
Detection of the Endangered Stone Crayfish Austropotamobius torrentium (Schrank, 1803) and Its Congeneric A. pallipes in Its Last Italian Biotope by eDNA Analysis

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
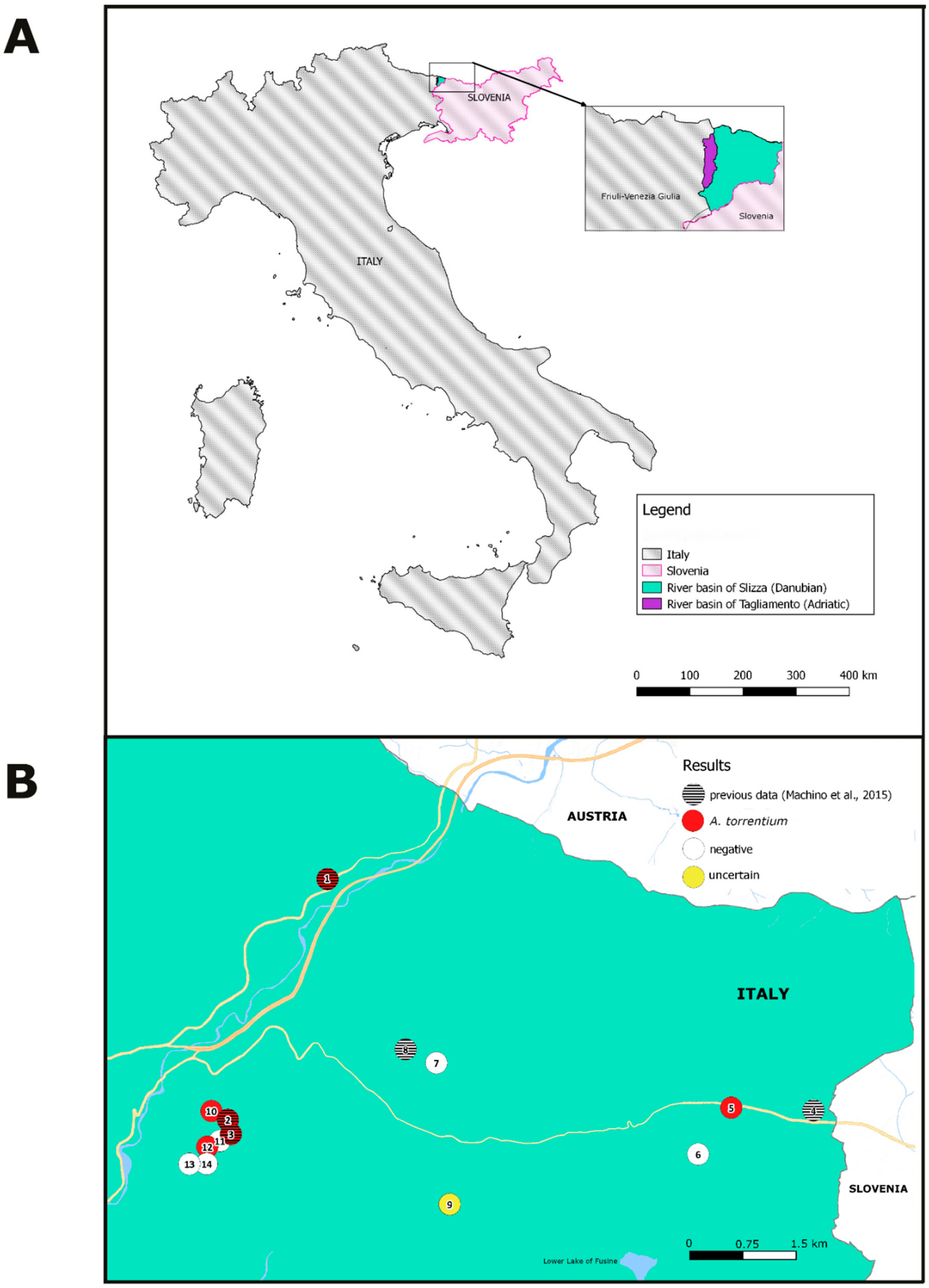
2.1. Study Design
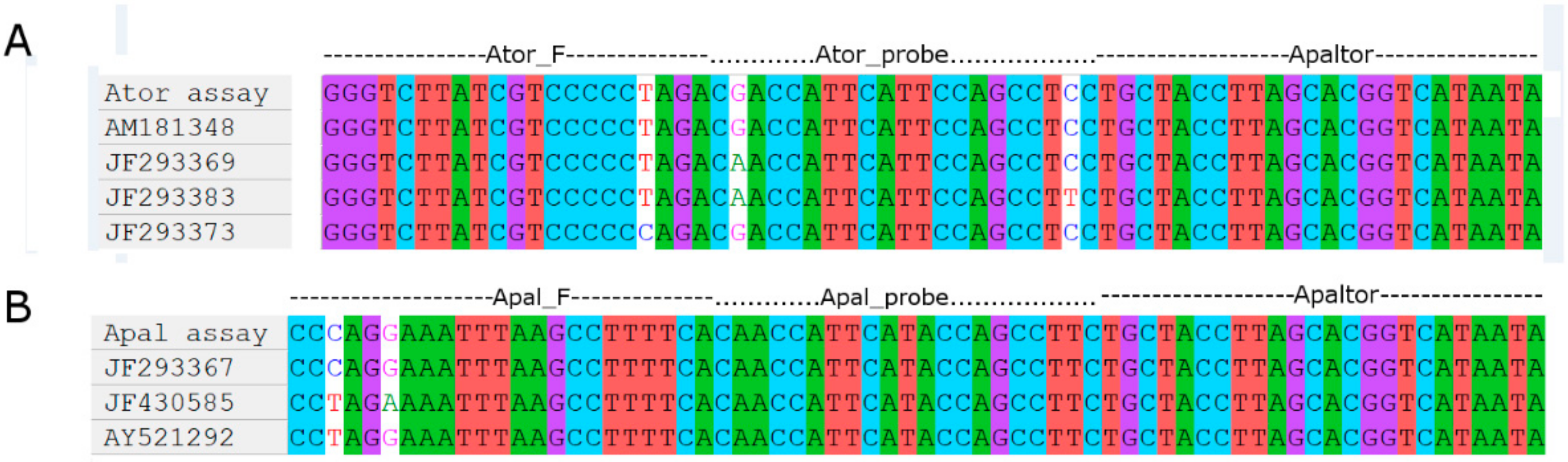
2.2. Primers Design and Species-Specificity Tests
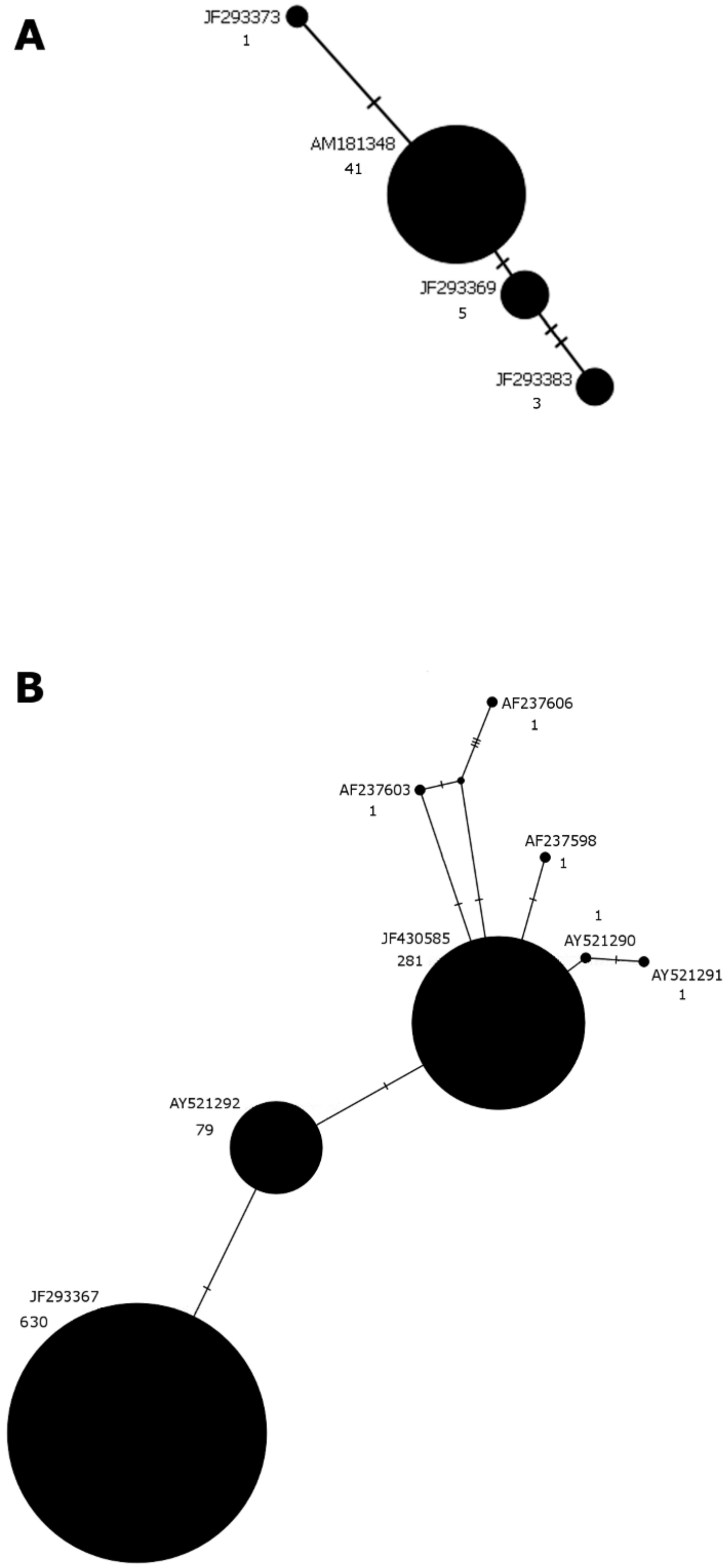
2.3. Animals Sampling and Genomics DNA Analysis
2.4. Environmental DNA Sampling and Analysis
3. Results
3.1. Apal and Ator Assays Species-Specificity
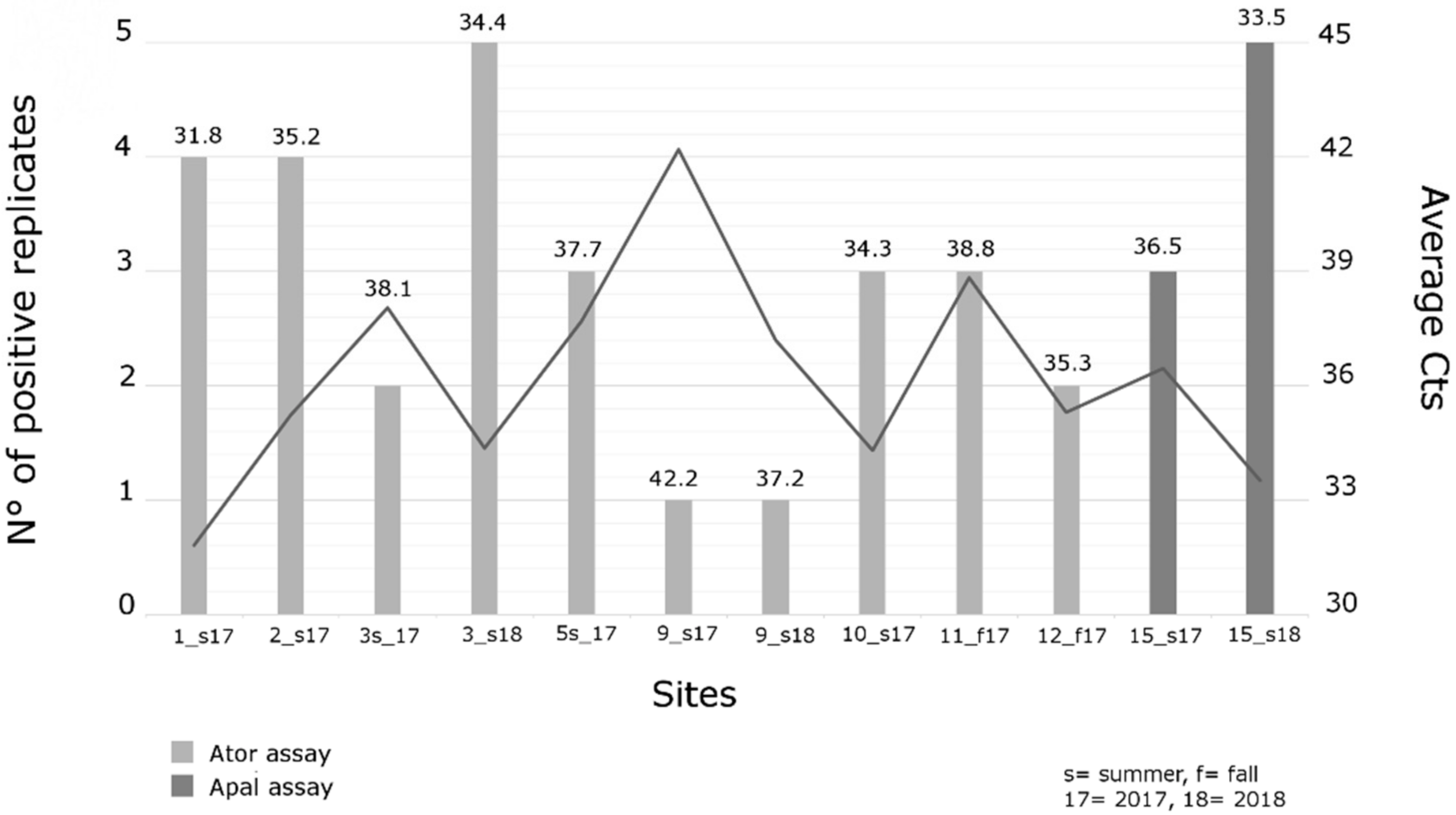
3.2. Results from Local Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, J.; Haffner, P. (Eds.) Publications Scientifiques du Muséum National d’Histoire Naturelle; Collection Patrimoines Naturels: Paris, France, 2006; pp. 49–130. [Google Scholar]
- Kouba, A.; Petrusek, A.; Kozák, P. Continental-wide distribution of crayfish species in Europe: Update and maps. Knowl. Manag. Aquat. Ecosyst. 2014, 413, 5. [Google Scholar] [CrossRef]
