

Streamlining Agroecological Management of Invasive Plant Species: The Case of Solanum elaeagnifolium Cav

Abstract
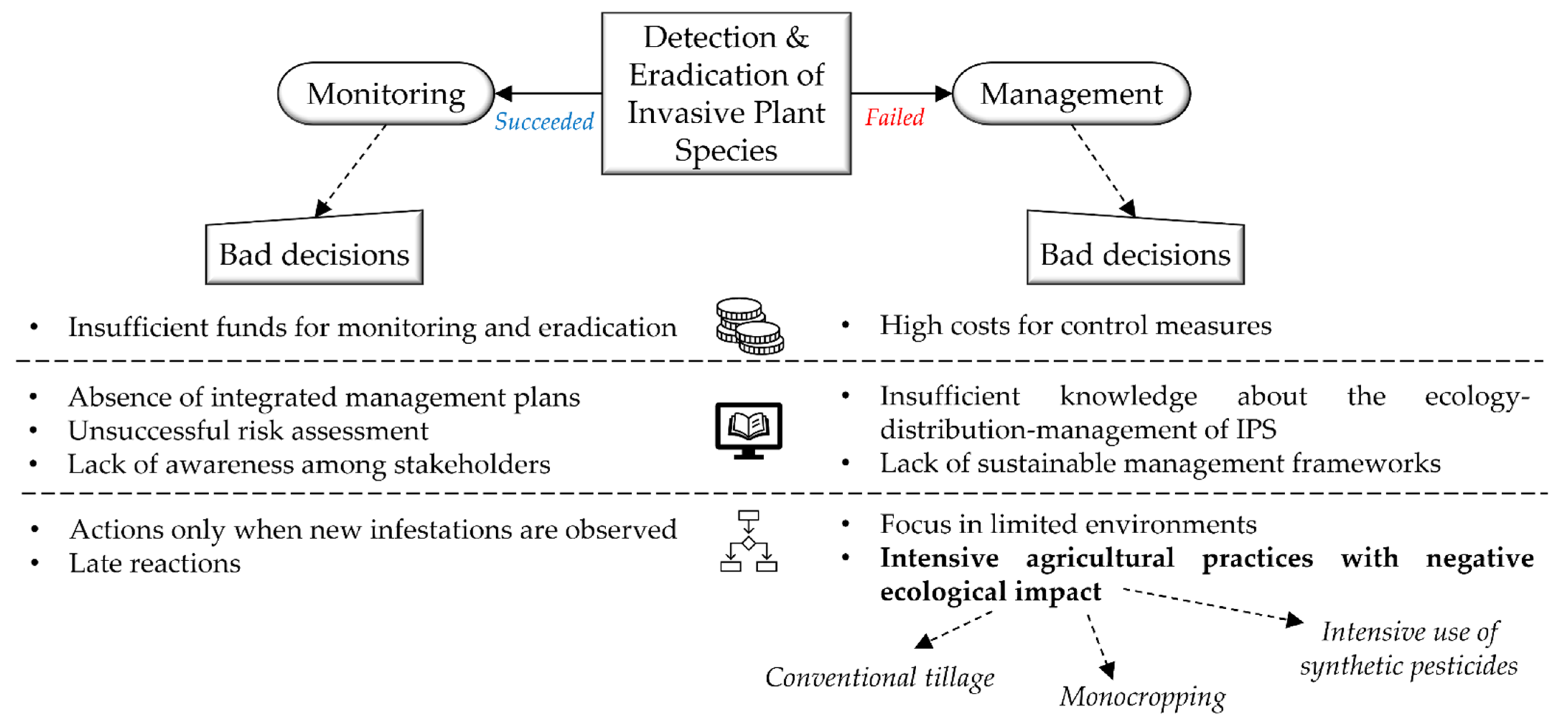
:1. Introduction
2. The Case of Silverleaf Nightshade
2.1. Solanum elaeagnifolium
2.2. Chemical Control
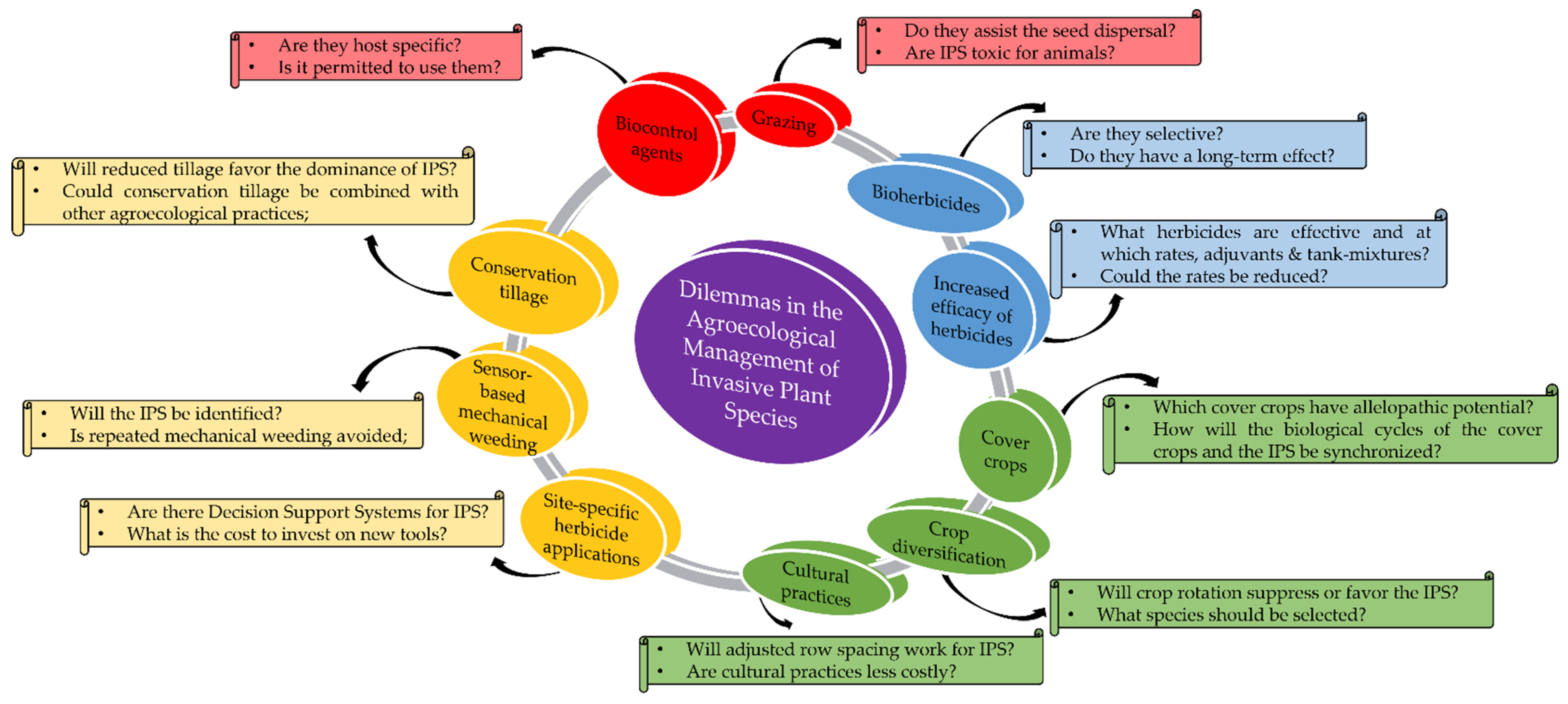
2.3. Agroecological Weed Management Strategies
2.3.1. Bio-Based Herbicides
2.3.2. Cultural and Novel Practices
2.3.3. Precision Agriculture Tools
2.3.4. Mechanical Weeding
2.3.5. Biological Control
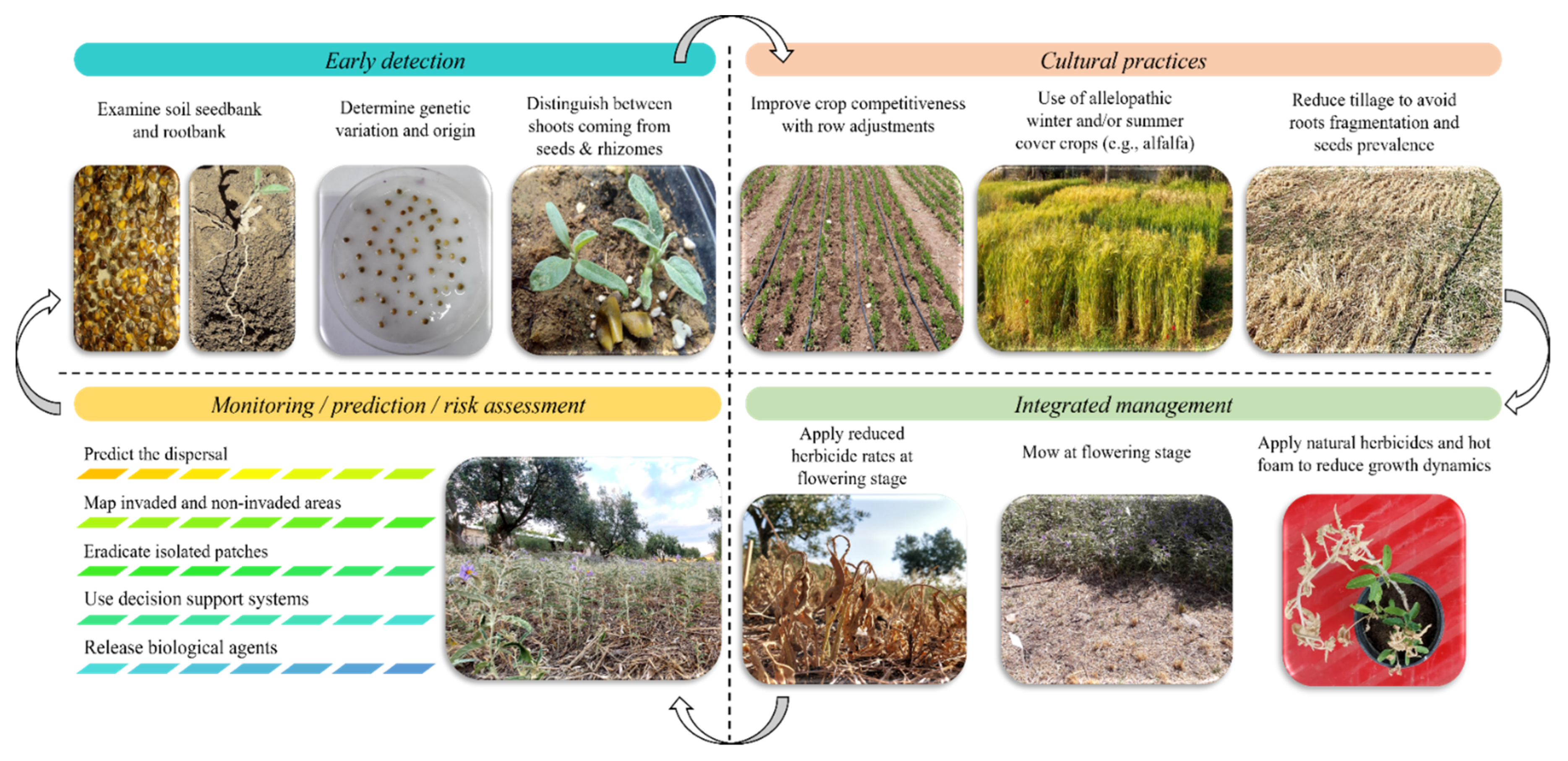
3. A Holistic Framework for Silverleaf Nightshade Management
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Storkey, J.; Mead, A.; Addy, J.; MacDonald, A.J. Agricultural intensification and climate change have increased the threat from weeds. Glob. Chang. Biol. 2021, 27, 2416–2425. [Google Scholar] [CrossRef]
- Tataridas, A.; Jabran, K.; Kanatas, P.; Oliveira, R.S.; Freitas, H.; Travlos, I. Early detection, herbicide resistance screening, and integrated management of Invasive Plant Species: A review. Pest Manag. Sci. 2022, 78, 3957–3972. [Google Scholar] [CrossRef] [PubMed]
- Tataridas, A.; Kanatas, P.; Chatzigeorgiou, A.; Zannopoulos, S.; Travlos, I. Sustainable crop and weed management in the era of the EU Green Deal: A survival guide. Agronomy 2022, 12, 589. [Google Scholar] [CrossRef]
- Cordeau, S. Conservation Agriculture and Agroecological Weed Management. Agronomy 2022, 12, 867. [Google Scholar] [CrossRef]
- EPPO. Solanum elaeagnifolium. EPPO Bull. 2007, 37, 236–245. [CrossRef]
- Uludag, A.; Gbehounou, G.; Kashefi, J.; Bouhache, M.; Bon, M.C.; Bell, C.; Lagopodi, A.L. Review of the current situation for Solanum elaeagnifolium in the Mediterranean Basin. EPPO Bull. 2016, 46, 139–147. [Google Scholar] [CrossRef]
