2018; p. 427. [Google Scholar]
- Polozhiy, A.V. Tilia cordata Mill. In The Red Book of the Tomsk Oblast; Revushkin, A.S., Ed.; Tomsk University Press: Tomsk, Russia, 2002; p. 342. [Google Scholar]
- Krylov, P.N. Linden in the Piedmonts of Kuznetskii Alatau; Tomsk University Press: Tomsk, Russia, 1891; p. 156. [Google Scholar]
- Kuminova, A.V. On the modern state of the lime forest in the Kuznetskii Alatau. Proc. Tomsk Univ: 1951, 116, 181–186. [Google Scholar]
- Khlonov, Y.P. Linden and Linden Forests in West Siberia; Nauka Press: Novosibirsk, Russia, 1965; p. 40. [Google Scholar]
- Krapivkina, E.D. Tilia sibirica Bayer. In The Red Book of the Kemerovo Oblast. Rare and Threatened Species of Plants and Fungi, 2nd ed.; Kupriyanov, A.N., Ed.; Irbis: Kemerovo, Russia, 2012; Volume 1, p. 446. [Google Scholar]
- Revyakina, N.V. Tilia sibirica Bayer. In The Red Book of the Kemerovo Oblast. Rare and Threatened Species of Plants and Fungi; IPP Altay: Barnaul, Russia, 2006; Volume 1, p. 186. [Google Scholar]
- Krylov, P.N. Flora Sibiriae Occidentalis. Editio Secunda et Completa. Florae Altaicae et Provinciae Tomskiensis. Volumen VIII. Geraniaceae—Cornaceae; Botanical Section of Tomsk Society of Nature Explorers: Tomsk, Russia, 1935; pp. 1819–2087. [Google Scholar]
- Mallev, V.P. Family XCIV—Tiliaceae. In Flora URSS.; Shishkin, B.K., Bobrov, B.K., Eds.; Editio Academiae Scientarum URSS: Moscow, Russia, 1949; Volume XV, pp. 1–23. [Google Scholar]
- Vlasova, N.V. Family Tiliaceae. In Flora sibirieae Tomus 10, Geraniaceae—Cornaceae; Peshkova, G.A., Ed.; Nauka: Novosibirsk, Russia, 1996; pp. 65–66. [Google Scholar]
- Stepanov, N.V. Tilia nasczokinii (Tiliaceae); A new species from the neighbourhood of Krasnoyarsk. Bot. Zhurnal 1993, 78, 137–145. [Google Scholar]
- Koropachinskiy, I.Y.; Vstovskaya, T.N. Woody Plants of the Asian Part of Russia; SB RAS Press: Novosibirsk, Russia, 2002; p. 707. [Google Scholar]
- Logan, S.A.; Chytrý, M.; Wolff, K. Genetic diversity and demographic history of the Siberian lime (Tilia sibirica). Perspect. Plant Ecol. Evol. Syst. 2018, 33, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Semerikova, S.A.; Isakov, I.Y.; Semerikov, V.L. Chloroplast DNA Variation shed light on the history of lime tree (Tilia cordata s. l.) in the eastern part of the range. Russ. J. Genet. 2020, 56, 192–203. [Google Scholar] [CrossRef]
- Ekart, A.K.; Larionova, A.Y.; Kravchenko, A.N.; Semerikova, S.A.; Sedaeva, M.I. Genetic structure and differentiation of relict lime populations based on the analysis of variability of nuclear microsatellite loci. Russ. J. Genet. 2021, 57, 920–927. [Google Scholar] [CrossRef]
- Tsvelev, N.N. Genus Tilia L. In Conspect of the Flora of Eastern Europe; Tsvelev, N.N., Ed.; KMK Scientific Press: Saint-Petersburg, Russia, 2012; pp. 471–474. [Google Scholar]
- Shekhovtsov, S.V.; Shekhovtsova, I.N.; Peltek, S.E. Phylogeny of Siberian species of Carex sect. Vesicariae based on nuclear and plastid markers. Nord. J. Bot. 2012, 30, 343–351. [Google Scholar] [CrossRef]
- Wendler, N.; Mascher, M.; Nöh, C.; Himmelbach, A.; Scholz, U.; Ruge-Wehling, B.; Stein, N. Unlocking the secondary gene-pool of barley with next-generation sequencing. Plant Biotechnol. J. 2014, 12, 1122–1131. [Google Scholar] [CrossRef]
