
Diversification in the Comoros: Review of the Laccophilus alluaudi Species Group with the Description of Four New Species (Coleoptera: Dytiscidae) †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
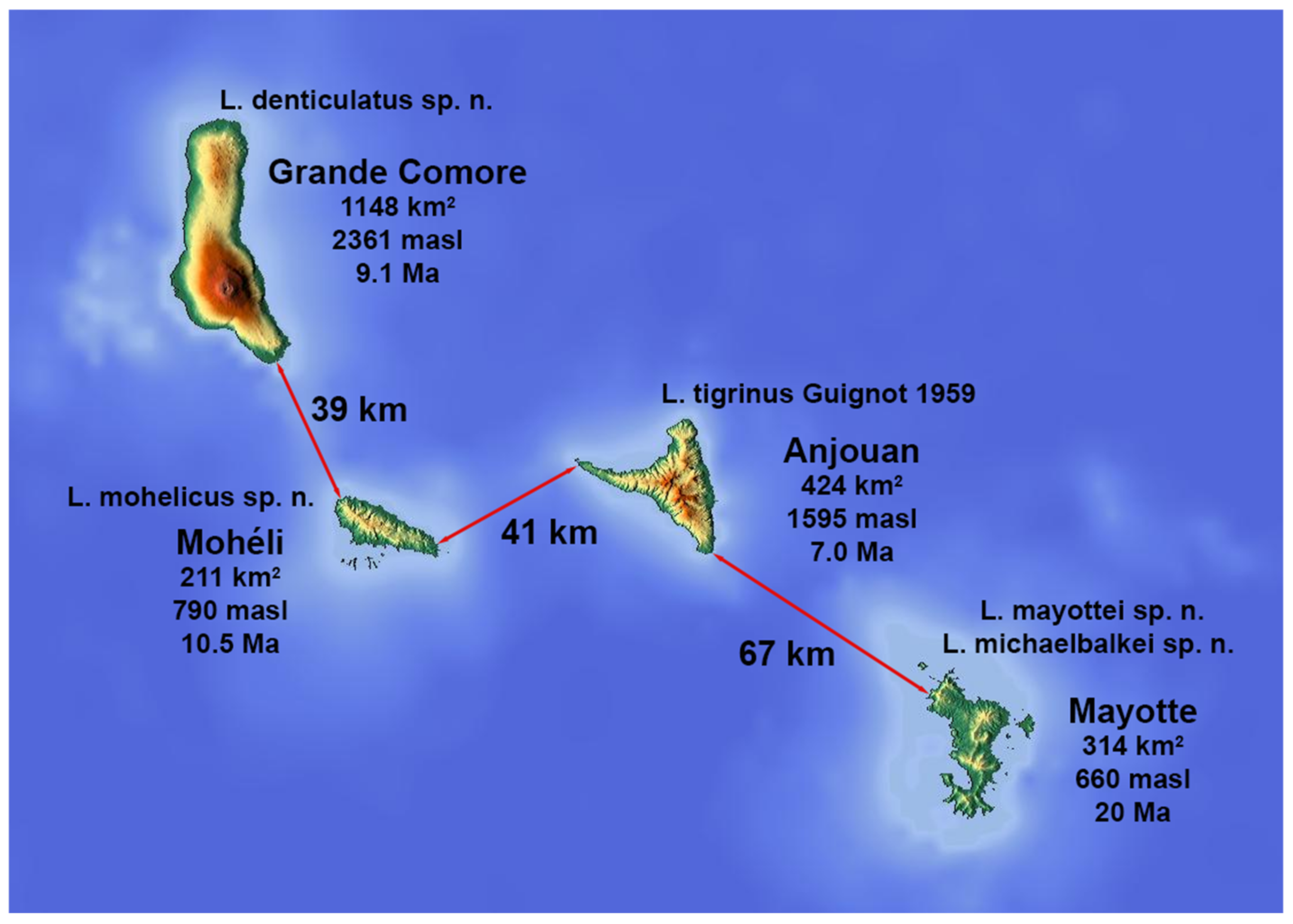
:1. Introduction
2. Material and Methods
2.1. Preparation Technique and Measurements
2.2. Photography
2.3. Depositories
- IRSNB—Institut Royal des Sciences Naturelles de Belqique, Brussels, Belgium
- MRAC—Musée Royal de l’Afrique centrale, Tervuren, Belgium
- MNHN—Muséum National d’Historie Naturelle, Paris, France
- MZH—Finnish Museum of Natural History, Helsinki, Finland
- NHRS—Swedish Museum of Natural History, Stockholm, Sweden
- ZSM—Zoologische Staatsammlung, München, Germany.