- Tataridas, A.; Oliveira, R.S.; Frazão, L.; Moreira, M.; Travlos, I.; Freitas, H. Solanum elaeagnifolium Cav. (Solanales: Solanaceae) presence confirmed in Portugal. EPPO Bull. 2022, 52, 499–504. [Google Scholar] [CrossRef]
- Viljoen, B.D.; Wassermann, V.D. Suppression of Silver-leaf bitter apple (Solanum elaeagnifolium Cav.) by cultivated pasture crops under dry-land conditions: A preliminary study. S. Afr. J. Plant Soil 2004, 21, 63–66. [Google Scholar] [CrossRef]
- Krigas, N.; Tsiafouli, M.A.; Katsoulis, G.; Votsi, N.E.; van Kleunen, M. Investigating the invasion pattern of the alien plant Solanum elaeagnifolium Cav. (silverleaf nightshade): Environmental and human-induced drivers. Plants 2021, 10, 805. [Google Scholar] [CrossRef]
- Eleftherohorinos, I.G.; Bell, C.E.; Kotoula-Syka, E. Silverleaf nightshade (Solanum elaeagnifolium) control with foliar herbicides. Weed Technol. 1993, 7, 808–811. [Google Scholar] [CrossRef]
- Gitsopoulos, T.K.; Damalas, C.A.; Georgoulas, I. Chemical options for the control of silverleaf nightshade (Solanum elaeagnifolium). Planta Daninha 2017, 35, e017162035. [Google Scholar] [CrossRef] [Green Version]
- Bouhache, M.; Gbẻhounou, G. Current situation of Solanum elaeagnifolium Cav. invasion in Near-East countries (Iraq, Jordan, Lebanon and Syria). Rev. Maroc. Prot. Plantes 2014, 6, 29–36. [Google Scholar]
- Baye, Y. Influence of certain factors on Solanum elaeagnifolium Cav. control by glyphosate and amitrole. EPPO Bull. 2007, 37, 153–155. [Google Scholar] [CrossRef]
- Dotray, P.A.; Keeling, J.W. Silverleaf nightshade (Solanum elaeagnifolium) control in cotton with glyphosate at reduced rates. Tex. J. Agric. Nat. Resour. 1996, 9, 33–41. [Google Scholar]
