
Spatial Variability in a Symbiont-Diverse Marine Host and the Use of Observational Data to Assess Ecological Interactions


Abstract
:1. Introduction
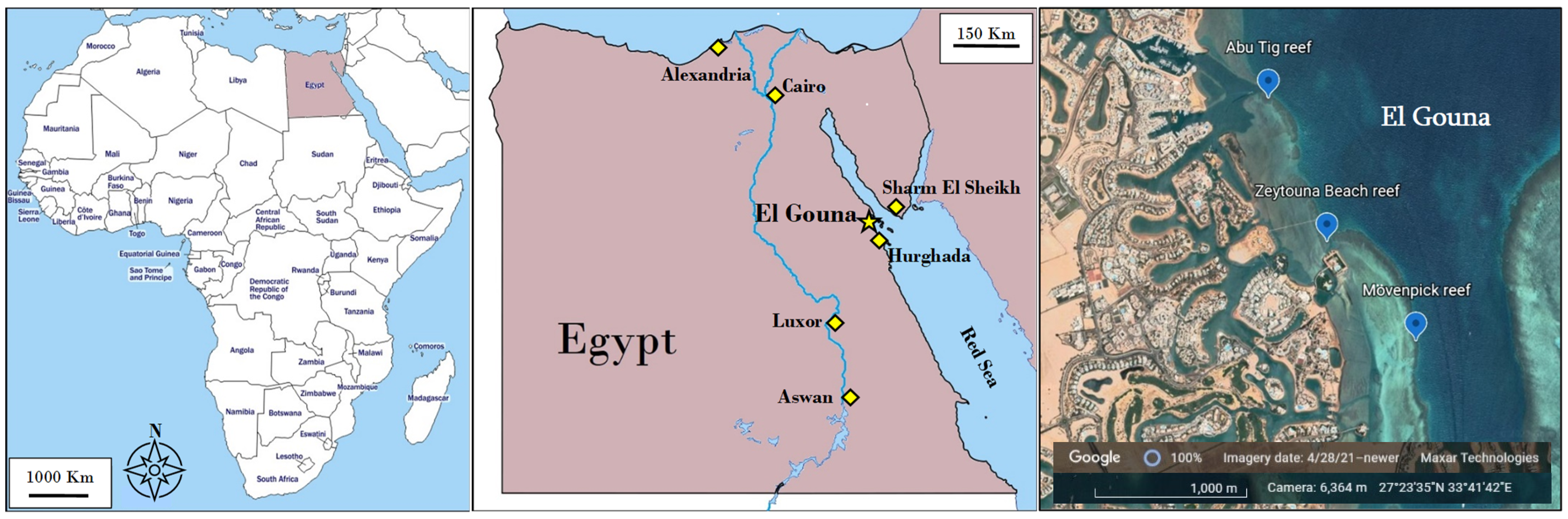
2. Materials and Methods
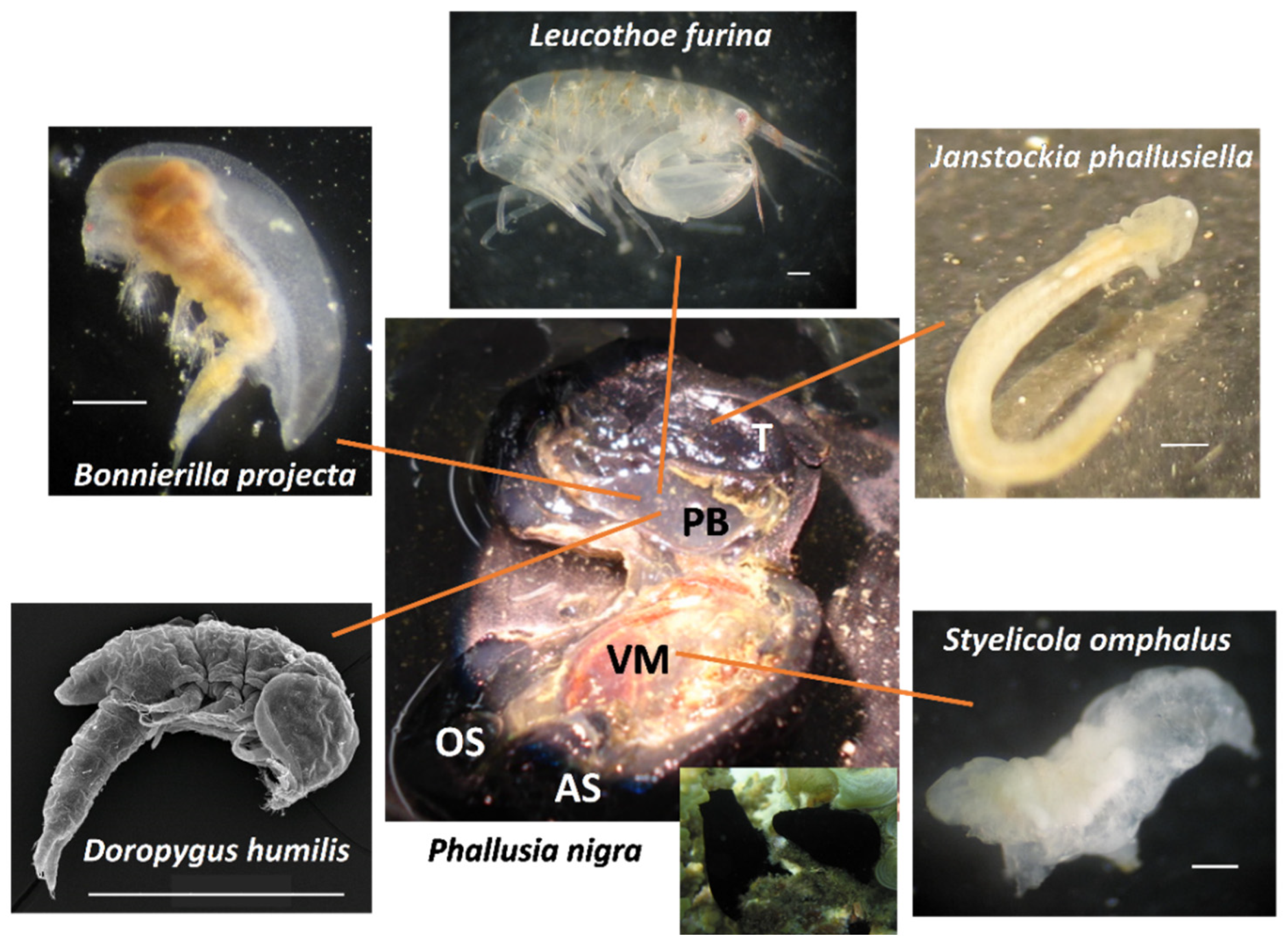
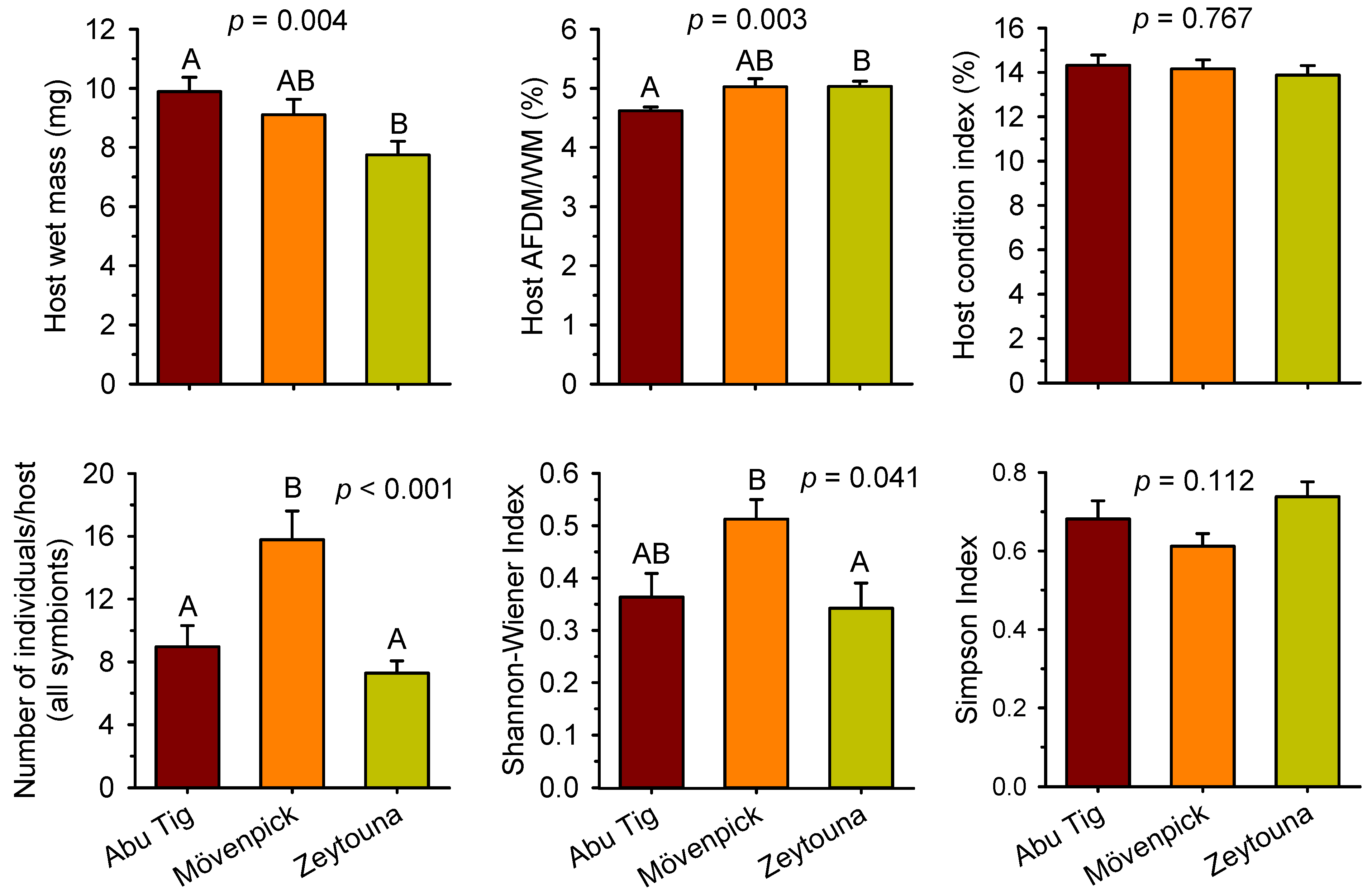
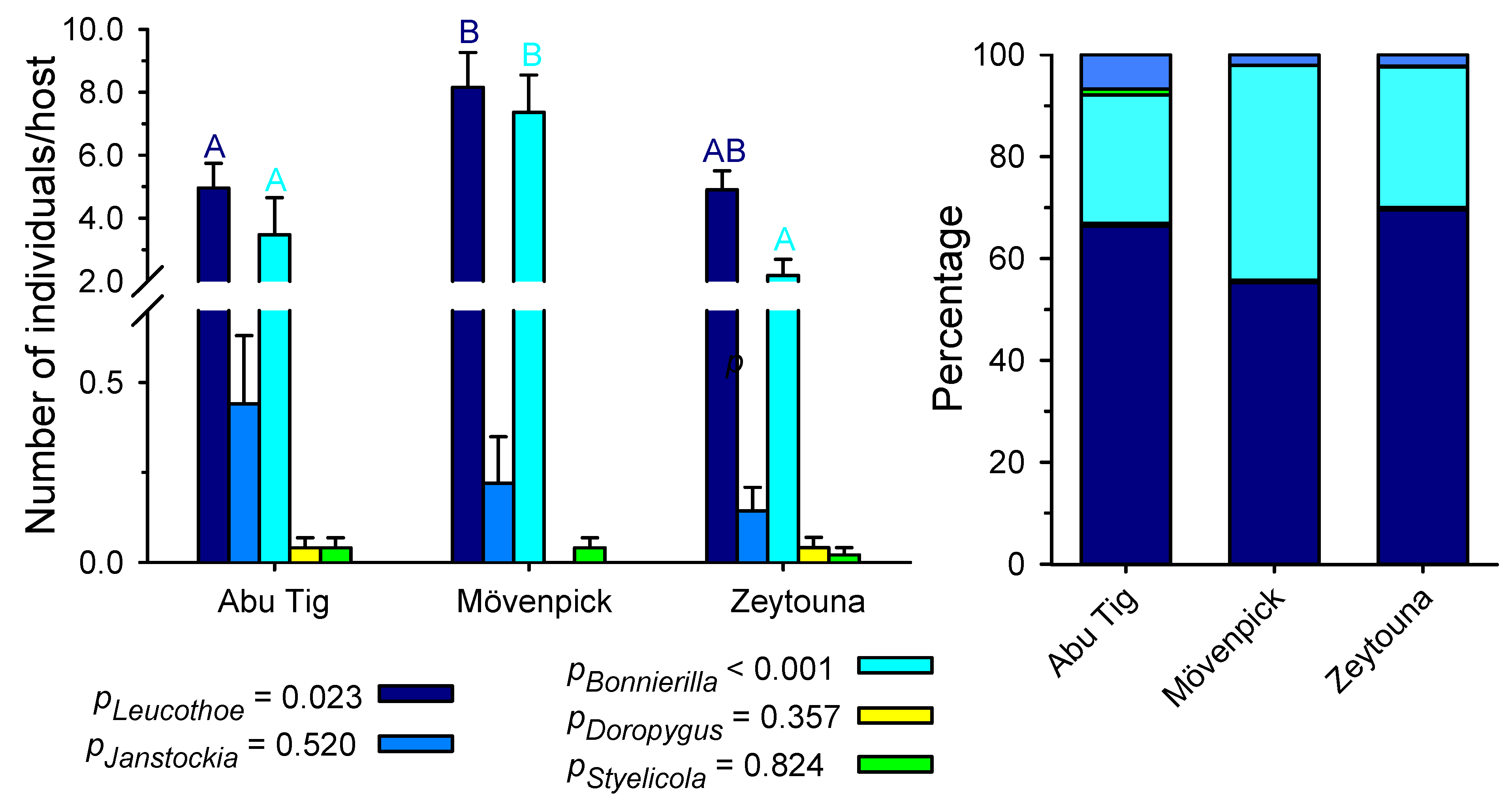
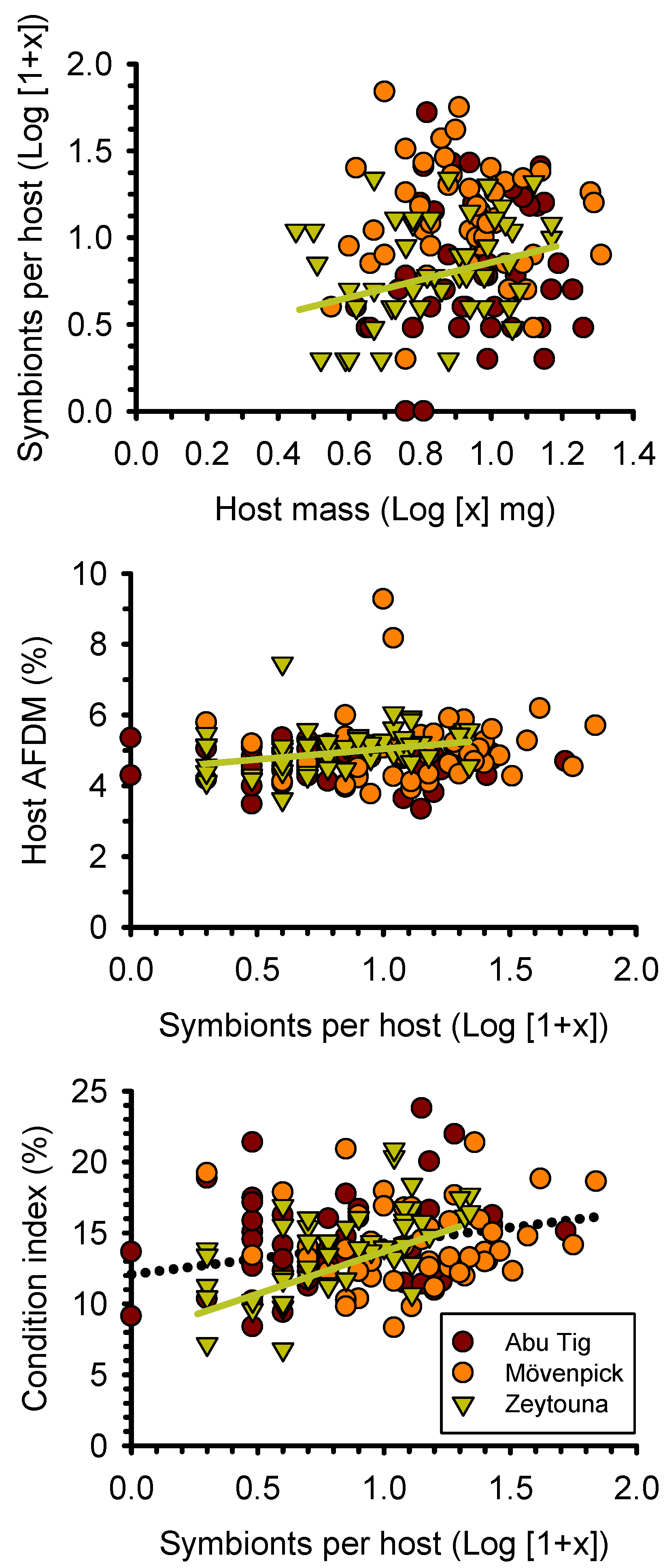
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Field Sites | Host Wet Mass (log x) | Host Total AFDM/WM | Host Tunic AFDM/WM | Host Body AFDM/WM | Host Condition Index | |
|---|---|---|---|---|---|---|
| Amphipods | 0.644 | 0.444 | 0.960 | 0.829 | 0.054 | |
| Copepods | 0.944 | 0.219 | 0.666 | 0.871 | 0.016 (0.039) | |
| All symbionts (log 1 + x) | 0.088 | 0.137 | 0.976 | 0.928 | 0.001 (0.068) | |
| Individual sites | ||||||
| Abu Tig | Amphipods Copepods All symbionts (log 1 + x) | 0.596 | 0.639 | 0.471 | 0.904 | 0.635 |
| 0.777 | 0.914 | 0.838 | 0.555 | 0.351 | ||
| 0.359 | 0.602 | 0.588 | 0.128 | 0.173 | ||
| Mövenpick | Amphipods Copepods All symbionts (log 1 + x) | 0.407 | 0.854 | 0.796 | 0.748 | 0.282 |
| 0.442 | 0.493 | 0.317 | 0.620 | 0.221 | ||
| 0.756 | 0.692 | 0.519 | 0.799 | 0.617 | ||
| Zeytouna Beach | Amphipods Copepods All symbionts (log 1 + x) | 0.113 | 0.260 | 0.976 | 0.989 | 0.020 (0.110) |
| 0.410 | 0.100 | 0.606 | 0.427 | <0.001 (0.212) | ||
| 0.027 (0.100) | 0.027 (0.100) | 0.653 | 0.780 | <0.001 (0.363) |
References
- Castro, P. Animal symbioses in coral reef communities: A review. Symbiosis 1988, 5, 161–184. [Google Scholar]
- Leung, T.L.F.; Poulin, R. Parasitism, commensalism, and mutualism: Exploring the many shades of symbioses. Vie Milieu 2008, 58, 107–115. [Google Scholar]
- Glynn, P.W.; Enochs, I.C. Invertebrates and their roles in coral reef ecosystems. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 273–325. ISBN 978-94-007-0113-7. [Google Scholar]
- Hirsch, A.M. Plant-microbe symbioses: A continuum from commensalism to parasitism. Symbiosis 2004, 37, 345–363. [Google Scholar]
- Cheney, K.L.; Côté, I.M. Mutualism or parasitism? The variable outcome of cleaning symbioses. Biol. Lett. 2005, 1, 162–165. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.L.; Creed, R.P.; Skelton, J.; Rollins, M.A.; Farrell, K.J. The fine line between mutualism and parasitism: Complex effects in a cleaning symbiosis demonstrated by multiple field experiments. Oecologia 2012, 170, 199–207. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Bruna, E.M. Pursuing the big questions about interspecific mutualism: A review of theoretical approaches. Oecologia 2000, 125, 321–330. [Google Scholar] [CrossRef]
- Silknetter, S.; Kanno, Y.; Kanapeckas Métris, K.L.; Cushman, E.; Darden, T.L.; Peoples, B.K. Mutualism or parasitism: Partner abundance affects host fitness in a fish reproductive interaction. Freshw. Biol. 2019, 64, 175–182. [Google Scholar] [CrossRef]
- Saffo, M.B.; McCoy, A.M.; Rieken, C.; Slamovits, C.H. Nephromyces, a beneficial apicomplexan symbiont in marine animals. Proc. Natl. Acad. Sci. USA 2010, 107, 16190–16195. [Google Scholar] [CrossRef] [Green Version]
