
Survey of Volvariella (Agaricales, Basidiomycota) including Two New Species, V. neovolvacea and V. thailandensis, from Northern Thailand



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Morphological Observation
2.3. DNA Extraction, Amplification and Sequencing
2.4. Sequence Alignment
2.5. Phylogenetic Analyses
3. Results
3.1. Sample Collection and Morphological Observation
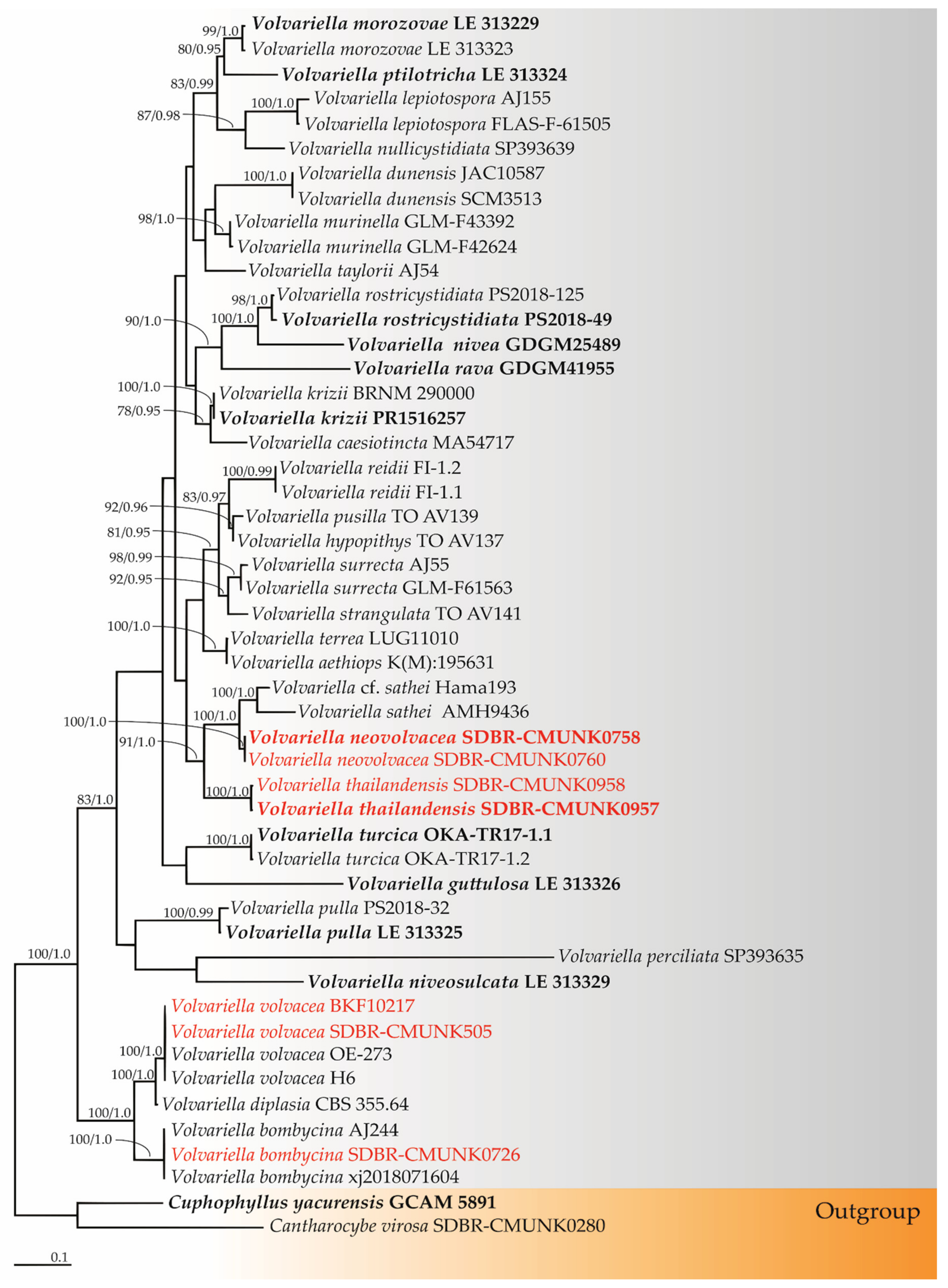
3.2. Phylogenetic Results
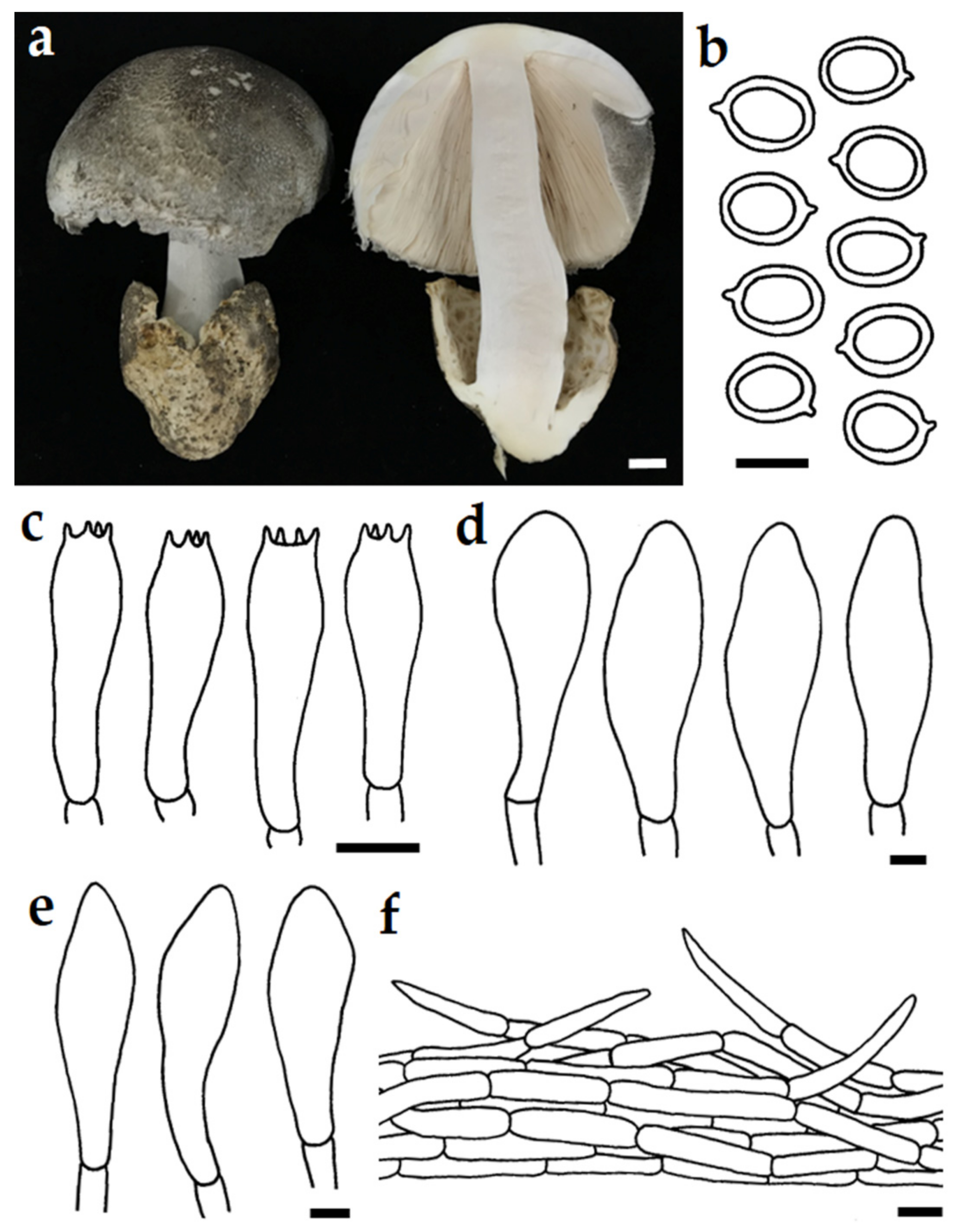
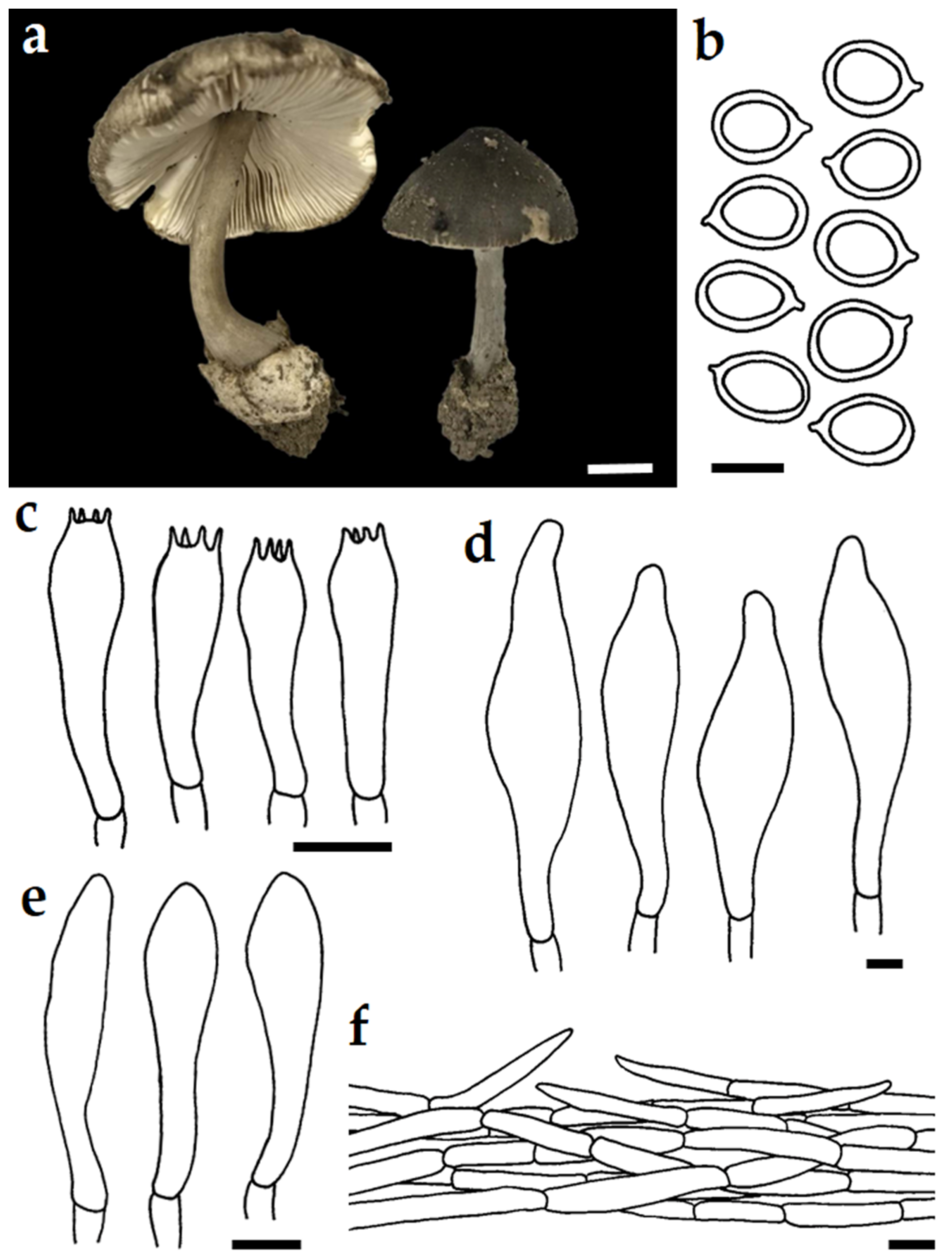
3.3. Taxonomic Description of New Species


4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Priest, M.; Conde, B. The occurrence of Volvariella volvacea (Bull.: Fr.) Singer in the Northern Territory, Australia. Australas. Mycol. 2006, 25, 65–67. [Google Scholar]
- Shaffer, R. Volvariella in North America. Mycologia 1957, 49, 545–579. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Capelari, M. Records and two new species of Volvariella (Pluteaceae, Agaricales) from Brazil. Mycotaxon 2009, 106, 385–398. [Google Scholar]
- Malysheva, E.; Malysheca, V.; Alexandrova, A.; Morozova, O. Observations on Pluteaceae in Vietnam. 1. New species and new records of Volvariella. Phytotaxa 2019, 408, 233–254. [Google Scholar] [CrossRef]
- Pegler, D.N. A preliminary agaric flora of East Africa. Kew Bull. Add. Ser. 1977, 6, 1–615. [Google Scholar]
- Niego, A.G.T.; Sysouphanthong, P.; Thongklang, N.; Hyde, K.D.; Phonemany, M.; Phokkamsak, R.; Raspé, O. A new species of Volvariella and the first record of Volvariella pulla (Agaricales: Incertae sedis) from Thailand. Phytotaxa 2020, 480, 237–250. [Google Scholar] [CrossRef]
- Singer, R. The Agaricales in modern taxonomy. Lilloa 1951, 22, 1–832. [Google Scholar]
- Kirk, P.M.; Cannon, P.; Stalpers, J. (Eds.) Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008; p. 728. [Google Scholar]
- Pegler, D.N. Agaric flora of Sri Lanka. Kew Bull. Add. Ser. 1986, 12, 1–519. [Google Scholar]