- Grandjean, F.; Souty-Grosset, C.; Bouchon, D. Systematics of the European Endangered Crayfish Species Austropotamobius Pallipes (Decapoda: Astacidae) with a Re-Examination of the Status of Austropotamobius Berndhauseri. J. Crustac. Biol. 2002, 22, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Pöckl, M.; Streissl, F. Austropotamobius Torrentium as an Indicator for Habitat Quality in Running Waters? Bull. Français Pêche Piscic. 2005, 376–377, 743–758. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Füreder, L. Linking species conservation management and legal species protection: A case study on stone crayfish. Freshw. Crayfish 2013, 19, 161–175. [Google Scholar]
- Füreder, L.; Edsman, L.; Holdich, D.; Kozak, P.; Machino, Y.; Pockl, M.; Renai, B.; Reynolds, J.; Schulz, R.W.; Schulz, H.K.; et al. Indigenous crayfish habitat and threats. In Atlas of Crayfish in Europe; Souty-Grosset, C., Holdich, D.M., Noel, P.Y., Reynolds, J.D., Haffner, P., Eds.; Museum National d’Histoire Naturelle: Paris, France, 2006; pp. 25–47. [Google Scholar]
- Maguire, I.; Erben, R.; Klobucar, G.; Lajtner, J. Year cycle of Austropotamobius torrentium (schrank) in streams on Medvednica Mountain (Croatia). Bull. Français Pêche Piscic. 2002, 367, 943–957. [Google Scholar] [CrossRef] [Green Version]
- Chucholl, C.; Schrimpf, A. The decline of endangered stone crayfish (Austropotamobius torrentium) in southern Germany is related to the spread of invasive alien species and land-use change. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 44–56. [Google Scholar] [CrossRef]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Cammà, C.; Ferri, N.; Zezza, D.; Marcacci, M.; Paolini, A.; Ricchiuti, L.; Lelli, R. Confirmation of crayfish plague in Italy: Detection of Aphanomyces astaci in white clawed crayfish. Dis. Aquat. Org. 2010, 89, 265–268. [Google Scholar] [CrossRef]
- Kušar, D.; Vrezec, A.; Ocepek, M.; Jencic, V. Aphanomyces astaci in wild crayfish populations in Slovenia: First report of persistent infection in a stone crayfish Austropotamobius torrentium population. Dis. Aquat. Org. 