- Cumbo, V.R.; Baird, A.H.; Moore, R.B.; Negri, A.P.; Neilan, B.A.; Salih, A.; van Oppen, M.J.H.; Wang, Y.; Marquis, C.P. Chromera velia is endosymbiotic in larvae of the reef corals Acropora digitifera and A. tenuis. Protist 2013, 164, 237–244. [Google Scholar] [CrossRef]
- Kwong, W.K.; del Campo, J.; Mathur, V.; Vermeij, M.J.A.; Keeling, P.J. A widespread coral-infecting apicomplexan with chlorophyll biosynthesis genes. Nature 2019, 568, 103–107. [Google Scholar] [CrossRef]
- Halliday-Isaac, A.K.; Robinson, J.B.; Cruz-Rivera, E.; Campbell, A.G.; Sikkel, P.C. Environmental correlates of prevalence of an intraerythrocytic apicomplexan infecting Caribbean damselfish. Parasitologia 2021, 1, 69–82. [Google Scholar] [CrossRef]
- Carlton, J.T.; Blakeslee, A.M.H.; Fowler, A.E. Accidental associates are not symbionts: The absence of a non-parasitic endosymbiotic community inside the common periwinkle Littorina littorea (Mollusca: Gastropoda). Mar. Biol. 2020, 167, 97. [Google Scholar] [CrossRef]
- Hirose, E.; Oka, A.T.; Akahori, M. Sexual reproduction of the photosymbiotic ascidian Diplosoma virens in the Ryukyu Archipelago, Japan: Vertical transmission, seasonal change, and possible impact of parasitic copepods. Mar. Biol. 2005, 146, 677–682. [Google Scholar] [CrossRef]
- Parmentier, E.; Michel, L. Boundary lines in symbiosis forms. Symbiosis 2013, 60, 1–5. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Interspecific competition of symbiotic and fouling species of red king crab in the Barents Sea. Doklady Biol. Sci. 2011, 440, 300–302. [Google Scholar] [CrossRef]
- Devlaming, V.; Grossman, G.; Chapman, F. On the use of the gonosomatic index. Comp. Biochem. Physiol. A 1982, 73, 31–39. [Google Scholar] [CrossRef]
- Lucas, A.; Beninger, P.G. The use of physiological condition indices in marine bivalve aquaculture. Aquaculture 1985, 44, 187–200. [Google Scholar] [CrossRef]
- Crosby, M.P.; Gale, L.D. A review and evaluation of bivalve condition index methodologies with a suggested standard method. J. Shellfish Res. 1990, 9, 233–237. [Google Scholar]
- West, G. Methods of assessing ovarian development in fishes: A review. Mar. Freshw. Res. 1990, 41, 199–222. [Google Scholar] [CrossRef]
- Fernandez, C.; Boudouresque, C.-F. Phenotypic plasticity of Paracentrotus lividus (Echinodermata: Echinoidea) in a lagoonal environment. Mar. Ecol. Prog. Ser. 1997, 152, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, C.; Boudouresque, C.-F. Nutrition of the sea urchin Paracentrotus lividus (Echinodermata: Echinoidea) fed different artificial food. Mar. Ecol. Prog. Ser. 2000, 204, 131–141. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Brown-Peterson, N.J.; Murua, H.; Tomkiewicz, J.; Wyanski, D.M.; Saborido-Rey, F. Emerging issues and methodological advances in fisheries reproductive biology. Mar. Coast. Fish. 2011, 3, 32–51. [Google Scholar] [CrossRef] [Green Version]
- Labocha, M.K.; Hayes, J.P. Morphometric indices of body condition in birds: A review. J. Ornithol. 2012, 153, 1–22. [Google Scholar] [CrossRef]
