Family Pterygosomatidae Oudemans, 1910.
3.2.1. Description
Female (holotype, range for nine paratypes).
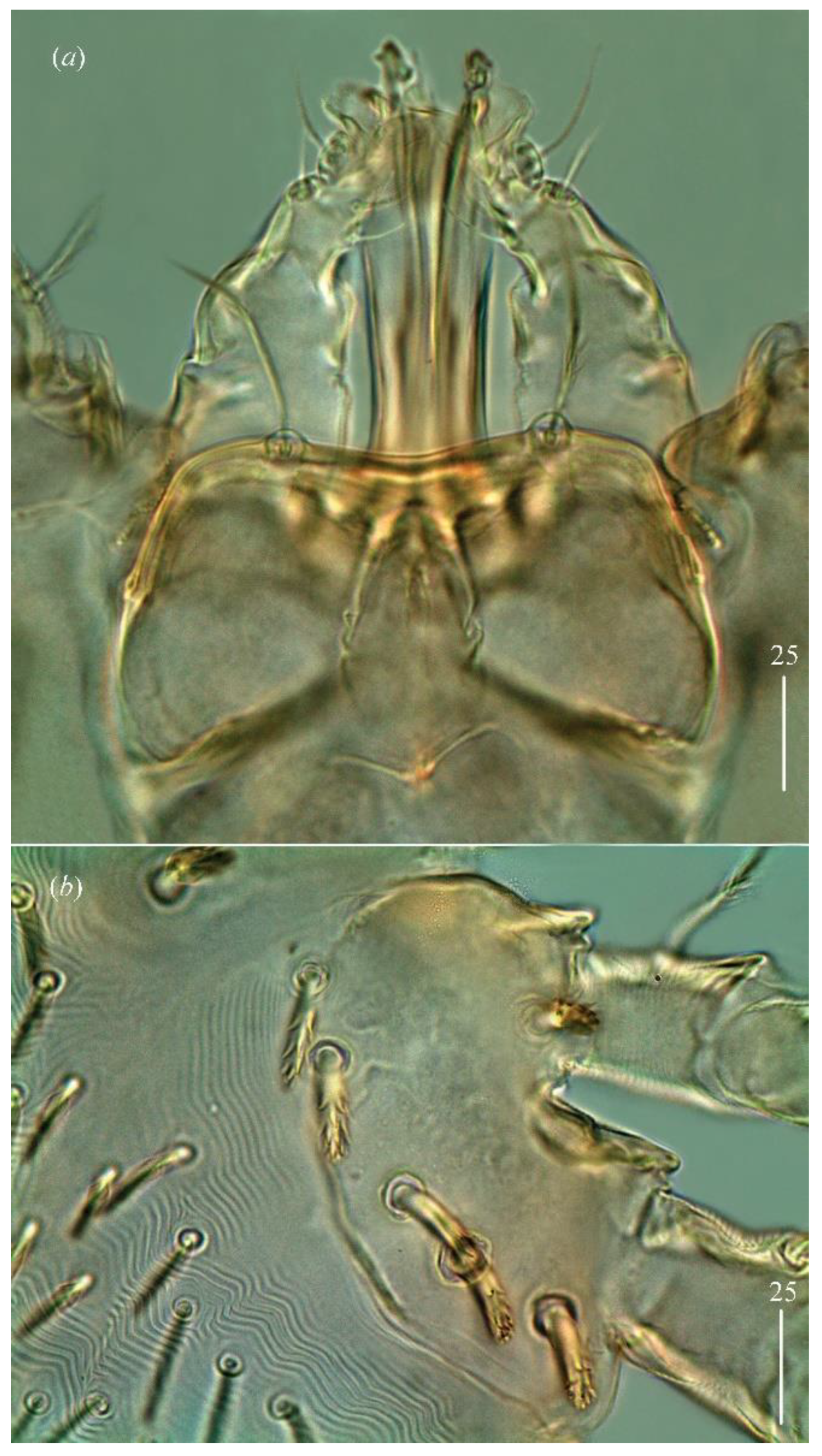
Gnathosoma. Chelicerae 85 (80–95) long. Swollen, proximal part of cheliceral base 45 (30–40) long and slender distal part 45 (45–55) long. Movable cheliceral