- Eaton, D.A.R.; Overcast, I. ipyrad: Interactive assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592–2594. [Google Scholar] [CrossRef]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Cariou, M.; Duret, L.; Charlat, S. Is RAD-seq suitable for phylogenetic inference? An in silico assessment and optimization. Ecol. Evol. 2013, 3, 846–852. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fineschi, S.; Salvini, D.; Taurchini, D.; Carnevale, S.; Vendramin, G.G. Chloroplast DNA variation of Tilia cordata (Tiliaceae). Can. J. For. Res. 2003, 33, 2503–2508. [Google Scholar] [CrossRef]
- Kuminova, A.V. Vegetation Cover of the Altai; Nauka Press: Novosibirsk, Russia, 1960; p. 450. [Google Scholar]
- Polozhiy, A.V.; Krapivkina, E.D. Relics of the Tertiary Broad-leaved Forests in the Flora of Siberia; Tomsk University Press: Tomsk, Russia, 1985; p. 158. [Google Scholar]
- Lashchinskiy, N.N.; Lashchinskaya, N.V. Higher vascular plants. In Flora of the Salair Ridge; Lashchinskiy, N.N., Ed.; GEO: Novosibirsk, Russia, 2007; pp. 155–250. [Google Scholar]
- Peshkova, G.A. On the accordance in development of mesophilous and xerophilous florae of the Baikalian Siberia in the Cenozoi. In The History of Vegetation Cover of North Asia; Malyshev, L.I., Ed.; Nauka Press: Novosibirsk, Russia, 1984; pp. 144–156. [Google Scholar]
- Arkhipov, S.A.; Volkova, V.S. Geological History, Landscapes and Climates of the Pleistocene in West Siberia; IGiG SO RAN: Novosibirsk, Russia, 1994; p. 106. [Google Scholar]
- Binney, H.; Edwards, M.; Macias-Fauria, M.; Lozhkin, A.; Anderson, P.; Kaplan, J.O.; Andreev, A.; Bezrukova, E.; Blyakharchuk, T.; Jankovska, V. Vegetation of Eurasia from the last glacial maximum to present: Key biogeographic patterns. Quat. Sci. Rev. 2017, 157, 80–97. [Google Scholar] [CrossRef]
- Tarasov, P.E.; Volkova, V.S.; Webb, T.; Guiot, J.; Andreev, A.A.; Bezusko, L.G.; Bezusko, T.V.; Bykova, G.V.; Dorofeyuk, N.I.; Kvavadze, E.V.; et al. Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia. J. Biogeogr. 2000, 27, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Amelin, I.I.; Blyakharchuk, T.A. Distribution of Siberian linden (Tilia sibirica Bayer) in Kemerovo oblast. Vestn. Tomsk. Gos. Univ. Biol. 2016, 2, 30–52. [Google Scholar] [CrossRef] [Green Version]
- Belova, V.A. Vegetation and Climate of the Late Cenozoic of the Southern Eastern Siberia; Nauka Press: Novosibirsk, Russia, 1985; p. 158. [Google Scholar]
- Volkova, V.S.; Belova, V.A. On a role of broad-leaved trees in vegetation of the Holocene in Siberia. In Paleopalynology of Siberia. Papers of the Soviet Palynologists to the V International Conference on Palynology (Cambridge, England, 1980); Saks, V.N., Ed.; Nauka Press: Moscow, Russia, 1980; pp. 112–117. [Google Scholar]
- Dubatolov, V.; Kosterin, O. Nemoral species of Lepidoptera (Insecta) in Siberia: A novel view on their history and the timing of their range disjunctions. Entomol. Fenn. 2000, 11, 141–166. [Google Scholar] [CrossRef] [Green Version]