- Bakacsy, L.; Bagi, I. Survival and regeneration ability of clonal common milkweed (Asclepias syriaca L.) after a single herbicide treatment in natural open sand grasslands. Sci. Rep. 2020, 10, 14222. [Google Scholar] [CrossRef]
- Choudhary, M.; Bordovsky, D.G. Timing of glyphosate application on control of silverleaf nightshade and glyphosate-resistant cotton yield. Weed Technol. 2006, 20, 198–203. [Google Scholar] [CrossRef]
- Tataridas, A.; Zervopoulou, A.; Travlos, I. Efficacy of different glyphosate formulations against the invasive weed Solanum elaeagnifolium Cav. In Proceedings of the Invasive Plants Working Group of EWRS Workshop “Our Path after COVID-19”, Palic, Serbia, 23–24 September 2021. [Google Scholar] [CrossRef]
- Stanton, R.; Wu, H.; Lemerle, D. Integrated management of silverleaf nightshade. In Proceedings of the 23rd Asian-Pacific Weed Science Society Conference, Cairns City, QLD, Australia, 26–29 September 2011; pp. 544–548. [Google Scholar]
- Leys, A.R.; Cuthbertson, E.G. Chemical control of silverleaf nightshade in winter annual pastures. In Proceedings of the Fifth Australian Weeds Conference, Melbourne, VIC, Australia, 29 March–2 April 1976; Volume 49. [Google Scholar]
- Baye, Y.; Ameur, A.; Bouhache, M.; Taleb, A. Strategy for the chemical control of Solanum elaeagnifolium Cav. in Morocco. EPPO Bull. 2007, 37, 145–152. [Google Scholar] [CrossRef]
- Wu, H.; Stanton, R.; Lemerle, D. Herbicidal control of Solanum elaeagnifolium Cav. in Australia. Crop Prot. 2016, 88, 58–64. [Google Scholar] [CrossRef]
- Ensbey, R. Noxious and Environmental Weed Control Handbook, a Guide to Weed Control in Non-Crop, Aquatic and Bushland Situations, 4th ed.; Department of Industry and Investment: Orange, NSW, Australia, 2009; p. 71. [Google Scholar]
