
Microplastics Occurrence in the European Common Frog (Rana temporaria) from Cottian Alps (Northwest Italy)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Water and Sediment Sampling
2.3. Sampling and Processing of Periphyton, Macroinvertebrates, Tadpoles, and Adult Frogs
2.4. Microplastic Determination
2.5. Quality Assurance/Quality Control (QA/QC)
2.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boucher, J.; Friot, D. Primary Microplastics in the Oceans: A Global Evaluation of Sources; IUCN: Gland, Switzerland, 2017; Available online: https://portals.iucn.org/library/sites/library/files/documents/2017-002-En.pdf (accessed on 3 November 2021).
- Blettler, M.C.; Wantzen, K.M. Threats underestimated in freshwater plastic pollution: Mini-review. Water Air Soil Pollut. 2017, 230, 174. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, P.; Manna, C.; Jain, M. Abundance, interaction, ingestion, ecological concerns, and mitigation policies of microplastic pollution in riverine ecosystem: A review. Sci. Total Environ. 2021, 782, 146695. [Google Scholar] [CrossRef]
- Mallik, A.; Xavier, K.M.; Naidu, B.C.; Nayak, B.B. Ecotoxicological and physiological risks of microplastics on fish and their possible mitigation measures. Sci. Total Environ. 2021, 779, 146433. [Google Scholar] [CrossRef]
- Plastic Europe. Plastics-The Facts 2013: An Analysis of European Latest Plastics Production, Demand and Waste Data. Available online: https://www.plasticseurope.org/application/files/7815/1689/9295/2013plastics_the_facts_PubOct2013.pdf (accessed on 3 November 2021).
- Plastics Europe. An Analysis of European Plastics Production, Demand and Waste Data. 2020. Available online: https://www.plasticseurope.org/it/resources/publications/4312-plastics-facts-2020 (accessed on 3 November 2021).
- Thompson, R.C.; Swan, S.H.; Moore, C.J.; von Saal, F.S. Our plastic age. Philos. Trans. R. Soc. B 2009, 364, 1973–1976. [Google Scholar] [CrossRef] [Green Version]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 0116. [Google Scholar] [CrossRef]
- Amobonye, A.; Bhagwat, P.; Singh, S.; Pillai, S. Plastic biodegradation: Frontline microbes and their enzymes. Sci. Total Environ. 2021, 759, 143536. [Google Scholar] [CrossRef]
- Gigault, J.; Halle, A.; Baudrimont, M.; Pascal, P.Y.; Gauffre, F.; Phi, T.L.; El Hadri, H.; Grassl, B.; Reynaud, S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Vendel, A.L.; Bessa, F.; Alves, V.E.N.; Amorim, A.L.A.; Patrício, J.; Palma, A.R.T. Widespread microplastic ingestion by fish assemblages in tropical estuaries subjected to anthropogenic pressures. Mar. Pollut. Bull. 2017, 117, 448–455. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, X.; Huang, W.; Li, J.; Wang, C.; Zhang, D.; Zhang, C. Microplastic pollution in deep-sea sediments and organisms of the Western Pacific Ocean. Environ. Pollut. 2020, 259, 113948. [Google Scholar] [CrossRef]
- Pastorino, P.; Pizzul, E.; Bertoli, M.; Anselmi, S.; Kušće, M.; Menconi, V.; Prearo, M.; Renzi, M. First insights into plastic and microplastic occurrence in biotic and abiotic compartments, and snow from a high-mountain lake (Carnic Alps). Chemosphere 2020, 265, 129121. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, N. Mechanistic implications of plastic degradation. Polym. Degrad. Stabil. 2008, 93, 561–584. [Google Scholar] [CrossRef]