3. Results
3.1. Key to Species of L. alluaudi Species Group in the Comoros
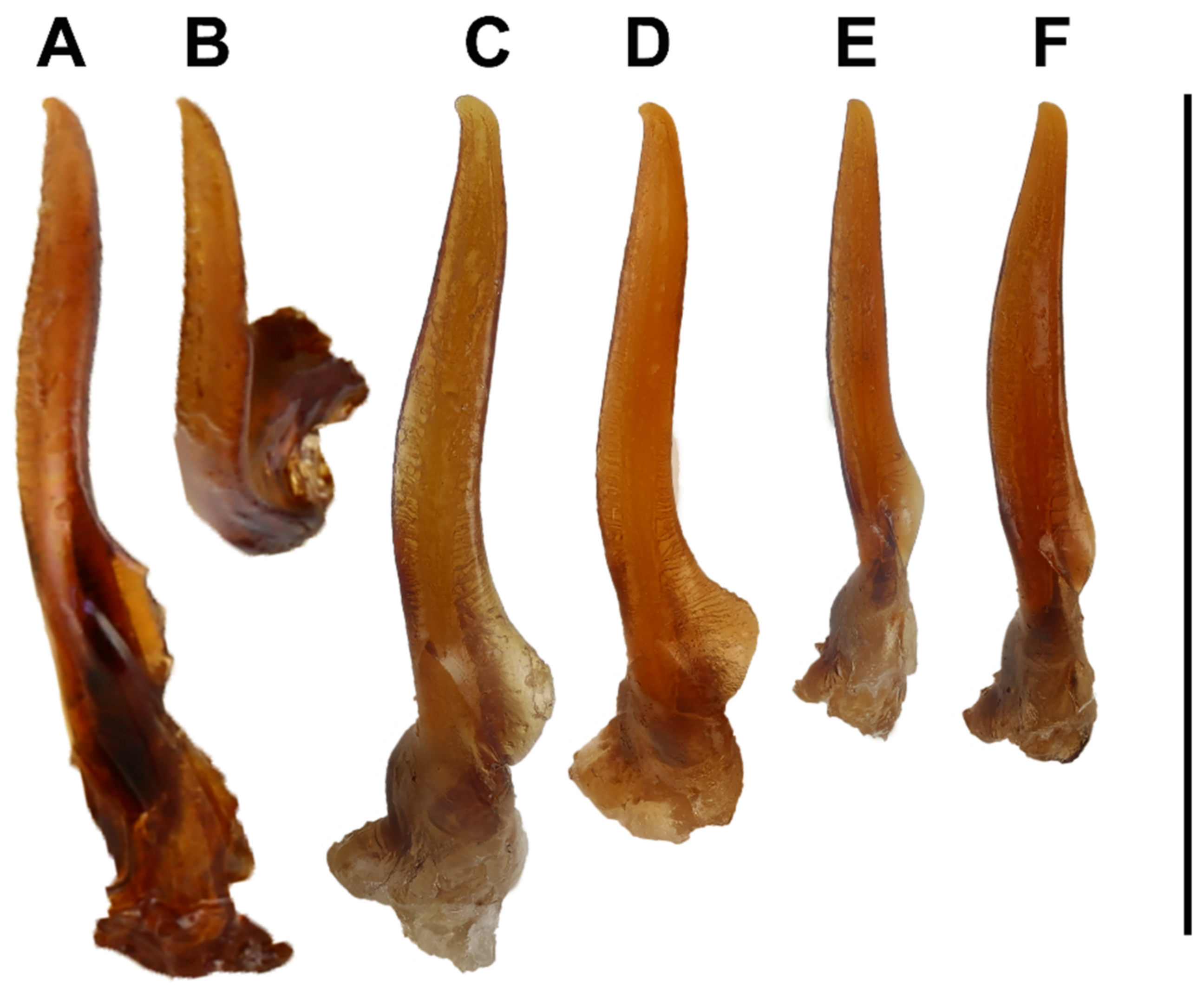
- Pronotum anteriorly and head posteriorly darker; ferruginous to dark brown (Figure 2A–D) (rarely dark area indistinct in old, dry specimens) ……… 2Pronotum and head unicolored testaceous to pale ferruginous (Figure 2E,F) ……… 4
- Smaller species (body length 3.36–3.76 mm); dark area on head and pronotum ferruginous; gradual delimitation of markings (rarely dark markings almost absent) (Figure 2D) (Mohéli) ……… L. mohelicusLarger species (body length 3.98–4.12 mm); dark area on head and pronotum darker and more distinctly delimited (Figure 2A–C) (Grande Comore, Anjouan) ……… 3
3.2. Taxonomic Treatment
Laccophilus tigrinus Guignot, 1959
Laccophilus denticulatus n. sp.
Laccophilus mohelicus n. sp.
Laccophilus mayottei n. sp.
Laccophilus michaelbalkei n. sp.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Grant, P.R. Ecology and Evolution of Darwin’s Finches, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Emerson, B.C. Evolution on oceanic islands: Molecular phylogenetic approaches to understanding pattern and process. Mol. Ecol. 2002, 11, 951–966. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Triantis, K.A.; Ladle, R.J. A general dynamic theory of oceanic island biogeography. J. Biogeogr. 2008, 35, 977–994. [Google Scholar] [CrossRef]
- Gillespie, R.G. Oceanic islands: Models of diversity. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2001; pp. 590–599. [Google Scholar]
- Gillespie, R.G.; Baldwin, B.G.; Waters, J.M.; Fraser, C.I.; Nikula, R.; Roderick, G.K. Long-distance dispersal: A framework for hypothesis testing. Trends Ecol. Evol. 2012, 27, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.R.; Meiri, S. Biodiversity growth on the volcanic ocean islands and the roles of in situ cladogenesis and immigration; case with the reptiles. Ecography 2019, 42, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.; Rocha, S. Comoros. In Encyclopedia of Islands; Gillespie, R., Clague, D., Eds.; University of California Press: Oakland, CA, USA, 2009; pp. 177–180. [Google Scholar]
- Michon, L. The volcanism of the Comoros archipelago integrated at a regional scale. In Active Volcanoes of the Southwest Indian Ocean, Piton de la Fournaise and Karthala; Bachelery, P., Lénat, J.-F., Di Muro, A., Michon, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 233–244. [Google Scholar]