- Justo, A.; Castro, M. The genus Volveriella in Spain: V. dunensis comb. & stat. nov. and observations on V. Earlei. Mycotaxon 2010, 112, 261–270. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986; pp. 455–457. [Google Scholar]
- Orton, P.D. British Fungus Flora, Agarics and Boleti 4—Pluteaceae: Pluteus and Volvariella; Royal Botanic Garden: Edinburgh, UK, 1986; p. 99. [Google Scholar]
- Mortimer, P.E.; Bao, E.; Hyde, K.D.; Li, H. Macrofungi as food. Ency. Mycol. 2021, 1, 405–417. [Google Scholar]
- Sakinah, N.M.; Misran, A.; Mahmud, T.M.M.; Abdullah, S. A review: Production and postharvest management of Volvariella volvacea. Int. Food. Res. J. 2019, 26, 367–376. [Google Scholar]
- Lee, J.Y.; Lee, Y.W.; Lim, J. Coloured Illustrations of Fungi of Korea; Baemunkak: Seoul, Korea, 1959; p. 138. [Google Scholar]
- Lee, J.Y. The list of the fungi of Korea. Korean J. Mycol. 1973, 1, 35–43. [Google Scholar]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.-M.; Ge, Z.-W.; Yang, Z.-L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Justo, A.; Vizzini, A.; Minnis, A.M.; Menolli, N., Jr.; Capelari, M.; Rodríguez, O.; Malysheva, E.; Contu, M.; Ghignone, S.; Hibbett, D.S. Phylogeny of the Pluteaceae (Agaricales, Basidiomycota): Taxonomy and character evolution. Fungal Biol. 2011, 115, 1–20. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Chandrasrikul, A.; Suwanarit, P.; Sangwanit, U.; Lunyong, S.; Payapanon, A.; Sanoamuang, N.; Pukahuta, C.; Petcharat, V.; Sardsud, U.; Duengkae, K.; et al. Checklist of Mushrooms (Basidiomycetes) in Thailand; Office of Natural Resources and Environmental Policy and Planning: Bangkok, Thailand, 2011; pp. 112–113. [Google Scholar]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Kumla, J.; Suwannarach, N.; Wannathes, W. Hymenagaricus saisamornae sp. nov. (Agaricales, Basidiomycota) from northern Thailand. Chiang Mai J. Sci. 2021, 48, 827–836. [Google Scholar]