2013, 103, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Jussila, J.; Vrezec, A.; Jaklič, T.; Kukkonen, H.; Makkonen, J.; Kokko, H. Aphanomyces astaci isolate from latently infected stone crayfish (Austropotamobius torrentium) population is virulent. J. Invertebr. Pathol. 2017, 149, 15–20. [Google Scholar] [CrossRef]
- Füreder, L.; Gherardi, F.; Souty-Grosset, C. Austropotamobius torrentium (errata version published in 2017). 2010. Available online: https://www.iucnredlist.org/species/2431/121724677 (accessed on 13 September 2021).
- Vigneux, E.; Souty-Grosset, C.; Thibault, M.; Marnell, F. Roundtable Session 2A: National Legislation, EU Directives and Conservation. Bull. Français Pêche Piscic. 2002, 367, 887–898. [Google Scholar] [CrossRef] [Green Version]
- Füreder, L.; Gherardi, F.; Holdich, D.; Reynolds, J.; Sibley, P.; Souty-Grosset, C. Austropotamobius pallipes. IUCN Red List. Threat. Species 2010, 8235, e.T2430A9438817. [Google Scholar]
- Berger, C.; Štambuk, A.; Maguire, I.; Weiss, S.; Füreder, L. Integrating genetics and morphometrics in species conservation—A case study on the stone crayfish, Austropotamobius torrentium. Limnologica 2018, 69, 28–38. [Google Scholar] [CrossRef]
- Machino, Y.; Füreder, L. How to Find a Stone Crayfish Austropotamobius Torrentium (Schrank, 1803): A Biogeographic Study in Europe. Bull. Français Pêche Piscic. 2005, 376–377, 507–577. [Google Scholar] [CrossRef]
- de Luise, G. Recenti Acquisizioni Sul Comportamento e Sulla Distribuzione Nelle Acque Dolci Della Regione; Specchi, M., Ed.; Ente Tutela Pesca del Friuli Venezia Giulia: Udine, Italy, 2006. [Google Scholar]
- Machino, Y.; Tolazzi, L.; de Bortoli, M.; Pontarini, R.; Lapini, L. Austropotamobius torrentium (Schrank, 1803) in Italia (Crustacea: Decapoda, Astacidae; Italia Nord-Orientale). Gortania Biol. Zool. 2015, 37, 29–34. [Google Scholar]
- EIONET. Species Assessments at EU Biogeographical Level, Article 17 of the Habitats Directive. Available online: https://www.eionet.europa.eu/article17/species/summary/ (accessed on 18 January 2022).