- Bolikhovskaya, N.S.; Derevyanko, A.P.; Shun’kov, M.V. The fossil palynoflora, geological age, and climatostratigraphy of the earliest deposits of the Karama site (Early Paleolithic, Altai Mountains). Paleontol. J. 2006, 40, S558–S566. [Google Scholar] [CrossRef]
- Reverdatto, V.V. Main aspects of development of the post-Tertiary flora of Siberia. Sov. Bot. 1940, 2, 48–60. [Google Scholar]
- Grosset, G.E. The age of thermophylous relic flora of the broad-leaved forests of the Russian Plain, South Ural and Siberia with respect to the paleography of the Pleistocene and Holocene. Bulleten’ MOIP Biol. Div. 1962, 17, 94–109. [Google Scholar]
- Khlonov, Y.P. Siberian linden—Tilia sibirica Bayer. In Biological Foundations of Protection of Rare and Threatened Siberian Plants; Sobolevskaya, K.A., Ed.; Nauka: Novosibirsk, Russia, 1990; pp. 58–80. [Google Scholar]
- Novák, J.; Trotsiuk, V.; Sýkora, O.; Svoboda, M.; Chytrý, M. Ecology of Tilia sibirica in a continental hemiboreal forest, southern Siberia: An analogue of a glacial refugium of broad-leaved temperate trees? Holocene 2014, 24, 908–918. [Google Scholar] [CrossRef]
- Sedelnikov, V.P.; Banaev, E.V.; Chindyaeva, L.N. CSBG and the landscape architecture of Novosibirsk Scientific Centre. In The Nature of Akademgorodok, 50 Years Later; Zhimulev, I.F., Ed.; SB RAS Press: Novosibirsk, Russia, 2007; pp. 32–40. [Google Scholar]
- Krylov, G.V.; Salatova, N.G. Landscaping of Cities and Towns of West Siberia; Vnitoles: Novosibirsk, Russia, 1955; p. 56. [Google Scholar]
- Lashchinskiy, N.N.; Makunina, N.I.; Mal’tseva, T.V. A sketch of vegetation. In Ecosystem Dynamics of the Novosibirsk Academy Town; Zhimulev, I.F., Ed.; SB RAS Press: Novosibirsk, Russia, 2013; pp. 85–104. [Google Scholar]
- Williams, D.M.; Kociolek, J.P. Pursuit of a natural classification of diatoms: History, monophyly and the rejection of paraphyletic taxa. Eur. J. Phycol. 2007, 42, 313–319. [Google Scholar] [CrossRef]
- Moonlight, P.W.; Ardi, W.H.; Padilla, L.A.; Chung, K.-F.; Fuller, D.; Girmansyah, D.; Hollands, R.; Jara-Muñoz, A.; Kiew, R.; Leong, W.-C.; et al. Dividing and conquering the fastest-growing genus: Towards a natural sectional classification of the mega-diverse genus Begonia (Begoniaceae). Taxon 2018, 67, 267–323. [Google Scholar] [CrossRef]
- Mayr, E. Of what use are subspecies? Auk 1982, 99, 593–595. [Google Scholar]
- Galushko, A.I. Flora of the Northern Caucasus; Rostov University Press: Rostov, Russia, 1980; Volume 2, 352p. [Google Scholar]
- Zernov, A.S. Flora of the Northwestern Caucasus; KMK Scientific Press: Moscow, Russia, 2006; 664p. [Google Scholar]
- Murtazaliev, R.A. Conspect of the Flora of Dagestan; Epokha: Makhachkala, Russia, 2009; Volume 1, 320p. [Google Scholar]
- Zhu, Y.C. (Ed.) Plantae Medicinales China Boreali-Orientalis; Heilongjiang Science and Technology Publishing House: Harbin, China, 1989; 1300p. [Google Scholar]
- Govaerts, R.; Lughadha, E.N.; Black, N.; Turner, R.; Paton, A. The World Checklist of Vascular Plants, a continuously updated resource for exploring global plant diversity. Sci. Data 2021, 8, 215. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: http://powo.science.kew.org (accessed on 1 March 2022).