- Jaichaliaw, C.; Kumla, J.; Vadthanarat, S.; Suwannarach, N.; Lumyong, S. Multigene phylogeny and morphology reveal three novel species and a novel record of Agaricus from northern Thailand. Front. Microbiol. 2021, 12, e650513. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Robert, V.; Vu, D.; Amor, A.B.; van de Wiele, N.; Brouwer, C.; Jabas, B.; Szoke, S.; Dridi, A.; Triki, M.; Ben Daoud, S.; et al. MycoBank gearing up for new horizons. IMA Fungus 2013, 4, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar]
- Kumla, J.; Suwannarach, N.; Lumyong, S. Cantharocybe virosa, first record of the genus in Thailand. Mycotaxon 2018, 133, 481–485. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.S.J.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A.; Contu, M.; Justo, A. Additional records of Volvariella dunensis (Basidiomycota, Agaricales): Morphological and molecular characterization. Mycotaxon 2011, 117, 37–43. [Google Scholar] [CrossRef]
- Kaygusuz, O.; Knudsen, H.; Türkekul, I.; Çolak, O.F. Volvariella turcica, a new species from Turkey, and a multigene phylogeny of Volvariella. Mycologia 2020, 112, 577–587. [Google Scholar] [CrossRef]
- Li, T.H.; Chen, X.L.; Shen, Y.H.; Li, T. 2009. A white species of Volvariella (Basidiomycota, Agaricales) from southern China. Mycotaxon 2009, 109, 255–261. [Google Scholar]
- Xu, J.; Li, T.H.; Shen, Y.H.; Zhang, M. Volvariella rava sp. nov. from southern China. Mycotaxon 2015, 130, 857–865. [Google Scholar] [CrossRef]
- Daniëls, P.P.; Hama, O.; Fernández, A.F.; García-Pantaleón, F.I.; Barage, M.; Ibrahim, D.; Alcántara, M.R. First records of some Asian macromycetes in Africa. Mycotaxon 2015, 130, 337–359. [Google Scholar] [CrossRef]
- Senthilarasu, G.; Sharma, R.; Singh, S. A new species of Volvariella from India. Mycotaxon 2012, 119, 467–476. [Google Scholar] [CrossRef]
- Adewoyin, A.G.; Barooah, M.; Oloke, J.K.; Bora, S.S. Identification and physiological properties of a Nigerian strain of Volvariella sp. isolated from oil palm waste. World J. Microbiol. Biotechnol. 2017, 33, 135. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml-vi-hpc: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. 2009. Available online: http://www.phylo.org/sub_sections/portal (accessed on 5 January 2022).