- Qasem, J.R. Silverleaf nightshade (Solanum elaeagnifolium) in the Jordan Valley: Field survey and chemical control. J. Hortic. Sci. Biotechnol. 2014, 89, 639–646. [Google Scholar] [CrossRef]
- Catt, M.J. The distribution and control. In of silver-leafed nightshade (Solanum elaeagnifolium) in South Australia. In Proceedings of the 4th Australian Weeds Conference, Hobart, TAS, Australia, 29 November–4 December 1970; pp. 355–356. [Google Scholar]
- Rosales-Robles, E.; Sánchez-de la Cruz, R.; Rodríguez-del Bosque, L.Á. Nozzle type does not change herbicide effectiveness of prosulfuron, bromoxynil and 2, 4-D. Agrociencia 2013, 47, 245–253. [Google Scholar]
- Runge, T.A.; Falk, A.D.; Pawelek, K.A.; Smith, F.S.; Schuster, G.L. Herbicide Efficacy on the Control of Silverleaf Nightshade; Texas A&M University-Kingsville: Kingsville, TX, USA, 2018. [Google Scholar]
- Viljoen, B.D.; Stoltsz, C.W.; Rijst, M.V.D. Preliminary evaluation of soil-residual herbicides for the control of silver-leaf bitter apple (Solanum elaeagnifolium Cav.). S. Afr. J. Plant Soil 2011, 28, 85–89. [Google Scholar] [CrossRef]
- Kidston, J.; Thompson, R.; Johnson, A. Primefact 237 Silverleaf Nightshade; New South Wales Department of Primary Industries: Orange, NSW, Australia, 2007; p. 5. [Google Scholar]
- Grichar, W.J.; Dotray, P.A. Influence of spray tip and spray volume on the efficacy of imazapic and imazethapyr on selected weed species. J. Exp. Agric. Int. 2015, 8, 75–86. [Google Scholar] [CrossRef]
- Umeda, K. Postemergence herbicide control of bindweed and silverleaf nightshade in bermudagrass. In Research Progress Reports; Western Society of Weed Science: Coeur d’Alene, ID, USA, 2017; Available online: http://www.wsweedscience.org/publications/research-reports/ (accessed on 26 September 2022).