- Thomsen, P.F.; Willerslev, E. Environmental DNA–An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Chadwick, D.D.A.; Pritchard, E.G.; Bradley, P.; Sayer, C.D.; Chadwick, M.A.; Eagle, L.J.B.; Axmacher, J.C. A novel ‘triple drawdown’ method highlights deficiencies in invasive alien crayfish survey and control techniques. J. Appl. Ecol. 2020, 58, 316–326. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef] [Green Version]
- Dejean, T.; Valentini, A.; Miquel, C.; Taberlet, P.; Bellemain, E.; Miaud, C. Improved detection of an alien invasive species through environmental DNA barcoding: The example of the American bullfrog Lithobates catesbeianus. J. Appl. Ecol. 2012, 49, 953–959. [Google Scholar] [CrossRef]
- Laramie, M.B.; Pilliod, D.S.; Goldberg, C.S. Characterizing the distribution of an endangered salmonid using environmental DNA analysis. Biol. Conserv. 2015, 183, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Doi, H.; Katano, I.; Sakata, Y.; Souma, R.; Kosuge, T.; Nagano, M.; Ikeda, K.; Yano, K.; Tojo, K. Detection of an endangered aquatic heteropteran using environmental DNA in a wetland ecosystem. R. Soc. Open Sci. 2017, 4, 170568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vences, M.; Lyra, M.L.; Perl, R.G.B.; Bletz, M.C.; Stanković, D.; Lopes, C.M.; Jarek, M.; Bhuju, S.; Geffers, R.; Haddad, C.F.B.; et al. Freshwater vertebrate metabarcoding on Illumina platforms using double-indexed primers of the mitochondrial 16S rRNA gene. Conserv. Genet. Resour. 2016, 8, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Bergman, P.S.; Schumer, G.; Blankenship, S.; Campbell, E. Detection of Adult Green Sturgeon Using Environmental DNA Analysis. PLoS ONE 2016, 11, e0153500. [Google Scholar] [CrossRef] [PubMed]
- Gorički, Š.; Stanković, D.; Snoj, A.; Kuntner, M.; Jeffery, W.R.; Trontelj, P.; Pavićević, M.; Grizelj, Z.; Năpăruş-Aljančič, M.; Aljančič, G. Environmental DNA in subterranean biology: Range extension and taxonomic implications for Proteus. Sci. Rep. 2017, 7, srep45054. [Google Scholar] [CrossRef] [PubMed]
- Troth, C.R.; Burian, A.; Mauvisseau, Q.; Bulling, M.; Nightingale, J.; Mauvisseau, C.; Sweet, M.J. Development and application of eDNA-based tools for the conservation of white-clawed crayfish. Sci. Total Environ. 2020, 748, 141394. [Google Scholar] [CrossRef]
- Atkinson, S.; Carlsson, J.E.L.; Ball, B.; Kelly-Quinn, M.; Carlsson, J. Field validation of an eDNA assay for the endangered white-clawed crayfish Austropotamobius pallipes. Freshw. Sci. 2019, 9, 503–509. [Google Scholar] [CrossRef]
- King, A.C.; Krieg, R.; Weston, A.; Zenker, A.K. Using eDNA to simultaneously detect the distribution of native and invasive crayfish within an entire country. J. Environ. Manag. 2022, 302, 113929. [Google Scholar] [CrossRef]
- Chucholl, F.; Fiolka, F.; Segelbacher, G.; Epp, L.S. eDNA Detection of Native and Invasive Crayfish Species Allows for Year-Round Monitoring and Large-Scale Screening of Lotic Systems. Front. Environ. Sci. 2021, 9, 639380. [Google Scholar] [CrossRef]
- Nukazawa, K.; Hamasuna, Y.; Suzuki, Y. Simulating the Advection and Degradation of the Environmental DNA of Common Carp along a River. Environ. Sci. Technol. 2018, 52, 10562–10570. [Google Scholar] [CrossRef]
- Sansom, B.J.; Sassoubre, L.M. Environmental DNA (eDNA) Shedding and Decay Rates to Model Freshwater Mussel eDNA Transport in a River. Environ. Sci. Technol. 2017, 51, 14244–14253. [Google Scholar] [CrossRef]
- Robson, H.L.A.; Noble, T.H.; Saunders, R.; Robson, S.; Burrows, D.W.; Jerry, D. Fine-tuning for the tropics: Application of eDNA technology for invasive fish detection in tropical freshwater ecosystems. Mol. Ecol. Resour. 2016, 16, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, C.; Stoll, S.; Strand, D.; Vrålstad, T.; Nowak, C.; Thines, M. eDNA-based crayfish plague monitoring is superior to conventional trap-based assessments in year-round detection probability. Hydrobiologia 2018, 807, 87–97. [Google Scholar] [CrossRef]
- Manfrin, C.; Souty-Grosset, C.; Anastácio, P.M.; Reynolds, J.; Giulianini, P.G. Detection and Control of Invasive Freshwater Crayfish: From Traditional to Innovative Methods. Diversity 2019, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Alderman, D.J.; Polglase, J.L. Disinfection for crayfish plague. Aquac. Res. 1985, 16, 203–205. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bertucci-Maresca, V. Genetic Characterization of Austropotamobius pallipes (Lereboullet, 1858) Complex in Friuli Venezia Giulia for Restocking Purposes. 2014. Available online: https://www.openstarts.units.it/bitstream/10077/10915/1/tesi_Bertucci Maresca.pdf (accessed on 14 April 2021).