- Ouréns, R.; Freire, J.; Fernández, L. Definition of a new unbiased gonad index for aquatic invertebrates and fish: Its application to the sea urchin Paracentrotus lividus. Aquat. Biol. 2012, 17, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Unglaub, B.; Steinfartz, S.; Kühne, D.; Haas, A.; Schmidt, B.R. The relationships between habitat suitability, population size and body condition in a pond-breeding amphibian. Basic Appl. Res. 2018, 27, 20–29. [Google Scholar] [CrossRef]
- Dunphy, B.J.; Wells, R.M.G. Endobiont infestation, shell strength and condition index in wild populations of New Zealand abalone, Haliotis iris. Mar. Freshwater Res. 2001, 52, 781–786. [Google Scholar] [CrossRef]
- O’Connell-Milne, S.A.; Poulin, R.; Savage, C.; Rayment, W. Reduced growth, body condition and foot length of the bivalve Austrovenus stutchburyi in response to parasite infection. J. Exp. Mar. Biol. Ecol. 2016, 474, 23–28. [Google Scholar] [CrossRef]
- Bazterrica, M.C.; Bruschetti, C.M.; Alvarez, M.F.; Iribarne, O.; Botto, F. Effects of macroalgae on the recruitment, growth, and body condition of an invasive reef forming polychaete in a south-western Atlantic coastal lagoon. J. Sea Res. 2014, 88, 121–129. [Google Scholar] [CrossRef]
- Shenkar, N. Ascidian (Chordata, Ascidiacea) diversity in the Red Sea. Mar. Biodiv. 2012, 42, 459–469. [Google Scholar] [CrossRef]
- Vandepas, L.E.; Oliveira, L.M.; Lee, S.S.C.; Hirose, E.; Rocha, R.M.; Swalla, B.J. Biogeography of Phallusia nigra: Is it really black and white? Biol. Bull. 2015, 228, 52–64. [Google Scholar] [CrossRef]
- Rocha, R.M.D.; Lotufo, T.M.D.C.; de Almeida Rodrigues, S. The biology of Phallusia nigra Savigny, 1816 (Tunicata: Ascidiacea) in Southern Brazil: Spatial distribution and reproductive cycle. Bull. Mar. Sci. 1999, 64, 77–88. [Google Scholar]
- Kondilatos, G.; Corsini-Foka, M.; Pancucci-Papadopoulou, M.-A. Occurrence of the first non-indigenous ascidian Phallusia nigra Savigny, 1816 (Tunicata: Ascidiacea) in Greek waters. Aquat. Inv. 2010, 5, 181–184. [Google Scholar] [CrossRef]
- Naser, H.A. Variability of marine macrofouling assemblages in a marina and a mariculture centre in Bahrain, Arabian Gulf. Reg. Stud. Mar. Sci. 2017, 16, 162–170. [Google Scholar] [CrossRef]
- Goodbody, I. The biology of Ascidia nigra (Savigny). I. Survival and mortality in an adult population. Biol. Bull. 1962, 122, 40–51. [Google Scholar] [CrossRef]
- Goodbody, I. The biology of Ascidia nigra (Savigny). III. The annual pattern of colonization. Biol. Bull. 1965, 129, 128–133. [Google Scholar] [CrossRef]
- Goodbody, I.; Gibson, J. The biology of Ascidia nigra (Savigny) V. Survival in populations settled at different times of the year. Biol. Bull. 1974, 146, 217–237. [Google Scholar] [CrossRef]
- Al-Sofyani, A.M.A.; Satheesh, S. Recruitment patterns of the solitary ascidian Phallusia nigra Savigny, 1816 on artificial substrates submerged in the central Red Sea, Saudi Arabia. Oceanol. Hydrobiol. Stud. 2019, 48, 262–269. [Google Scholar] [CrossRef]