- Waldschläger, K.; Lechthaler, S.; Stauch, G.; Schüttrumpf, H. The way of microplastic through the environment–Application of the source-pathway-receptor model. Sci. Total Environ. 2020, 713, 136584. [Google Scholar] [CrossRef]
- Bertoli, M.; Pastorino, P.; Lesa, D.; Renzi, M.; Anselmi, S.; Prearo, M.; Pizzul, E. Microplastics accumulation in functional feeding guilds and functional habit groups of freshwater macrobenthic invertebrates: Novel insights in a riverine ecosystem. Sci. Total Environ. 2021, 804, 150207. [Google Scholar] [CrossRef]
- Pastorino, P.; Prearo, M. High-Mountain Lakes, Indicators of Global Change: Ecological Characterization and Environmental Pressures. Diversity 2020, 12, 260. [Google Scholar] [CrossRef]
- Becker, C.G.; Fonseca, C.R.; Haddad, C.F.B.; Batista, R.F.; Prado, P.I. Habitat split and the global decline of amphibians. Science 2007, 318, 1775–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, S.; Muehlbauer, J.D.; Marti, E. Resource subsidies between stream and terrestrial ecosystems under global change. Glob. Chang. Biol. 2016, 22, 2489–2504. [Google Scholar] [CrossRef]
- Johansson, M.; Primmer, C.R.; Merilä, J. Does habitat fragmentation reduce fitness and adaptability? A case study of the common frog (Rana temporaria). Mol. Ecol. 2007, 16, 2693–2700. [Google Scholar] [CrossRef]
- Marchesini, A.; Ficetola, G.F.; Cornetti, L.; Battisti, A.; Vernesi, C. Fine-scale phylogeography of Rana temporaria (Anura: Ranidae) in a putative secondary contact zone in the southern Alps. Biol. J. Linn. Soc. 2017, 122, 824–837. [Google Scholar] [CrossRef] [Green Version]
- Pastorino, P.; Polazzo, F.; Bertoli, M.; Santi, M.; Righetti, M.; Pizzul, E.; Prearo, M. Consequences of fish introduction in fishless Alpine lakes: Preliminary notes from a sanitary point of view. Turk. J. Fish. Aquat. Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Rondinini, C.; Battistoni, A.; Peronace, V.; Teofili, C. Lista Rossa IUCN dei Vertebrati Italiani, Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy. Available online: http://www.iucn.it/pdf/Comitato_IUCN_Lista_Rossa_dei_vertebrati_italiani.pdf (accessed on 3 November 2021).
- Da Costa Araújo, A.P.; de Melo, N.F.S.; de Oliveira Junior, A.G.; Rodrigues, F.P.; Fernandes, T.; de Andrade Vieira, J.E.; Rocha, T.L.; Malafaia, G. How much are microplastics harmful to the health of amphibians? A study with pristine polyethylene microplastics and Physalaemus cuvieri. J. Hazard. Mater. 2020, 382, 121066. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Araújo, A.P.; Rocha, T.L.; e Silva, D.D.M.; Malafaia, G. Micro (nano)plastics as an emerging risk factor to the health of amphibian: A scientometric and systematic review. Chemosphere 2021, 283, 131090. [Google Scholar] [CrossRef]
- Di Nicola, M.R.; Cavigioli, L.; Luiselli, L.; Andreone, F. Anfibi e Rettili d’Italia; Edizioni Belvedere: Roma, Italy, 2019; pp. 1–576. [Google Scholar]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Council of Europe, Strasbourg. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 10 September 2021).