{kind=link}
{kind=link}
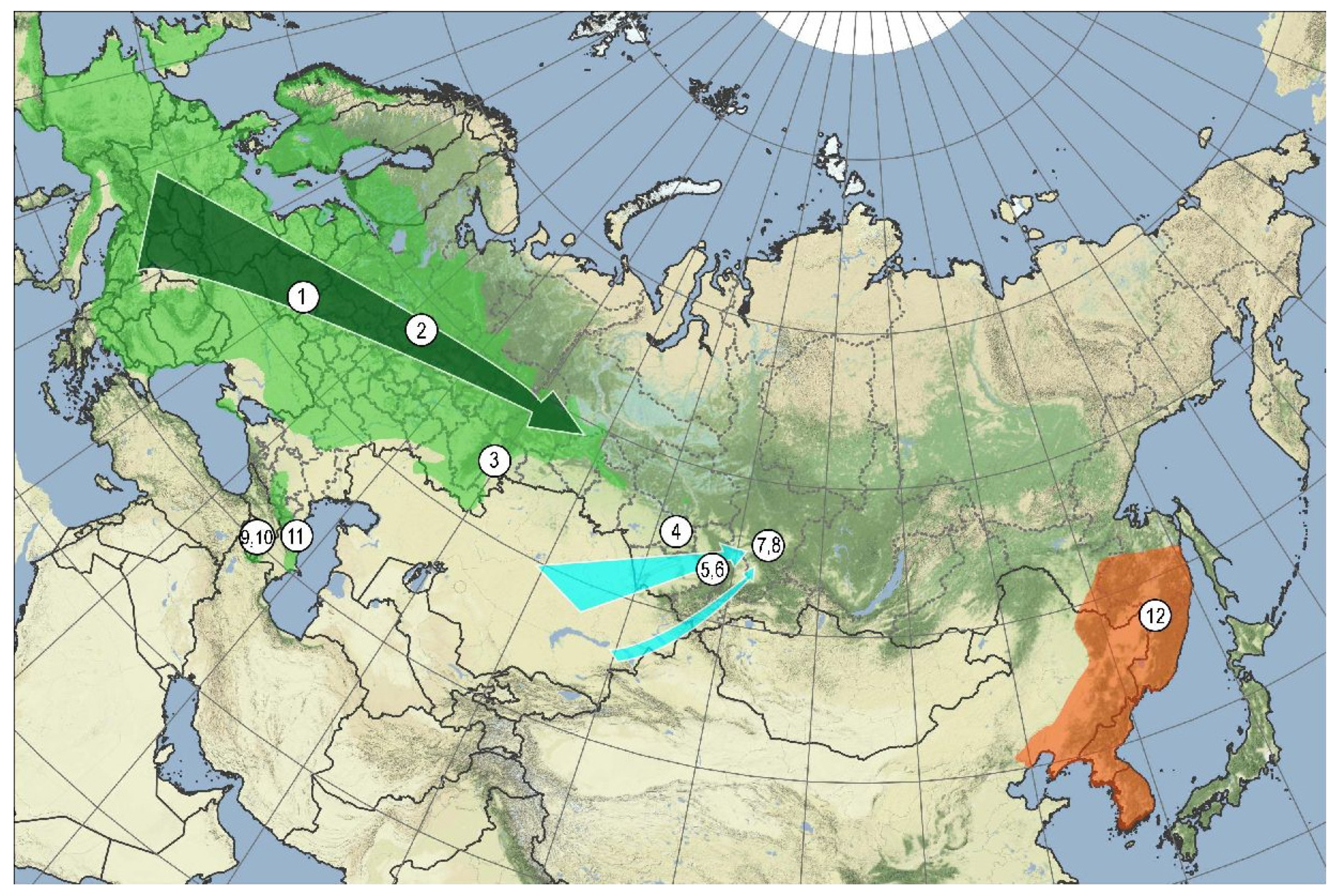
| No. | Species | Location |
|---|---|---|
| 1 | T. cordata cordata | Belarus, Krichev Natural Reserve, mixed forest, N53.643, E31.883 (NSK0089748) |
| 2 | -//- | Russia, Kostroma Oblast, Sudislavl’ District, Sledovo village, N57.76333 E41.2518368, 137 m a.s.l. (NSK0089749) |
| 3 | -//- | Russia, Chelyabinsk Oblast, Il’menskiy State Nature Reserve, slope of the Lake Bol’shoy Tatkul’ SW bank, mixed forest, N55.1893, E60.2870 (NSK0089750)) |
| 4 | Tilia cordata subsp. | Russia, Novosibirsk Oblast, Novosibirsk Academy Town, a lingen alley at Academician Lavrentyev avenue 10, N54.84788, E83.10825 (NSK0089751) |
| 5 | T. cordata nasczokinii | Russia, Krasnoyarskiy Kray, Krasnoyarsk, Kashtak Valley (the type locality of Tilia nasczokini), N55.9575, E92.7793 (NSK0089752) |
| 6 | -//- | Russia, Krasnoyarskiy Kray, Krasnoyarsk District, 3 km NE of Ust’-Mana village, Manskoe Zaymishche terrain, N55.9643, E92.5029 (NSK0089753) |
| 7 | T. cordata sibirica | Russia, Kemerovo Oblast, Novokuznetsk District, 4.5 km SE of Kuzedeevo village, Novokuznetsk District, a ridge along the Kondoma River right bank, N53.3204, E87.2533 (NSK0089754) |
| 8 | -//- | Russia, Kemerovo Oblast, 6 km ESE of Kuzedeevo village, the interfluve of the Bol’shoy and Malyy Tesh Rivers, N53.3281, E87.2806 (NSK0089755) |