- Granot, I.; Shenkar, N.; Belmaker, J. Habitat niche breadth predicts invasiveness in solitary ascidians. Ecol. Evol. 2017, 7, 7838–7847. [Google Scholar] [CrossRef]
- Ghazilou, A.; Koochaknejad, E.; Ershadifar, H.; Negarestan, H.; Kor, K.; Baskaleh, G. Infestation biology of Phallusia nigra (Tunicata, Phlebobranchia) on hard corals in a subtropical bay. Mar. Ecol. Prog. Ser. 2019, 626, 135–143. [Google Scholar] [CrossRef]
- Hirose, E.; Yamashiro, H.; Mori, Y. Properties of tunic acid in the ascidian Phallusia nigra (Ascidiidae, Phlebobranchia). Zool. Sci. 2001, 18, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Odate, S.; Pawlik, J.R. The Role of vanadium in the chemical defense of the solitary tunicate, Phallusia nigra. J. Chem. Ecol. 2007, 33, 643–654. [Google Scholar] [CrossRef]
- Mayzel, B.; Haber, M.; Ilan, M. Chemical defense against fouling in the solitary ascidian Phallusia nigra. Biol. Bull. 2014, 227, 232–241. [Google Scholar] [CrossRef]
- Stock, J.H. Report on the Notodelphyidae (Copepoda, Cyclopoida) of the Israel South Red Sea Expedition. Bull. Sea Fish. Res. Stat. Israel 1967, 27, 1–126. [Google Scholar]
- White, K.N. A Taxonomic review of the Leucothoidae (Crustacea: Amphipoda). Zootaxa 2011, 3078, 1–113. [Google Scholar] [CrossRef]
- Kim, I.-H.; Cruz-Rivera, E.; Sherif, M.-E.-D.; El-Sahhar, S. Cyclopoid copepods (Ascidicolidae, Notodelphyidae) associated with Phallusia nigra Savigny, 1816 (Ascidiacea) in the Red Sea: A new ascidicolid and first descriptions of the males from two notodelphyids. J. Crust. Biol. 2016, 36, 553–566. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Nygren, A.; Cruz-Rivera, E. Proceraea exoryxae sp. nov. (Annelida, Syllidae, Autolytinae), the first known polychaete miner tunneling into the tunic of an ascidian. PeerJ 2017, 5, e3374. [Google Scholar] [CrossRef] [Green Version]
- White, K.N.; Krapp-Schickel, T. Red Sea Leucothoidae (Crustacea: Amphipoda) including new and re-described species. Eur. J. Taxon. 2017, 324, 1–40. [Google Scholar] [CrossRef] [Green Version]
- White, K.N. Caribbean Leucothoidae (Crustacea: Amphipoda) of Panama. Gulf Caribb. Res. 2011, 23, 23–35. [Google Scholar] [CrossRef]
- Senna, A.R.; Andrade, L.F.; Ramos, B.S.; Skinner, L.F. A new ascidian-dwelling species of Leucothoe Leach, 1814 (Amphipoda: Leucothoidae) from Ilha Grande Bay, Rio de Janeiro State, Brazil. J. Nat. Hist. 2021, 55, 1441–1460. [Google Scholar] [CrossRef]
- Hernández, J.E.; Bolaños, J.A.; Palazón, J.L.; Hernández, G.; Lira, C.; Baeza, J.A. The enigmatic life history of the symbiotic crab Tunicotheres moseri (Crustacea, Brachyura, Pinnotheridae): Implications for its mating system and population structure. Biol. Bull. 2012, 223, 278–290. [Google Scholar] [CrossRef]
- Thomas, J.D.; Klebba, K.N. New species and host associations of commensal leucothoid amphipods from coral reefs in Florida and Belize (Crustacea:Amphipoda). Zootaxa 2007, 1494, 1–44. [Google Scholar] [CrossRef]
- White, K.; Reimer, J. Commensal Leucothoidae (Crustacea, Amphipoda) of the Ryukyu Archipelago, Japan. Part I: Ascidian-dwellers. ZooKeys 2012, 163, 13–55. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.-S. Origin and evolution of the parasitic cyclopoid copepods. Int. J. Parasitol. 1994, 24, 1293–1300. [Google Scholar] [CrossRef]