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov Chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary Biology, University of Edinburgh. Available online: http://treebioedacuk/software/figtree/ (accessed on 8 January 2021).
- Favre, A.; Vialard, J. Volvariella aethiops nov. sp. Bull. Mycol. Bot. Dauphiné-Savoie 2007, 47, 19–23. [Google Scholar]
- Halama, M. Volvariella caesiotincta P.D. Orton, a new species in the mycobiota of Poland. Acta Mycol. 2009, 44, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Boekhout, T. Notulae ad floram agaricinam Neerlandicam—XII: Small, saprophytic Volvariella species in the Netherlands. Persoonia 1986, 13, 197–211. [Google Scholar]
- Le, H.; Chu, N. New records of genus Volvariella (Pluteaceae) from Cuc Phuong National Park. Vietnam. J. Sci. Technol. Eng. 2018, 60, 81–88. [Google Scholar] [CrossRef]
- Seok, S.J.; Kim, Y.S.; Weon, H.Y.; Lee, K.H.; Park, K.M.; Min, K.H.; Yoo, K.H. Taxonomic study on Volvariella in Korea. Mycobiology 2002, 30, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Wartchow, F. Volvariella cubensis: A rare neotropical agaric new to South America. Mycotaxon 2009, 107, 181–187. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Taxa | Strain/Voucher | Country | GenBank Accession Number | Reference | |
|---|---|---|---|---|---|
| ITS | nrLSU | ||||
| Cantharocybe virosa | SDBR-CMUNK0280 | Thailand | MG694692 | MG694690 | [31] |
| Cuphophyllus yacurensisT | QCAM5891 | Ecuador | NR166388 | KY780119 | [32] |
| Volvariella aethiops | K(M):195631 | United Kingdom | MZ159529 | – | Unpublished |
| Volvariella bombycina | AJ244 | Spain | HM562212 | HM562256 | [18] |
| Volvariella bombycina | SDBR-CMUNK0726 | Thailand | OM417508 | OM373644 | This study |
| Volvariella bombycina | xj2018071604 | China | MT351048 | – | Unpublished |