- Warren, B.H.; Bermingham, E.; Bowie, R.C.K.; Prys-Jones, R.P.; Christophe Thébaud, C. Molecular phylogeography reveals island colonization history and diversification of western Indian Ocean sunbirds (Nectarinia: Nectariniidae). Mol. Phyl. Evol. 2003, 29, 67–85. [Google Scholar] [CrossRef]
- Hawlitschek, O.; Toussaint, E.F.A.T.; Gehring, P.S.; Ratsoavina, F.M.; Cole, N.; Crottini, A.; Nopper, J.; Lam, A.W.; Vences, M.; Glaw, F. Gecko phylogeography in the Western Indian Ocean region: The oldest clade of Ebenavia inunguis lives on the youngest island. J. Biogeogr. 2017, 44, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Adler, G.H. Avifaunal diversity and endemism on tropical Indian Ocean islands. J. Biogeogr. 1994, 21, 85–95. [Google Scholar] [CrossRef]
- Rocha, S.; Carretero, M.A.; Harris, J.D. Mitochondrial DNA sequence data suggests two independent colonizations of the Comoros archipelago by Chameleons of the genus Furcifer. Belg. J. Zool. 2005, 135, 39–42. [Google Scholar]
- Rocha, S.; Carretero, M.A.; Harris, J.D. Diversity and phylogenetic relationships of Hemidactylus geckos from the Comoro Islands. Mol. Phyl. Evol. 2005, 35, 292–299. [Google Scholar] [CrossRef]
- Rocha, S.; Carretero, M.A.; Vences, M.; Glaw, F.; Harris, J.D. Decipering patterns of transoceanic dispersal: The evolutionary origin and biogeography of coastal lizards (Cryptoblepharus) in the Western Indian Ocean Region. J. Biogeogr. 2006, 33, 13–22. [Google Scholar] [CrossRef]
- Rocha, S.; Posada, D.; Carretero, M.A.; Harris, J.D. Phylogenetic affinities of Comoran and East African day geckos (genus Phelsuma): Multiple natural colonisations, indtroductions and island radiations. Mol. Phyl. Evol. 2007, 43, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschek, O.; Nagy, Z.T.; Glaw, F. Island evolution and systematic revision of Comoran snakes: Why and when subspecies still make sense. PLoS ONE 2012, 7, e42970. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschek, O.; Ramírez Garrido, S.; Glaw, F. How marine currents influenced the widespread natural overseas dispersal of reptiles in the Western Indian Ocean region. J. Biogeogr. 2017, 44, 1435–1440. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.; Pons, J.-M.; Goodman, S.M.; Bretagnolle, V.; Melo, M.; Bowie, R.C.K.; Currie, D.; Safford, R.; Virani, M.Z.; Thomsett, S.; et al. Tracing the colonization history of the Indian Ocean scops-owls (Strigiformes: Otus) with further insight into the spatio-temporal origin of the Malagasy avifauna. BMC Evol. Biol. 2008, 8, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, J.; Lemoine, D.; Parra, J.L.; Pons, J.-M.; Raherilalao, M.J.; Prys-Jones, R.; Thebaud, C.; Warren, B.H.; Goodman, S.M. Long-distance dispersal and inter-island colonization across the western Malagasy Region explain diversification in brush-warblers (Passeriformes: Nesillas). Biol. J. Linn. Soc. 2016, 119, 873–889. [Google Scholar] [CrossRef]