digit three-pronged while fixed cheliceral digit spinous and approximately 5 (5–10) long. Palpal femur with thick plumose seta
dF 15 (15–20) long; palpal genu with filiform smooth seta
dG, 65 (50–60) long. Palpal tibia with three smooth setae (
dTi,
l′Ti and
l″Ti) and slender curved claw. Palpal tarsi with four smooth setae. Subcapitular seta
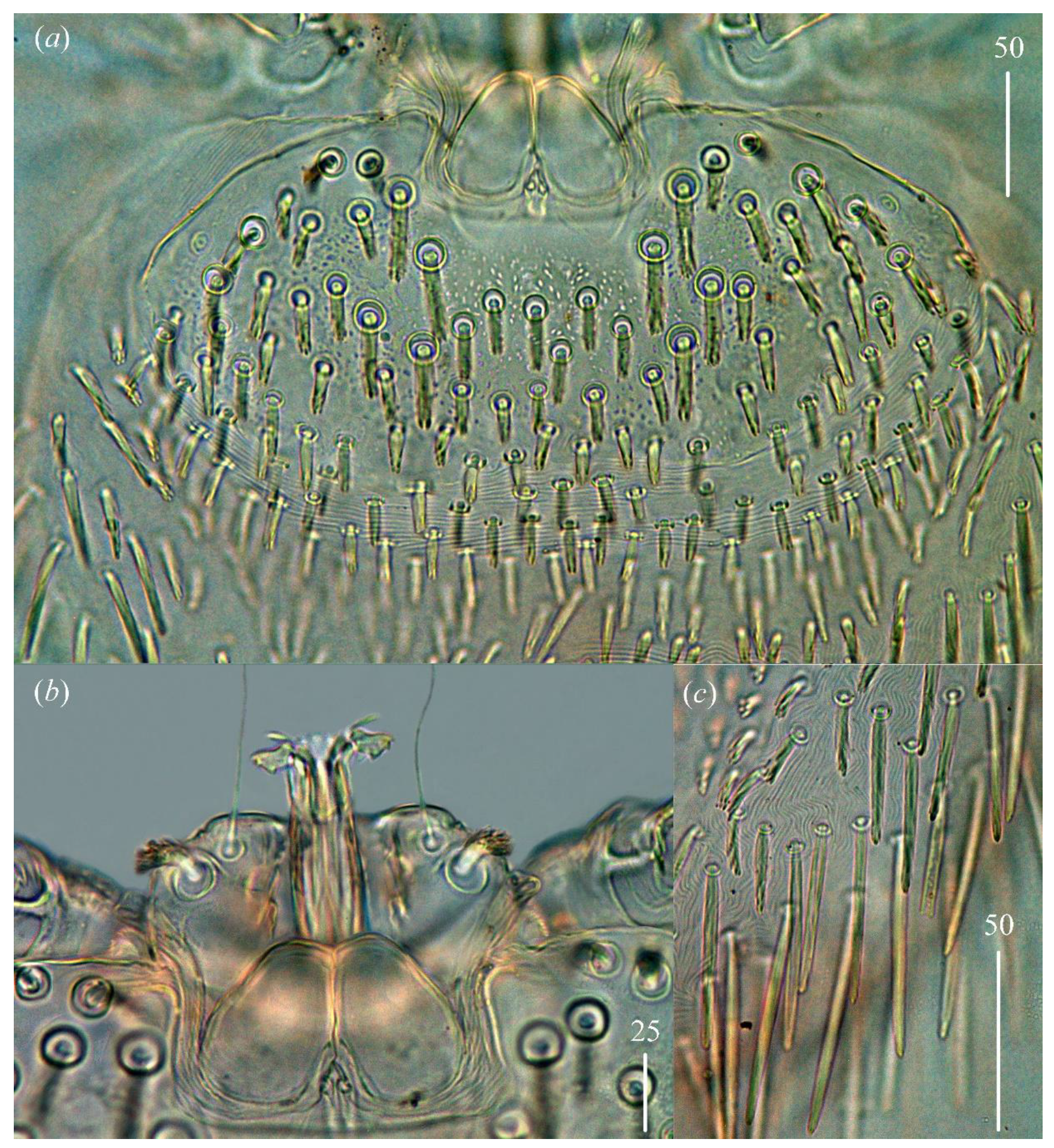
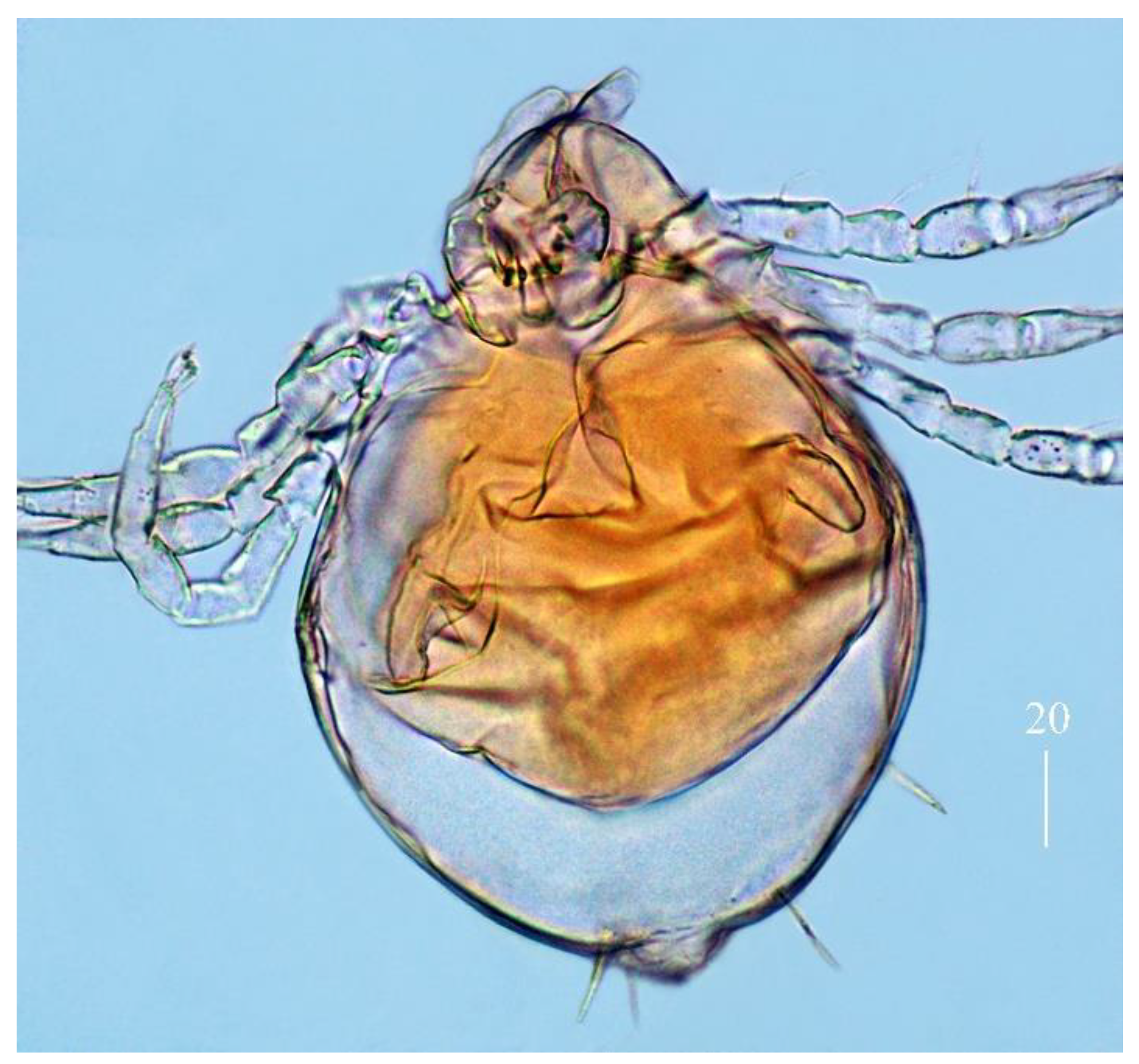
n filiform and smooth, about 45 (40–50) long. Each branch of peritremes with barely visible chambers 75 (70–85) long. Hypostome with ornamented apex (
Figure 2b).