| Volvariella caesinotincta | MA54717 | Spain | HM562211 | HM562255 | [18] |
| Volvariella diplasia | CBS 355.64 | India | MH858454 | MH870086 | [21] |
| Volvariella dunensis | JAC10587 | New Zealand | MN738630 | MN738630 | Unpublished |
| Volvariella dunensis | SCM3513 | Spain | JF415140 | – | [33] |
| Volvariella guttulosaT | LE 313326 | Vietnam | – | NG068890 | [4] |
| Volvariella hypopithys | TO AV137 | Italy | HM246492 | HM246488 | Unpublished |
| Volvariella kriziiT | PR1516257 | Czech Republic | – | MK770133 | [34] |
| Volvariella krizii | BRNM 290000 | Czech Republic | – | MK770133 | [34] |
| Volvariella lepiotospora | AJ155 | USA | HM562214 | HM562259 | Unpublished |
| Volvariella lepiotospora | FLAS-F-61505 | USA | MH281874 | – | Unpublished |
| Volvariella morozovaeT | LE 313229 | Vietnam | MF377507 | MF377508 | [32] |
| Volvariella morozovae | LE 313323 | Vietnam | MK882995 | – | [32] |
| Volvariella murinella | GLM:GLM-F42624 | Germany | MK412400 | MK412400 | Unpublished |
| Volvariella murinella | GLM:GLM-F43392 | Germany | MK412361 | – | Unpublished |
| Volvariella neovolvaceaT | SDBR-CMUNK0758 | Thailand | OM417503 | OM373653 | This study |
| Volvariella neovolvacea | SDBR-CMUNK0760 | Thailand | OM417505 | OM417507 | This study |
| Volvariella niveaT | GDGM25489 | China | FJ749127 | – | [35] |
| Volvariella niveosulcataT | LE 313329 | Vietnam | MK882996 | MK883002 | [4] |
| Volvariella nullicystidiata | SP393639 | Brazil | EU920671 | EU920670 | [3] |
| Volvariella perciliata | SP393635 | Brazil | EU920672 | EU920668 | [3] |
| Volvariella ptilotrichaT | LE 313324 | Vietnam | NR164612 | MK882997 | [4] |
| Volvariella pullaT | LE 313325 | Vietnam | NR164613 | MK883003 | [4] |
| Volvariella pulla | MFLU 19-1534 | Thailand | MT074696 | – | [6] |
| Volvariella pusilla | TO AV139 | Italy | HM246494 | HM246479 | Unpublished |
| Volvariella ravaT | GDGM41955 | China | KP784686 | – | [36] |
| Volvariella reidii | F1-1.1 | Finland | MK770139 | MK770131 | [34] |
| Volvariella reidii | F1-1.2 | Finland | MK770140 | MK770132 | [34] |