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef]
- Peruzza, L.; Piazza, F.; Manfrin, C.; Bonzi, L.; Battistella, S.; Giulianini, P.G. Reproductive plasticity of a Procambarus clarkii population living 10 °C below its thermal optimum. Aquat. Invasions 2015, 10, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Govedič, M. Ugotavljanje prisotnosti tujerodnih vrst potočnih rakov na območju Krajinskega parka Ljubljansko barje Ugotavljanje prisotnosti tujerodnih vrst potočnih rakov na območju Krajinskega parka Ljubljansko barje. 2019. Available online: https://www.ljubljanskobarje.si/wp-content/uploads/2021/07/raki_barje_porocilo_2019_KONCNO.pdf (accessed on 8 July 2021).
- Govedič, M. First record of the spiny-cheek crayfish (Orconectes limosus) in Slovenia–300 km upstream from its known distribution in the Drava River. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 7. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2019, 2, 271–282. [Google Scholar] [CrossRef] [Green Version]
- van Doorn, R.; Klerks, M.M.; van Gent-Pelzer, M.P.E.; Speksnijder, A.G.C.L.; Kowalchuk, G.A.; Schoen, C.D. Accurate Quantification of Microorganisms in PCR-Inhibiting Environmental DNA Extracts by a Novel Internal Amplification Control Approach Using Biotrove OpenArrays. Appl. Environ. Microbiol. 2009, 75, 7253–7260. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, C.S.; Strickler, K.M.; Fremier, A.K. Degradation and dispersion limit environmental DNA detection of rare amphibians in wetlands: Increasing efficacy of sampling designs. Sci. Total Environ. 2018, 633, 695–703. [Google Scholar] [CrossRef]
- Lovrenčić, L.; Pavić, V.; Majnarić, S.; Abramović, L.; Jelić, M.; Maguire, I. Morphological diversity of the stone crayfish−Traditional and geometric morphometric approach. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 1. [Google Scholar] [CrossRef]
- Machino, Y. L’écrevisse de torrent Austropotamobius torrentium (Schrank, 1803) est bord de l’extinction en Italie. L’Astaciculteur Fr. 1996, 49, 9–12. [Google Scholar]
- Machino, Y.; Holdich, D.M. Distribution of Crayfish in Europe and Adjacent Countries: Updates and Comment. Freshw. Crayfish 2006, 15, 292–323. [Google Scholar]
- Daněk, T.; Musil, J.; Vlašánek, P.; Svobodová, J.; Johnsen, S.I.; Barteková, T.; Štrunc, D.; Barankiewicz, M.; Bouše, E.; Andersen, O. Telemetry of co-occurring noble crayfish (Astacus astacus) and stone crayfish (Austropotamobius torrentium): Diel changes in movement and local activity. Fundam. Appl. Limnol. 2018, 191, 339–352. [Google Scholar] [CrossRef]
- Takahashi, M.K.; Meyer, M.J.; McPhee, C.; Gaston, J.R.; Venesky, M.D.; Case, B.F. Seasonal and diel signature of eastern hellbender environmental DNA. J. Wildl. Manag. 2018, 82, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Buxton, A.S.; Groombridge, J.J.; Griffiths, R.A. Seasonal variation in environmental DNA detection in sediment and water samples. PLoS ONE 2018, 13, e0191737. [Google Scholar] [CrossRef] [PubMed]
- Furlan, E.M.; Gleeson, D.; Wisniewski, C.; Yick, J.; Duncan, R.P. eDNA surveys to detect species at very low densities: A case study of European carp eradication in Tasmania, Australia. J. Appl. Ecol. 2019, 56, 2505–2517. [Google Scholar] [CrossRef]