- Glaw, F.; Hawlitschek, O.; Glaw, K.; Vences, M. Integrative evidence confirms new endemic island frogs and transmarine dispersal of amphibians between Madagascar and Mayotte (Comoros archipelago). Sci. Nat. 2019, 106, 19. [Google Scholar] [CrossRef]
- Schipper, J. Southern Africa: Island Group between Madagascar. 2021. Available online: https://www.worldwidllife.org/ecoregions/at0105 (accessed on 16 December 2021).
- Safford, R.J. The Comoros. In Important Bird Areas in Africa and Associated Islands: Priority Sites for Conservation; Fishpool, L.D.C., Evans, M.I., Eds.; BirdLife Conservation Series No. 11; Pisces Publications: Newbury, UK; BirdLife International: Cambridge, UK, 2001; pp. 411–464. [Google Scholar]
- Hawlitschek, O.; Brückmann, B.; Berger, J.; Green, K.; Glaw, F. Integrating field surveys and remote sensing data to study distribution, habitat use and conservation status of the herpetofauna of the Comoro Islands. ZooKeys 2011, 144, 21–79. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kents, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Agnarsson, I.; Kuntner, M. The generation of a biodiversity hotspot: Biogeography and phylogeography of the western Indian ocean islands. In Current Topics in Phylogenetics and Phylogeography of Terrestrial and Aquatic Systems; Anamthawat-Jonsson, K., Ed.; InTech Publishers: Rijeka, Croatia, 2012; pp. 33–82. [Google Scholar]
- Wikström, N.; Avino, M.; Razafimandimbison, S.G.; Bremer, B. Historical biogeography of the coffee family (Rubiaceae, Gentianales) in Madagascar: Case studies from the tribes Knoxieae, Naucleeae, Paederieae and Vanguerieae. J. Biogeogr. 2010, 37, 1094–1113. [Google Scholar] [CrossRef]
- Goodman, S.M.; Weyeneth, N.; Ibrahim, Y.; Said, I.; Ruedi, M. A review of the bat fauna of the Comoro Archipelago. Acta Chiropterologica 2010, 12, 117–141. [Google Scholar] [CrossRef]
- Miller, K.B.; Bergsten, J. Diving Beetles of the World. Systematics and Biology of the Dytiscidae; Johns Hopkins University Press: Baltimore, MD, USA, 2016; 320p. [Google Scholar]
- Biström, O.; Nilsson, A.N.; Bergsten, J. Taxonomic revision of Afrotropical Laccophilus Leach, 1815 (Coleoptera, Dytiscidae). ZooKeys 2015, 542, 1–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuel, M.; Ramahandrison, A.T. Four new species of the diving beetle genus Laccophilus Leach, 1815 from Madagascar (Coleoptera, Dytiscidae, Laccophilini). Zootaxa 2020, 4822, zootaxa-4822. [Google Scholar] [CrossRef] [PubMed]