| Volvariella rostricystidiataT | MFLU 19-1528 | Thailand | MT074694 | – | [6] |
| Volvariella rostricystidiata | MFLU 19-1531 | Thailand | MT074695 | – | [6] |
| Volvariella cf. sathei | Hama193 | Niger | KF926663 | – | [37] |
| Volvariella sathei | AMH 9436 | India | JN792550 | – | [38] |
| Volvariella strangulata | TO AV141 | Italy | HM246493 | HM246484 | Unpublished |
| Volvariella surrecta | GLM:GLM-F61563 | Germany | MK412358 | MK412358 | Unpublished |
| Volvariella surrecta | AJ55 | Spain | HM562213 | HM562254 | [18] |
| Volvariella taylorii | AJ54 | Portugal | HM562210 | HM562260 | [18] |
| Volvariella terrea | LUG11010 | France | JF415141 | – | [33] |
| Volvariella thailandensisT | SDBR-CMUNK0957 | Thailand | OM417510 | OM417509 | This study |
| Volvariella thailandensis | SDBR-CMUNK0958 | Thailand | OM417510 | OM386669 | This study |
| Volvariella turcicaT | OKA-TR17.1 | Turkey | MK770135 | MK770128 | [34] |
| Volvariella turcica | OKA-TR17.2 | Turkey | MK770136 | MK770129 | [34] |
| Volvariella volvacea | H6 | Nigeria | KC894929 | – | [39] |
| Volvariella volvacea | OE-273 | India | KC14211 | – | Unpublished |
| Volvariella volvacea | BKF10217 | Thailand | MN492647 | OM373563 | This study |
| Volvariella volvacea | SDBR-CMUNK505 | Thailand | OM417506 | OM373623 | This study |
| Volvariella Species | Distribution | Pileus (mm in Diameter) | Basidiospore (μm) | Basidia (μm) | Pleurocystidia (μm) | Cheilocystidia (μm) | Reference |
|---|---|---|---|---|---|---|---|
| V. aethiops | France | 30–50 | 7.0–11 × 3.7–7.2 | NR | 60–100 × 10–25 | 60–100 × 10–25 | [48] |
| V. bakeri | USA | 100 | 6.9–9.3 × 4.6–6.9 | 21–35 × 7–11 | 47–71 × 11–31 | 27–71 × 11–31 | [2,52] |
| V. caesiotincta | Europe and North Africa | 44 | 6.2–7.7 × 4.2–5.4 | 21–28 × 6.5–9 | 35–46 × 10–25 | 34–65 × 9.6–19 | [18,49] |
| V. morozovae | Vietnam | 30–40 | 5.0–6.2 × 3.3–4.3 | 13.5–20 × 7–8.5 | 45–50 × 18–30 | 40–50 × 13–18 | [32] |