- Kim, I.-H.; Boxshall, G.A. Untold diversity: The astonishing species richness of the Notodelphyidae (Copepoda: Cyclopoida), a family of symbiotic copepods associated with ascidians (Tunicata). Megataxa 2020, 4, 1–660. [Google Scholar] [CrossRef]
- Kim, I.-H.; Boxshall, G.A. Copepods (Cyclopoida) associated with ascidian hosts: Ascidicolidae, Buproridae, Botryllophilidae, and Enteropsidae, with descriptions of 84 new species. Zootaxa 2021, 4978, 1–286. [Google Scholar] [CrossRef]
- Ortiz, M. Claves ilustradas para la clasificación de los anfípodos (Crustacea, Peracarida) de Cuba: Morfología y taxonomía. Rev. Investig. Mar. 2021, 41, 1–108. [Google Scholar]
- Thiel, M. Host-use and population demographics of the ascidian-dwelling amphipod Leucothoe spinicarpa: Indication for extended parental care and advanced social behaviour. J. Nat. Hist. 1999, 33, 193–206. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Marchenkov, A. A new genus of notodelphyid copepod (Crustacea, Copepoda, Cyclopoida) from a compound ascidian host collected in the Suez Canal. Zoosystema 2005, 27, 483–497. [Google Scholar]
- Schellenberg, A. Neuo Notodelphyiden des Berliner und Hamburger Museums mit einer Übersicht der ascidien bowohnenden Gattungen und Arten. Mitt. Zool. Mus. 1922, 10, 219–274. [Google Scholar]
- Gurney, R. Report on the Crustacea:—Copepoda (littoral and semi-parasitic). Trans. Zool. Soc. 1927, 22, 451–577. [Google Scholar] [CrossRef]
- El Sherbiny Ahmed, H.; Sherif Ahmed, H.; Hassan Ali, N. Model for environmental risk assessment of tourism project construction on the Egyptian Red Sea Coast. J. Environ. Eng. 2006, 132, 1272–1281. [Google Scholar] [CrossRef]
- Vanderstraete, T.; Goossens, R.; Ghabour, T.K. The use of multi-temporal Landsat Images for the change detection of the coastal zone near Hurghada, Egypt. Int. J. Remote Sens. 2006, 27, 3645–3655. [Google Scholar] [CrossRef]
- Marshall, N.A.; Marshall, P.A.; Abdulla, A.; Rouphael, T.; Ali, A. preparing for climate change: Recognising its early impacts through the perceptions of dive tourists and dive operators in the Egyptian Red Sea. Curr. Issues Tour. 2011, 14, 507–518. [Google Scholar] [CrossRef]
- Nassar, K.; El-Adawy, A.; Zakaria, M.; Diab, R.; Masria, A. Quantitative appraisal of naturalistic/anthropic shoreline shifts for Hurghada: Egypt. Mar. Georesour. Geotechnol. 2021. [Google Scholar] [CrossRef]
- Moufaddal, W.M. Use of Satellite imagery as environmental impact assessment tool: A case study from the NW Egyptian Red Sea coastal zone. Environ. Monit. Assess. 2005, 107, 427–452. [Google Scholar] [CrossRef]
- Ballarin, L.; Burighel, P. Tunicata and Cephalochordata. In Biological Science Fundamentals and Systematics; Encyclopedia of Biological, Physiological and Health Sciences; Minelli, A., Contrafatto, G., Eds.; EOLSS Publishers, UNESCO: Oxford, UK, 2009; Volume IV, pp. 43–67. ISBN 978-84826-189-1. [Google Scholar]