- Ziajahromi, S.; Neale, P.A.; Rintoul, L.; Leusch, F.D.L. Wastewater treatment plants as a pathway for microplastics: Development of a new approach to sample wastewayter-based microplastics. Water Res. 2017, 112, 93–99. [Google Scholar] [CrossRef]
- Galgani, F.; Hanke, G.; Werner, S.; Oosterbaan, L.; Nilsson, P.; Fleet, D.; Kinsey, S.; Thompson, R.C.; van Franeker, J.; Vlachogianni, T.; et al. Guidance on Monitoring of Marine Litter in European Seas; Publications Office of the European Union: Luxembourg, 2014; pp. 1–128. [Google Scholar]
- Enders, K.; Lenz, R.; do Sul, J.A.I.; Tagg, A.S.; Labrenz, M. When every particle matters: A QuEChERS approach to extract microplastics from environmental samples. MethodsX 2020, 7, 100784. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Nor, N.H.M.; Hermsen, E.; Kooi, M.; Mintenig, S.M.; De France, J. Microplastics in freshwaters and drinking water: Critical review and assessment of data quality. Water Res. 2019, 155, 410–422. [Google Scholar] [CrossRef]
- Jâms, I.B.; Windsor, F.M.; Poudevigne-Durance, T.; Ormerod, S.J.; Durance, I. Estimating the size distribution of plastics ingested by animals. Nat. Commun. 2020, 11, 1594. [Google Scholar] [CrossRef] [Green Version]
- Smalling, K.L.; Anderson, C.W.; Honeycutt, R.K.; Cozzarelli, I.M.; Preston, T.; Hossack, B.R. Associations between environmental pollutants and larval amphibians in wetlands contaminated by energy-related brines are potentially mediated by feeding traits. Environ. Pollut. 2019, 248, 260–268. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Abrahams, C.; Meilink, W.R.; Iosif, R.; Zuiderwijk, A. Amphibian decline, pond loss and reduced population connectivity under agricultural intensification over a 38 year period. Biodivers. Conserv. 2017, 26, 1411–1430. [Google Scholar] [CrossRef]
- Trakimas, G.; Jardine, T.D.; Barisevičiūtė, R.; Garbaras, A.; Skipitytė, R.; Remeikis, V. Ontogenetic dietary shifts in European common frog (Rana temporaria) revealed by stable isotopes. Hydrobiologia 2011, 675, 87–95. [Google Scholar] [CrossRef]
- Merritt, R.W.; Cummins, K.W. An Introduction to the Aquatic Insects of North America, 3rd ed.; Hunt Publishing Company: Kendal, UK, 1996. [Google Scholar]
- Baldwin, A.K.; Spanjer, A.R.; Rosen, M.R.; Thom, T. Microplastics in Lake Mead National Recreation Area, USA: Occurrence and biological uptake. PLoS ONE 2020, 15, e0228896. [Google Scholar]
- Pastorino, P.; Prearo, M.; Anselmi, S.; Menconi, V.; Bertoli, M.; Dondo, A.; Pizzul, E.; Renzi, M. Use of the Zebra Mussel Dreissena polymorpha (Mollusca, Bivalvia) as a Bioindicator of Microplastics Pollution in Freshwater Ecosystems: A Case Study from Lake Iseo (North Italy). Water 2021, 13, 434. [Google Scholar] [CrossRef]
- Kolenda, K.; Kuśmierek, N.; Pstrowska, K. Microplastic ingestion by tadpoles of pond-breeding amphibians—first results from Central Europe (SW Poland). Environ. Sci. Pollut. Res. 2020, 27, 33380–33384. [Google Scholar] [CrossRef]
- Dris, R.; Gasperi, J.; Saad, M.; Mirande, C.; Tassin, B. Synthetic fibers in atmospheric fallout: A source of microplastics in the environment? Mar. Pollut. Bull. 2016, 104, 290–293. [Google Scholar] [CrossRef]