| V. murinella | Europe and Vietnam | 30–60 | 4.8–5.6 × 4.9–6.6 | 19–25 × 6–7 | 20–30 × 10–15 | 45–65 × 8–15 | [50,51] |
| V. neovolvacea | Thailand | 65–80 | 6.5–8.0 × 5.0–7.0 | 27–40 × 7.5–11 | 45–98 × 23–77 | 29–57 × 9–22 | This study |
| V. pseudovolvacea | Sri Lanka | 30–60 | 4.2–5.2 × 2.5–3.5 | 14–16 × 4.5–6.5 | 32–45 × 12–16 | 30–38 × 12–14 | [9] |
| V. pulla | Thailand and Vietnam | 40–75 | 5.5–8.0 × 4.0–5.0 | 16–19.5 × 7–9 | 25–56 × 6–32 | 24–87 × 9–17 | [4,6] |
| V. rava | China | 25–60 | 4.0–6.5 × 4.0–5.0 | 24–30 × 7–10 | 35–71 × 11–30 | 55–80 × 15–25 | [36] |
| V. rostricystidiata | Thailand | 55–85 | 5.0–7.0 × 3.5–5.5 | 17.5–29 × 7–10 | 41–117 × 10–25 | 51–80 × 14–39 | [6] |
| V. taylorii | Europe, South Korea, USA and Vietnam | 20–50 | 5.0–7.0 × 3.0–4.8 | 20–24 × 6–8 | 35–49 × 8–10 | 28–43 × 9–13 | [18,51,52] |
| V. terastia | Sri Lanka | 60–110 | 5.0–6.5 × 4.2–5.2 | 17–23 × 5.5–7 | 35–50 × 15–22 | 32–36 × 10–13 | [9] |
| V. thailandensis | Thailand | 35–50 | 5.5–7.0 × 4.0–6.0 | 15–35 × 6–8 | 35–77 × 17–30 | 32–48 × 6–10 | This study |
| V. volvacea | Asia and East Africa | 50–100 | 6.0–9.0 × 4.7–5.7 | 30–35 × 7.5–9.0 | 45–120 × 17–30 | 40–70 × 10–15 | [9,51], This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumla, J.; Suwannarach, N.; Wannathes, N.; Lumyong, S. Survey of Volvariella (Agaricales, Basidiomycota) including Two New Species, V. neovolvacea and V. thailandensis, from Northern Thailand. Diversity 2022, 14, 161. https://doi.org/10.3390/d14030161
Kumla J, Suwannarach N, Wannathes N, Lumyong S. Survey of Volvariella (Agaricales, Basidiomycota) including Two New Species, V. neovolvacea and V. thailandensis, from Northern Thailand. Diversity. 2022; 14(3):161. https://doi.org/10.3390/d14030161
Chicago/Turabian StyleKumla, Jaturong, Nakarin Suwannarach, Nopparat Wannathes, and Saisamorn Lumyong. 2022. "Survey of Volvariella (Agaricales, Basidiomycota) including Two New Species, V. neovolvacea and V. thailandensis, from Northern Thailand" Diversity 14, no. 3: 161. https://doi.org/10.3390/d14030161