- Cruz-Rivera, E.; Hay, M.E. Macroalgal traits and the feeding and fitness of an herbivorous amphipod: The roles of selectivity, mixing, and compensation. Mar. Ecol. Prog. Ser. 2001, 218, 249–266. [Google Scholar] [CrossRef]
- Prado, P.; Heck, K.L. Seagrass selection by omnivorous and herbivorous consumers: Determining factors. Mar. Ecol. Prog. Ser. 2011, 429, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Rivera, E.; Friedlander, M. Effects of algal phenotype on mesograzer feeding. Mar. Ecol. Prog. Ser. 2013, 490, 69–78. [Google Scholar] [CrossRef]
- Okumuş, İ.; Stirling, H.P. Seasonal variations in the meat weight, condition index and biochemical composition of mussels (Mytilus edulis, L.) in suspended culture in two Scottish Sea lochs. Aquaculture 1998, 159, 249–261. [Google Scholar] [CrossRef]
- Adjei-Boateng, D.; Wilson, J.G. Body condition and gametogenic cycle of Galatea paradoxa (Mollusca: Bivalvia) in the Volta River Estuary, Ghana. Estuar. Coast. Shelf Sci. 2013, 132, 94–98. [Google Scholar] [CrossRef]
- Rainier, J.S.; Mann, R.L. A comparison of methods for calculating condition index in eastern oyster, Crassostrea virginica (Gmelin, 1791). J. Shellfish Res. 1992, 11, 55–58. [Google Scholar]
- Zeng, Y.; Yang, H. Review of molluscan bivalve condition index calculations and application in northern quahogs Mercenaria mercenaria. Aquac. Res. 2021, 52, 23–36. [Google Scholar] [CrossRef]
- Gage, J. Seasonal Cycles of Notodelphys and Ascidicola, copepod associates with Ascidiella (Ascidiacea). J. Zool. 1966, 150, 223–233. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Tierno de Figueroa, J.M.; Navarro-Barranco, C.; Ros, M.; Sánchez-Moyano, J.E.; Moreira, J. Dietary analysis of the marine Amphipoda (Crustacea: Peracarida) from the Iberian Peninsula. J. Sea Res. 2014, 85, 508–517. [Google Scholar] [CrossRef]
- da Silva Ramos, E.K.; Batista Rosa, A.H.; Cobo, V.J. Influence of the endo-symbiont Leucothoe wuriti (Thomas & Klebba, 2007) (Crustacea, Leucothoidae) on the biomass of Phallusia nigra (Savigny, 1816) (Tunicata, Ascididae), in the northeastern coast of the São Paulo State, Brazil. Rev. Biociên. 2015, 21, 38–43. [Google Scholar]
- Egan, E.A. The seasonal occurrence of the copepod Pachypygus australis Gotto (Notodelphyidae) in its host Pyura pachydermatina (Herdman) Pyuridae: Ascidiacea. J. Exp. Mar. Biol. Ecol. 1984, 76, 247–262. [Google Scholar] [CrossRef]
- Svavarsson, J. Life Cycle and population dynamics of the symbiotic copepod Lichomolgus canui Sars associated with the ascidian Halocynthia pyriformis (Rathke). J. Exp. Mar. Biol. Ecol. 1990, 142, 1–12. [Google Scholar] [CrossRef]
- Saito, S. Density and adult ratio of the symbiotic harpacticoid copepod Idomene purpurocincta in the compound ascidian host Aplidium yamazii. Plankton Benthos Res. 2009, 4, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Jakob, E.M.; Marshall, S.D.; Uetz, G.W. Estimating fitness: A comparison of body condition indices. Oikos 1996, 77, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Peig, J.; Green, A.J. The paradigm of body condition: A critical reappraisal of current methods based on mass and length. Funct. Ecol. 2010, 24, 1323–1332. [Google Scholar] [CrossRef]