- Sleugh, B.S.; Peterson, V.F.; Halstvedt, M.; Whitson, T.D.; Dewey, S.A.; Lym, R.G. Aminopyralid: New efficacy research on noxious and invasive weeds. Proc. West. Soc. Weed Sci. 2009, 62, 27. [Google Scholar]
- Baumann, P.A.; Janak, T.W.; Matocha, M.E.; Castner, E.P. Performance of aminocyclopyrachlor for pernicious weed and brush management in Texas pastures and rangeland. Proc. Weed Sci. Soc. Am. 2012, 318. [Google Scholar]
- Henry, G.M.; Bates, A.N.; McKenney, C.B. Tolerance of evening primrose (Oenothera elata Kunth) transplants to preemergence herbicides. Proc. Weed Sci. Soc. Am. 2009, 49, 212. [Google Scholar]
- Mouemar, A.A. La Morelle jaune (Solanum elaeagnifolium Cav.), une espèce envahissante des cultures cotonnières du Nord de la Syrie. In Invasive Plants in the Mediterranean-Type Regions of the World; Brunel, S., Ed.; Council of European Publishing: Strasbourg, France, 2006; p. 248. [Google Scholar]
- Ameur, A.; Baye, Y.; Bouhache, M.; Taleb, A. Review of control methods against silverleaf nightshade (Solanum elaeagnifolium Cav.) in Morocco. EPPO Bull. 2007, 37, 137–144. [Google Scholar] [CrossRef]
- Travlos, I.; Rapti, E.; Gazoulis, I.; Kanatas, P.; Tataridas, A.; Kakabouki, I.; Papastylianou, P. The herbicidal potential of different pelargonic acid products and essential oils against several important weed species. Agronomy 2020, 10, 1687. [Google Scholar] [CrossRef]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef]
- Samada, L.H.; Tambunan, U.S.F. Biopesticides as promising alternatives to chemical pesticides: A review of their current and future status. OnLine J. Biol. Sci. 2020, 20, 66–76. [Google Scholar] [CrossRef]
- Zhang, J.; An, M.; Wu, H.; Li Liu, D.; Stanton, R. Chemical composition of essential oils of four Eucalyptus species and their phytotoxicity on silverleaf nightshade (Solanum elaeagnifolium Cav.) in Australia. Plant Growth Regul. 2012, 68, 231–237. [Google Scholar] [CrossRef]
- Stanton, R.; Wu, H.; An, M.; Lemerle, D. Home among the gum trees–not necessarily so for silverleaf nightshade. In Proceedings of the 16th Australian Weeds Conference, Cairns, QLD, Australia, 18–22 May W2008; pp. 330–332. [Google Scholar]
- Formozis, G.; Tsakaldimi, M.; Ganatsas, P. Are Mediterranean forest ecosystems under the threat of invasive species Solanum elaeagnifolium? iForest 2021, 14, 236. [Google Scholar] [CrossRef]
- Bouhache, M.; Boulet, C.; Mounir, H. Lutte chimique contre Solanum elaeagnifolium Cav. dans les zones non cultivées. Al Awamia 1993, 83, 139–152. [Google Scholar]
- Davis, C.H.; Smith, T.J.; Hawkins, R.S. Eradication of White Horse Nettle in Southern Arizona; Agricultural Experiment Station, University of Arizona: Tucson, AZ, USA, 1945; Volume 195, pp. 1–14. [Google Scholar]
- Mellado, M.; García, J.E.; Arévalo, J.R.; Pittroff, W. Replacement value of Solanum elaeagnifolium for alfalfa hay offered to growing goats. J. Arid. Environ. 2008, 72, 2034–2039. [Google Scholar] [CrossRef]
- Hawker, V. Controlling silverleaf nightshade (Solanum elaeagnifolium Cav.), a deep-rooted perennial weed, with herbicides and grazing. In Proceedings of the 14th Australian Weeds Conference, Wagga Wagga, NSW, Australia, 6–9 September 2004; p. 267. [Google Scholar]
- Rammou, D.L.; Astaras, C.; Migli, D.; Boutsis, G.; Galanaki, A.; Kominos, T.; Youlatos, D. European ground squirrels at the edge: Current distribution status and anticipated impact of climate on Europe’s southernmost population. Land 2022, 11, 301. [Google Scholar] [CrossRef]
- Food and Agricultural Organization of the United Nations (FAO). Iraq and Syria under Attack from Devastating Alien Weed: Silverleaf Nightshade Takes Root in Lebanon and Jordan Too. Working Sheet 24-05-2011. 2011. Available online: https://www.fao.org/news/story/en/item/75333/icode/ (accessed on 26 September 2022).