| 9 | Tilia cordata undescr. subsp. | Armenia, Lori Province, Chkalov settlement, N40.928 E 44.626 (NSK0089756) |
| 10 | T. begoniifolia | Armenia, Tavush Province, Gosh village, N40.729, E45.002 (NSK0089757) |
| 11 | Tilia cordata undescr. subsp. | Russia, Dagestan Republic, Tabasaran District, 2.8 km WSW Kuzhnik village, beach forest margin, N41.9445 E47.7908 (NSK0089758) |
| 12 | T. amurensis | Russia, Khabarovskiy kray, Khabarovsk District, Bol’shekhekhtsyrskiy Nature Reserve, N48.88 E134.76 (NSK0089759) |
| -//- | T. taquetii | -//- (NSK0089760) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shekhovtsov, S.V.; Shekhovtsova, I.N.; Kosterin, O.E. Genotyping-by-Sequencing Analysis Shows That Siberian Lindens Are Nested within Tilia cordata Mill. Diversity 2022, 14, 256. https://doi.org/10.3390/d14040256
Shekhovtsov SV, Shekhovtsova IN, Kosterin OE. Genotyping-by-Sequencing Analysis Shows That Siberian Lindens Are Nested within Tilia cordata Mill. Diversity. 2022; 14(4):256. https://doi.org/10.3390/d14040256
Chicago/Turabian StyleShekhovtsov, Sergei V., Irina N. Shekhovtsova, and Oleg E. Kosterin. 2022. "Genotyping-by-Sequencing Analysis Shows That Siberian Lindens Are Nested within Tilia cordata Mill" Diversity 14, no. 4: 256. https://doi.org/10.3390/d14040256