- Labocha, M.K.; Schutz, H.; Hayes, J.P. Which body condition index is best? Oikos 2014, 123, 111–119. [Google Scholar] [CrossRef]
- Addicott, J.F. A multispecies aphid–ant association: Density dependence and species-specific effects. Can. J. Zool. 1979, 57, 558–569. [Google Scholar] [CrossRef]
- Drew, G.; King, K. More or less? The effect of symbiont density in protective mutualisms. Am. Nat. 2022. [Google Scholar] [CrossRef]
- Illg, P.L. North American copepods of the family Notodelphyidae. Proc. US Natl. Mus. 1958, 107, 463–649. [Google Scholar] [CrossRef] [Green Version]
- Dudley, P.L. A light and electron microscopic study of tissue interactions between a parasitic copepod, Scolecodes huntsmani (Henderson), and its host ascidian, Styela gibbsii (Stimpson). J. Morphol. 1968, 124, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Conradi, M.; López-González, P.J. A new family of cyclopoid copepods (Fratiidae) symbiotic in the ascidian (Clavelina dellavallei) from Cádiz, Spain. J. Zool. 1998, 246, 39–48. [Google Scholar]
- Ho, J.; Kim, I.-H. Two species of copepoda parasitic in the algal-bearing ascidian, Didemnum molle (Herdman), in Okinawa, Japan. Proc. Biol. Soc. Wash. 2009, 122, 414–425. [Google Scholar] [CrossRef]
- Marchenkov, A.; Boxshall, G.A. A new notodelphyid copepod, Paranotodelphys illgi n. sp. (Copepoda: Cyclopoida), parasitic in the ascidian Corynascidia herdmani Ritter in the North Pacific. Syst. Parasitol. 2003, 54, 43–52. [Google Scholar] [CrossRef]
- Stock, J.H. Parasite or commensal? Notodelphys weberi, a new South African ascidicole copepod. Amsterdam Nat. 1950, 1, 36–42. [Google Scholar]
- Illg, P.L. Occurrence in Sagami Bay, Japan, of Scolecodes, a remarkable copepod parasite of ascidians. Publ. Seto Mar. Biol. Lab. 1970, 18, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.D.; Taylor, G.W. Mouthpart morphology and feeding strategies of the commensal amphipod, Anamixis hanseni Stebbing. Bull. Mar. Sci. 1981, 31, 462–467. [Google Scholar]
- Fleming, T.H.; Holland, J.N. The evolution of obligate pollination mutualisms: Senita cactus and senita moth. Oecologia 1998, 114, 368–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldroyd, G.E.D.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Mortier, V.; Holsters, M.; Goormachtig, S. Never too many? How legumes control nodule numbers. Plant Cell Environ. 2012, 35, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.O.; Pitzschke, A. Plants make galls to accommodate foreigners: Some are friends, most are foes. New Phytol. 2020, 225, 1852–1872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yan, C.; Zhang, H. Mutualism between antagonists: Its ecological and evolutionary implications. Integr. Zool. 2021, 16, 84–96. [Google Scholar] [CrossRef]
- Alfaya, J.E.F.; Galván, D.E.; Machordom, A.; Penchaszadeh, P.E.; Bigatti, G. Malacobdella arrokeana: Parasite or commensal of the giant clam Panopea abbreviata? Zool. Sci. 2015, 32, 523–530. [Google Scholar] [CrossRef] [Green Version]