- Travlos, I.S.; Gatos, A.; Kanatas, P.J. Interference between silverleaf nightshade (Solanum elaeagnifolium Cav.) and alfalfa (Medicago sativa L.) cultivars. Hell. Plant Prot. J. 2013, 6, 41–48. [Google Scholar]
- Ford, A.J.; Dotray, P.A.; Keeling, J.W.; Wilkerson, J.B.; Wilcut, J.W.; Gilbert, L.V. Site-specific weed management in cotton using WebHADSS™. Weed Technol. 2011, 25, 107–112. [Google Scholar] [CrossRef]
- Boyd, J.W.; Murray, D.S.; Tyrl, R.J. Silverleaf nightshade, Solanum elaeagnifolium, origin, distribution, and relation to man. Econ. Bot. 1984, 38, 210–217. [Google Scholar] [CrossRef]
- Olckers, T.; Zimmermann, H.G. Biological control of silverleaf nightshade, Solanum elaegnifolium, and bugweed, Solanum mauritianum, (Solanaceae) in South Africa. Agric. Ecosyst Environ. 1991, 37, 137–155. [Google Scholar] [CrossRef]
- Stanton, R.; Wu, H.; Lemerle, D. Root regenerative ability of silverleaf nightshade (‘Solanum elaeagnifolium’Cav.) in the glasshouse. Plant Prot. Q. 2011, 26, 54–56. [Google Scholar]
- Chavana, J.; Singh, S.; Vazquez, A.; Christoffersen, B.; Racelis, A.; Kariyat, R.R. Local adaptation to continuous mowing makes the noxious weed Solanum elaeagnifolium a superweed candidate by improving fitness and defense traits. Sci. Rep. 2021, 11, 6634. [Google Scholar] [CrossRef]
- Wapshere, A.H. Prospects for the biological control of silver-leaf nightshade, Solanum elaeagnifolium, in Australia. In Proceedings of the Eighth Australian Weeds Conference, Sydney, NSW, Australia, 21–25 September 1987; p. 104. [Google Scholar]
- Lefoe, G.; Haegi, L.; Rumpff, L.; Gopurenko, D.; Slater, A.T.; Butler, K.; Hauser, C.E. Assessing the fundamental host-range of Leptinotarsa texana Schaeffer as an essential precursor to biological control risk analysis. Biol. Con. 2020, 143, 104165. [Google Scholar] [CrossRef]
- Olckers, T.; Hoffmann, J.H.; Moran, V.C.; Impson, F.A.C.; Hill, M.P. The initiation of biological control programmes against Solanum elaeagnifolium Cavanilles and S. sisymbriifolium Lamarck (Solanaceae) in South Africa. Afr. Entomol. Mem. 1999, 1, 55–63. [Google Scholar]
- Field, R.P.; Kwong, R.M.; Sagliocco, J.L. Host specificity of Ditylenchus phyllobius, a potential biological control agent of silver-leaf nightshade (Solanum elaeagnifolium Cav.) in Australia. Plant Prot. Q. 2009, 24, 141–145. [Google Scholar]
- Paynter, Q.; Hill, R.; Bellgard, S.; Dawson, M. Improving Targeting of Weed Biological Control Projects in Australia; Land and Water Australia: Canberra, Australia, 2009. [Google Scholar]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: A review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Hill, M.P.; Hulley, P.E.; Olckers, T. Insect herbivores on the exotic weeds Solanum elaeagnifolium Cavanilles and S. sisymbriifolium Lamarck (Solanaceae) in South Africa. Afr. Entomol. 1993, 1, 175–182. [Google Scholar]
- Hoffmann, J.H.; Moran, V.C.; Impson, F.A.C. Promising results from the first biological control programme against a solanaceous weed (Solanum elaeagnifolium). Agric. Ecosyst. Environ. 1998, 70, 145–150. [Google Scholar] [CrossRef]
- Klein, H. The satansbos leaf beetle. Biological Control of Solanum Weeds (Solanaceae Potato Family). Pretoria, ARC-PPRI 2007.