- Ikeda, K.; Doi, H.; Tanaka, K.; Kawai, T.; Negishi, J.N. Using environmental DNA to detect an endangered crayfish Cambaroides japonicus in streams. Conserv. Genet. Resour. 2016, 8, 231–234. [Google Scholar] [CrossRef]
- Harper, K.; Anucha, P.; Turnbull, J.; Bean, C.; Leaver, M. Searching for a signal: Environmental DNA (eDNA) for the detection of invasive signal crayfish, Pacifastacus leniusculus (Dana, 1852). Manag. Biol. Invasions 2018, 9, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, S.; Ushimaru, A.; Minamoto, T. A basin-scale application of environmental DNA assessment for rare endemic species and closely related exotic species in rivers: A case study of giant salamanders in Japan. J. Appl. Ecol. 2015, 52, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Troth, C.R.; Sweet, M.J.; Nightingale, J.; Burian, A. Seasonality, DNA degradation and spatial heterogeneity as drivers of eDNA detection dynamics. Sci. Total Environ. 2021, 768, 144466. [Google Scholar] [CrossRef]
- Buxton, A.S.; Groombridge, J.J.; Zakaria, N.; Griffiths, R.A. Seasonal variation in environmental DNA in relation to population size and environmental factors. Sci. Rep. 2017, 7, srep46294. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Klobučar, G.I.V.; Podnar, M.; Jelić, M.; Franjević, D.; Faller, M.; Štambuk, A.; Gottstein, S.; Simić, V.; Maguire, I. Role of the Dinaric Karst (western Balkans) in shaping the phylogeographic structure of the threatened crayfish Austropotamobius torrentium. Freshw. Biol. 2013, 58, 1089–1105. [Google Scholar] [CrossRef]
- Trontelj, P.; Machino, Y.; Sket, B. Phylogenetic and phylogeographic relationships in the crayfish genus Austropotamobius inferred from mitochondrial COI gene sequences. Mol. Phylogenetics Evol. 2005, 34, 212–226. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Species | Sequence ID | 5′-3′ Sequence | Amplicon Size (Base Pairs) |
|---|---|---|---|
| A. torrentium | Ator_F | GGGTCTTATCGTCCCCCTAGA | 152 bp |
| Ator_probe | [FAM]CGACCATTCATTCCAGCCTCC[BHQ1] | ||
| A. pallipes | Apal_F | CCCAGGAAATTTAAGCCTTTTCA | 136 bp |
| Apal_probe | [FAM]CAACCATTCATACCAGCCTTC[MGBEQ] | ||
| In common | Apaltor_R | TATTATGACCGTGCTAAGGTAGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfrin, C.; Zanetti, M.; Stanković, D.; Fattori, U.; Bertucci-Maresca, V.; Giulianini, P.G.; Pallavicini, A. Detection of the Endangered Stone Crayfish Austropotamobius torrentium (Schrank, 1803) and Its Congeneric A. pallipes in Its Last Italian Biotope by eDNA Analysis. Diversity 2022, 14, 205. https://doi.org/10.3390/d14030205
Manfrin C, Zanetti M, Stanković D, Fattori U, Bertucci-Maresca V, Giulianini PG, Pallavicini A. Detection of the Endangered Stone Crayfish Austropotamobius torrentium (Schrank, 1803) and Its Congeneric A. pallipes in Its Last Italian Biotope by eDNA Analysis. Diversity. 2022; 14(3):205. https://doi.org/10.3390/d14030205
Chicago/Turabian StyleManfrin, Chiara, Massimo Zanetti, David Stanković, Umberto Fattori, Victoria Bertucci-Maresca, Piero G. Giulianini, and Alberto Pallavicini. 2022. "Detection of the Endangered Stone Crayfish Austropotamobius torrentium (Schrank, 1803) and Its Congeneric A. pallipes in Its Last Italian Biotope by eDNA Analysis" Diversity 14, no. 3: 205. https://doi.org/10.3390/d14030205