| Symbiont | Geographic Location | References |
|---|---|---|
| Crustacea Amphipoda | ||
| Amphilochus ascidicola Ortiz and Atienza, 2001 | Caribbean (Venezuela) | [57] |
| Leucothoe angraensis Senna, Andrade, Ramos & Skinner, 2021 | South Atlantic (Brazil) | [50] |
| L. flammosa Thomas and Klebba 2007 | Caribbean (Cuba) | [57] |
| L. furina (Savigny, 1816) | Red Sea (Egypt) | [46] |
| L. spinicarpa (Abildgaard, 1789) | North Atlantic (USA) | [58] |
| L. wuriti Thomas and Klebba 2007 | North Atlantic (USA), Caribbean (Belize, Panama) | [49,52] |
| Brachiura | ||
| Tunicotheres moseri (Rathbun, 1918) | Caribbean (Jamaica, Venezuela) | [35,51] |
| Copepoda | ||
| Bonnierilla projecta Stock, 1967 | Red Sea (Egypt, Erithrea) | [44,46] |
| Doropygus humilis1 Stock, 1967 | Red Sea (Egypt, Erithrea) | [44,46] |
| Janhius brevis2 (Stock, 1967) | Red Sea (Erithrea) | [44] |
| Janstockia phallusiella Boxshall & Marchenkov, 2005 | Red Sea (Egypt) | [46,59] |
| Lonchidiopsis tripes Stock, 1967 | Red Sea (Erithrea) | [44] |
| Notodelphys ciliata Schellenberg, 1922 | Red Sea (Egypt) | [60] |
| Notodelphys steinitzi Stock, 1967 | Red Sea (Erithrea) | [44] |
| Paranotodelphys phallusiae (Gurney, 1927) | Red Sea (Egypt) | [61] |
| Styelicola omphalus Kim I.H., Cruz-Rivera, Sherif & El-Sahhar, 2016 | Red Sea (Egypt) | [46] |
| Annelida Polychaeta | ||
| Proceraea exoryxae Martin, Nygren & Cruz-Rivera, 2017 | Red Sea (Egypt) | [47] |
| All Field Sites | Bonnierilla | Doropygus | Janstockia | Styelicola | |
|---|---|---|---|---|---|
| Leucothoe | 0.037 | 0.251 | 0.502 | 0.773 | |
| Bonnierilla | 0.092 | 0.474 | 0.817 | ||
| Doropygus | 0.579 | 0.078 | |||
| Janstockia | 0.761 | ||||
| Individual sites | |||||
| Abu Tig | Leucothoe | 0.593 | 0.526 | 0.863 | 0.360 |
| Bonnierilla | 0.861 | 0.418 | 0.548 | ||
| Doropygus | 0.641 | 0.774 | |||
| Janstockia | 0.553 | ||||
| Mövenpick | Leucothoe | 0.048 | - | 0.553 | 0.832 |
| Bonnierilla | - | 0.985 | 0.847 | ||
| Doropygus | - | - | |||
| Janstockia | 0.731 | ||||
| Zeytouna Beach | Leucothoe | 0.645 | 0.421 | 0.81 | 0.657 |
| Bonnierilla | <0.001 | 0.950 | 0.741 | ||
| Doropygus | 0.656 | 0.839 | |||
| Janstockia | 0.755 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Rivera, E.; Sherif, M.-E.-D.; El-Sahhar, S.; Lombardi, T. Spatial Variability in a Symbiont-Diverse Marine Host and the Use of Observational Data to Assess Ecological Interactions. Diversity 2022, 14, 197. https://doi.org/10.3390/d14030197
Cruz-Rivera E, Sherif M-E-D, El-Sahhar S, Lombardi T. Spatial Variability in a Symbiont-Diverse Marine Host and the Use of Observational Data to Assess Ecological Interactions. Diversity. 2022; 14(3):197. https://doi.org/10.3390/d14030197
Chicago/Turabian StyleCruz-Rivera, Edwin, Mohy-El-Din Sherif, Salma El-Sahhar, and Thomas Lombardi. 2022. "Spatial Variability in a Symbiont-Diverse Marine Host and the Use of Observational Data to Assess Ecological Interactions" Diversity 14, no. 3: 197. https://doi.org/10.3390/d14030197