- Bergsten, J.; Manuel, M.; Ranarilalatiana, T.; Ramahandrison, A.T.; Hájek, J. Dytiscidae, diving beetles, tsikovoka. In The New Natural History of Madagascar; Goodman, S.M., Ed.; Princeton University Press: Princeton, NJ, USA, in press.
- Guignot, F. Dytiscides et Gyrinides des Comores. Mem. L’institute Sci. Madag. 1959, 10, 75–79. [Google Scholar]
- Guignot, F. Revision des hydrocanthares d’Afrique (Coleoptera Dytiscoidea), 3. In Annales du Musée Royal du Congo Belge; Série 8vo Sciences Zoologiques; Musée Royal du Congo Belge: New York, NY, USA, 1961; Volume 90, pp. 653–995. [Google Scholar]
- Wewalka, G. Results of the Austrian Hydrobiological Mission, 1974, to the Seychelles-, Comores- and Mascarene Archipelagos. Part VII: Dytiscidae, Gyrinidae (Coleoptera). Ann. Nat. Mus. Wien 1980, 83, 723–732. [Google Scholar]
- Nilsson, A.N. Dytiscidae. In World Catalogue of Insects; Apollo Books: Stenstrup, Denmark, 2001; Volume 3, pp. 1–395. [Google Scholar]
- Nilsson, A.N.; Hajek, J. A World Catalogue of the Family Dytiscidae, or the Diving Beetles (Coleoptera, Adephaga). Version 1.I.2021. 2021, pp. 1–315. Available online: www.waterbeetles.eu (accessed on 4 November 2021).
- Ibouroi, M.T.; Dhurham, S.A.O.; Besnard, A.; Lescureux, N. What is the main driver of unsustainable natural resource use in the Comoro Islands? bioRxiv 2021. [Google Scholar] [CrossRef]
- Le Ministère de la Transition Écologique 2021. Création d’une Réserve Naturelle Nationale des Forêts à Mayotte. Available online: https://biodiversite.gouv.fr/actualite/creation-dune-reserve-naturelle-nationale-des-forets-mayotte (accessed on 16 December 2021).




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergsten, J.; Biström, O. Diversification in the Comoros: Review of the Laccophilus alluaudi Species Group with the Description of Four New Species (Coleoptera: Dytiscidae). Diversity 2022, 14, 81. https://doi.org/10.3390/d14020081
Bergsten J, Biström O. Diversification in the Comoros: Review of the Laccophilus alluaudi Species Group with the Description of Four New Species (Coleoptera: Dytiscidae). Diversity. 2022; 14(2):81. https://doi.org/10.3390/d14020081
Chicago/Turabian StyleBergsten, Johannes, and Olof Biström. 2022. "Diversification in the Comoros: Review of the Laccophilus alluaudi Species Group with the Description of Four New Species (Coleoptera: Dytiscidae)" Diversity 14, no. 2: 81. https://doi.org/10.3390/d14020081