- Pitso, D.C. A Biophysical and Economic Evaluation of Biological and Chemical Control Methods for Solanum elaegnifolium (Silverleaf Nightshade) in the Limpopo Province, South Africa. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2010. [Google Scholar]
- Senaratne, K.A.D.W.; Palmer, W.A.; Sutherst, R.W. Applications of CLIMEX modelling leading to improved biological control. In Proceedings of the 16th Australian Weeds Conference, Cairns, QLD, Australia, 18–22 May 2008; Queensland Weeds Society: Brisbane, QLD, Australia, 2008; pp. 234–236. [Google Scholar]
- Adamski, D.; Brown, J.W. A new species of Frumenta Busck (Lepidoptera: Gelechiidae: Gnorimoschemini) from México: A potential biocontrol agent against Solanum elaeagnifolium (Solanaceae). Proc. Entomol. Soc. Wash. 2022, 104, 1029–1035. [Google Scholar]
- Olckers, T.; Hulley, P.E. Resolving ambiguous results of host-specificity tests: The case of two Leptinotarsa species (Coleoptera: Chrysomelidae) for biological control of Solanum elaeagnifolium (Solanaceae) in South Africa. Afric. Entomol. 1994, 2, 137–144. [Google Scholar]
- SPHDS. National Diagnostic Protocol for Colorado Potato Beetle, Leptinotarsa decemlineata; Australian Government, Department of Agriculture: Canberra, Australia, 2018.
- Kwong, R.M.; Sagliocco, J.L.; Weiss, J.; Hunt, T.; Morfe, T. Feasibility of biological control of silverleaf nightshade in Australia. In Proceedings of the 16th Australian Weeds Conference, Cairns, QLD, Australia, 18–22 May 2008; Cairns Convention Centre, North Queensland: Cairns, QLD, Australia, 2008; pp. 18–22. [Google Scholar]
- Sforza, R.E.N.E.; Jones, W.A. Potential for classical biocontrol of silverleaf nightshade in the Mediterranean Basin. EPPO Bull. 2007, 37, 156–162. [Google Scholar] [CrossRef]
- Skinner, J.A.; Orr, C.C.; Robinson, A.F. Histopathogenesis of the galls induced by Nothanguina phyllobia in Solanum elaeagnifolium. J. Nematol. 1980, 12, 141. [Google Scholar]
- Parker, P.E. Nematode control of silverleaf nightshade (Solanum elaeagnifolium); a biological control pilot project. Weed Sci. 1986, 34, 33–34. [Google Scholar] [CrossRef]
- Northam, F.E.; Orr, C.C. Effects of a Nematode on Biomass and Density of Silverleaf Nightshade. J. Range Manag. 1982, 35, 536–537. [Google Scholar] [CrossRef]
- Robinson, A.F.; Orr, C.C.; Abernathy, J.R. Distribution of Nothanguina phyllobia and its potential as a biological control agent for silver-leaf nightshade. J. Nematol. 1978, 10, 362. [Google Scholar]
- Gómez, E.Μ.; Ramírez Suárez, A.; Cuevas Ojeda, J.; Martínez Gómez, D. Identification and phylogenetic analysis of the leaf-galling nematode Orrina phyllobia affecting Solanum elaeagnifolium Cav. in Guanajuato, Mexico. Rev. Mex. Fitopatol. 2016, 34, 184–199. [Google Scholar]
- Scott, M.B. A lost range study of Orrina phyllobia. a potential biocontrol agent of satansbos (Solanum elaeagnifolium) in South Africa. Phytophylactica 1991, 23, 315. [Google Scholar]
- Kariyat, R.R.; Chavana, J. Field data on plant growth and insect damage on the noxious weed Solanum eleaegnifolium in an unexplored native range. Data Brief 2018, 19, 2348–2351. [Google Scholar] [CrossRef] [PubMed]
- Medal, J.C.; Pitelli, R.A.; Santana, A.; Gandolfo, D.; Gravena, R.; Habeck, D.H. Host specificity of Metriona elatior, a potential biological control agent of tropical soda apple, Solanum viarum, in the USA. BioControl 1999, 44, 421–436. [Google Scholar] [CrossRef]
- Olckers, T. Biology and host range of Platyphora semiviridis, a leaf beetle evaluated as a potential biological control agent for Solanum mauritianum in South Africa. BioControl 1998, 43, 225–239. [Google Scholar] [CrossRef]