- Cera, A.; Cesarini, G.; Scalici, M. Microplastics in freshwater: What is the news from the world? Diversity 2020, 12, 276. [Google Scholar] [CrossRef]
- Hu, L.; Chernick, M.; Hinton, D.E.; Shi, H. Microplastics in small waterbodies and tadpoles from Yangtze River Delta, China. Environ. Sci. Technol. 2018, 52, 8885–8893. [Google Scholar] [CrossRef]
- Karaoğlu, K.; Gül, S. Characterization of microplastic pollution in tadpoles living in small water-bodies from Rize, the northeast of Turkey. Chemosphere 2020, 255, 126915. [Google Scholar] [CrossRef]
- Hu, L.; Su, L.; Xue, Y.; Mu, J.; Zhu, J.; Xu, J.; Shi, H. Uptake, accumulation and elimination of polystyrene microspheres in tadpoles of Xenopus tropicalis. Chemosphere 2016, 164, 611–617. [Google Scholar] [CrossRef]
- Boyero, L.; López-Rojo, N.; Bosch, J.; Alonso, A.; Correa-Araneda, F.; Pérez, J. Microplastics impair amphibian survival, body condition and function. Chemosphere 2020, 244, 125500. [Google Scholar] [CrossRef]
- Rochman, C.M. The complex mixture, fate and toxicity of chemicals associated with plastic debris in the marine environment. In Marine Anthropogenic Litter; Springer: Cham, Switzerland, 2015; pp. 117–140. [Google Scholar]
- Beebee, T.J.; Griffiths, R.A. The amphibian decline crisis: A watershed for conservation biology? Biol. Conserv. 2005, 125, 271–285. [Google Scholar] [CrossRef]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Organ. 2010, 92, 93–99. [Google Scholar] [CrossRef]
- Kestemont, B. The bottom-up assessment of threatened species. Nat. Conserv. Res. 2019, 4, 93–106. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
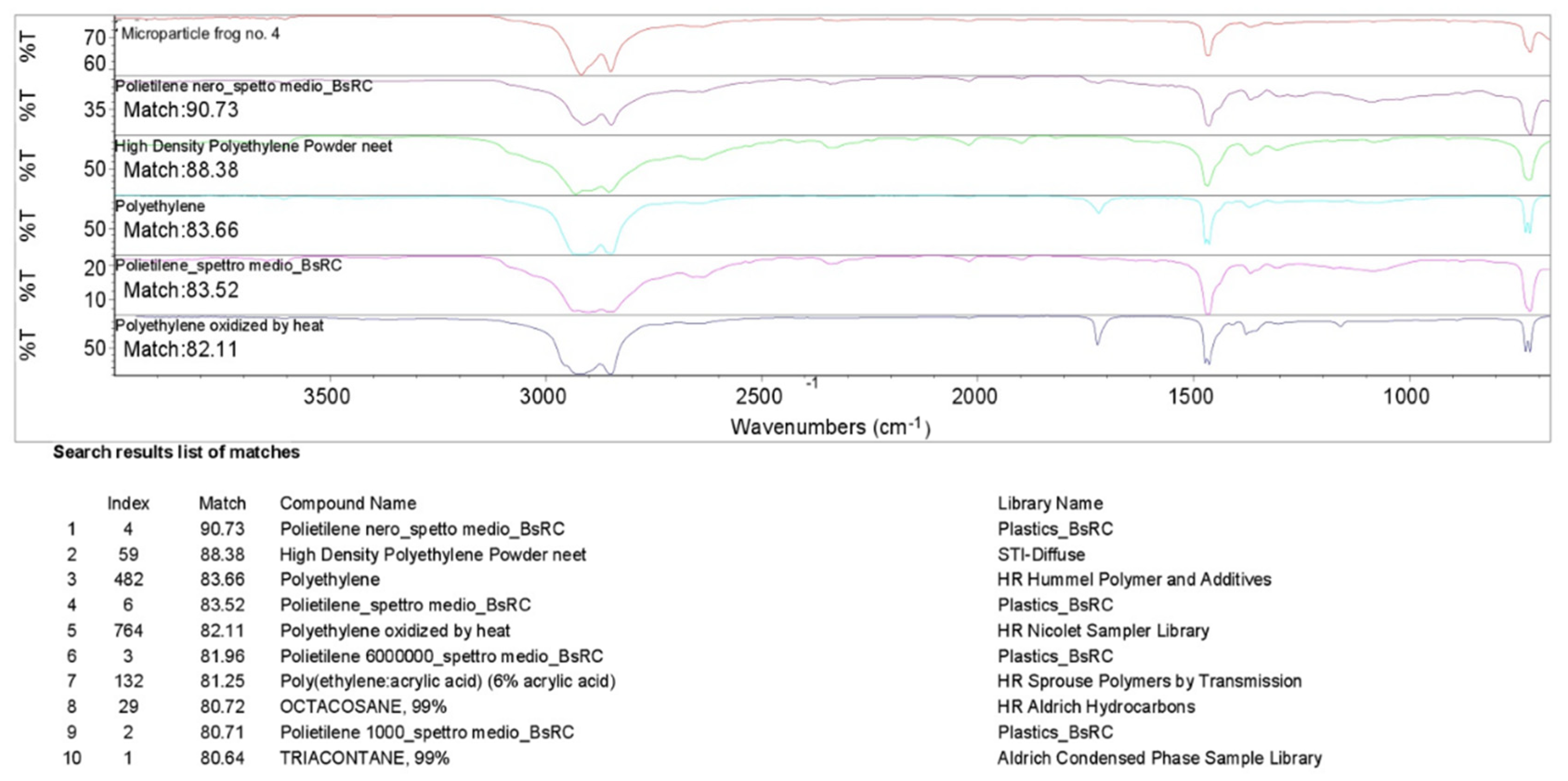{kind=link}
| Specimen | Snout–Vent Length (cm) | Body Mass (g) | Total Items | Shape | Color | Size (µm) | Chemical Composition |
|---|---|---|---|---|---|---|---|
| 1 | 9.0 | 94 | 1 | fibre | blue | 2056.70 | PA |
| 2 | 8.4 | 89 | 1 | fibre | light-blue | 1049.74 | PA |
| 3 | 9.2 | 98 | 1 | fibre | black | 2355.51 | PET |
| 4 | 8.2 | 88 | 1 | fibre | blue | 550.91 | PE |
| 5 | 8.7 | 92 | 1 | fibre | blue | 1689.12 | PA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastorino, P.; Prearo, M.; Di Blasio, A.; Barcelò, D.; Anselmi, S.; Colussi, S.; Alberti, S.; Tedde, G.; Dondo, A.; Ottino, M.; et al. Microplastics Occurrence in the European Common Frog (Rana temporaria) from Cottian Alps (Northwest Italy). Diversity 2022, 14, 66. https://doi.org/10.3390/d14020066
Pastorino P, Prearo M, Di Blasio A, Barcelò D, Anselmi S, Colussi S, Alberti S, Tedde G, Dondo A, Ottino M, et al. Microplastics Occurrence in the European Common Frog (Rana temporaria) from Cottian Alps (Northwest Italy). Diversity. 2022; 14(2):66. https://doi.org/10.3390/d14020066
Chicago/Turabian StylePastorino, Paolo, Marino Prearo, Alessia Di Blasio, Damià Barcelò, Serena Anselmi, Silvia Colussi, Silvia Alberti, Giovanni Tedde, Alessandro Dondo, Michele Ottino, and et al. 2022. "Microplastics Occurrence in the European Common Frog (Rana temporaria) from Cottian Alps (Northwest Italy)" Diversity 14, no. 2: 66. https://doi.org/10.3390/d14020066