- Goeden, R.D. Insect ecology of silverleaf nightshade. Weed Sci. 1971, 19, 45–51. [Google Scholar] [CrossRef]
- Adjim, Z.; Kazi Tani, C. L’infestation par Solanum elaeagnifolium menace l’Algérie. Rev. D’écologie (Terre et Vie) 2018, 73, 569–581. [Google Scholar] [CrossRef]
- Dellow, J.J. Distribution of silverleaf nightshade (Solanum elaeagnifolium) in the sheep/wheat belt of New South Wales. In Proceedings of the 10th Australian Weeds Conference/14th Asian-Pacific Weed Science Society, Brisbane, QLD, Australia, 6–10 September 1993; p. 131. [Google Scholar]
- Boyd, J.W.; Murray, D.S. Effects of shade on silverleaf nightshade (Solanum elaeagnifolium). Weed Sci. 1982, 30, 264–269. [Google Scholar] [CrossRef]
- Tataridas, A.; Kanatas, P.; Travlos, I. Effect of a winter cover crop on maize production and management of the invasive weed Solanum elaeagnifolium Cav. In Proceedings of the Frontiers of Science and Technology in Crop Breeding and Production, Maize Research Institute, Zemun, Serbia, 8–9 June 2021. [Google Scholar] [CrossRef]
- Tataridas, A.; Smyri, E.; Kanatas, P.; Travlos, I. Efficacy of post-emergence herbicides against Solanum elaeagnifolium Cav.: A major threat for soybean production. In Proceedings of the Frontiers of Science and Technology in Crop Breeding and Production, Maize Research Institute, Zemun, Serbia, 8–9 June 2021. [Google Scholar] [CrossRef]
- Kriticos, D.; Crossman, N.D.; Ota, N.; Scott, J.K. Climate Change and Invasive Plants in South Australia; CSIRO Climate Adaptation Flagship: Canberra, Australia, 2010; pp. 86–92. [Google Scholar]
- Kanatas, P.; Travlos, I.S.; Gazoulis, I.; Tataridas, A.; Tsekoura, A.; Antonopoulos, N. Benefits and limitations of decision support systems (DSS) with a special emphasis on weeds. Agronomy 2020, 10, 548. [Google Scholar] [CrossRef]
- Kanatas, P.; Travlos, I.; Tataridas, A.; Gazoulis, I. Decision-Making and Decision Support System for a Successful Weed Management. In Information and Communication Technologies for Agriculture—Theme III: Decision; Springer: Cham, Switzerland, 2020; pp. 159–179. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Habitat | Method | S. elaeagnifolium Growth Stage | Season | Targets |
|---|---|---|---|---|
| Field crops (maize, cotton, soybean, peanut, vegetables) |
| Dormancy Seedling | Early in spring |
|
| Perennial crops (olive groves, citrus orchards, vineyards) |
| Before flowering or during flowering | Late spring and early to mid-summer |
|
| Roadsides (highways and road network) |
| Before flowering or during flowering for mowing Seedling to flowering for defoliators biological agents | Early spring to late summer |
|
| Grasslands |
| Seedling to flowering | Early spring to mid-summer |
|
| Urban areas (parks, flower beds, roadsides) |
| Seedling to berry | Early spring to autumn |
|
| Abandoned places |
| Seedling to berry | Early spring to autumn |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tataridas, A.; Kanatas, P.; Travlos, I. Streamlining Agroecological Management of Invasive Plant Species: The Case of Solanum elaeagnifolium Cav. Diversity 2022, 14, 1101. https://doi.org/10.3390/d14121101
Tataridas A, Kanatas P, Travlos I. Streamlining Agroecological Management of Invasive Plant Species: The Case of Solanum elaeagnifolium Cav. Diversity. 2022; 14(12):1101. https://doi.org/10.3390/d14121101
Chicago/Turabian StyleTataridas, Alexandros, Panagiotis Kanatas, and Ilias Travlos. 2022. "Streamlining Agroecological Management of Invasive Plant Species: The Case of Solanum elaeagnifolium Cav" Diversity 14, no. 12: 1101. https://doi.org/10.3390/d14121101