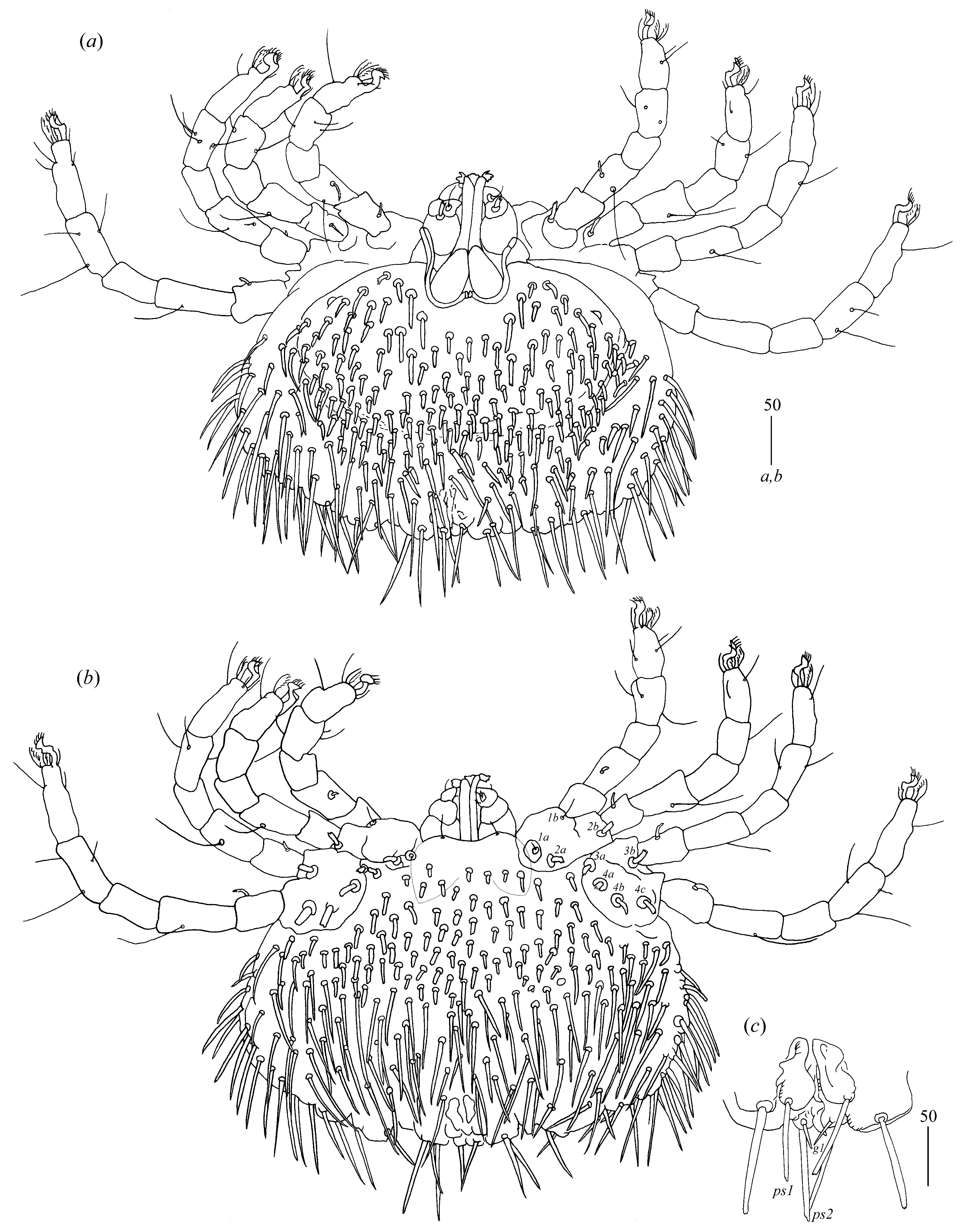
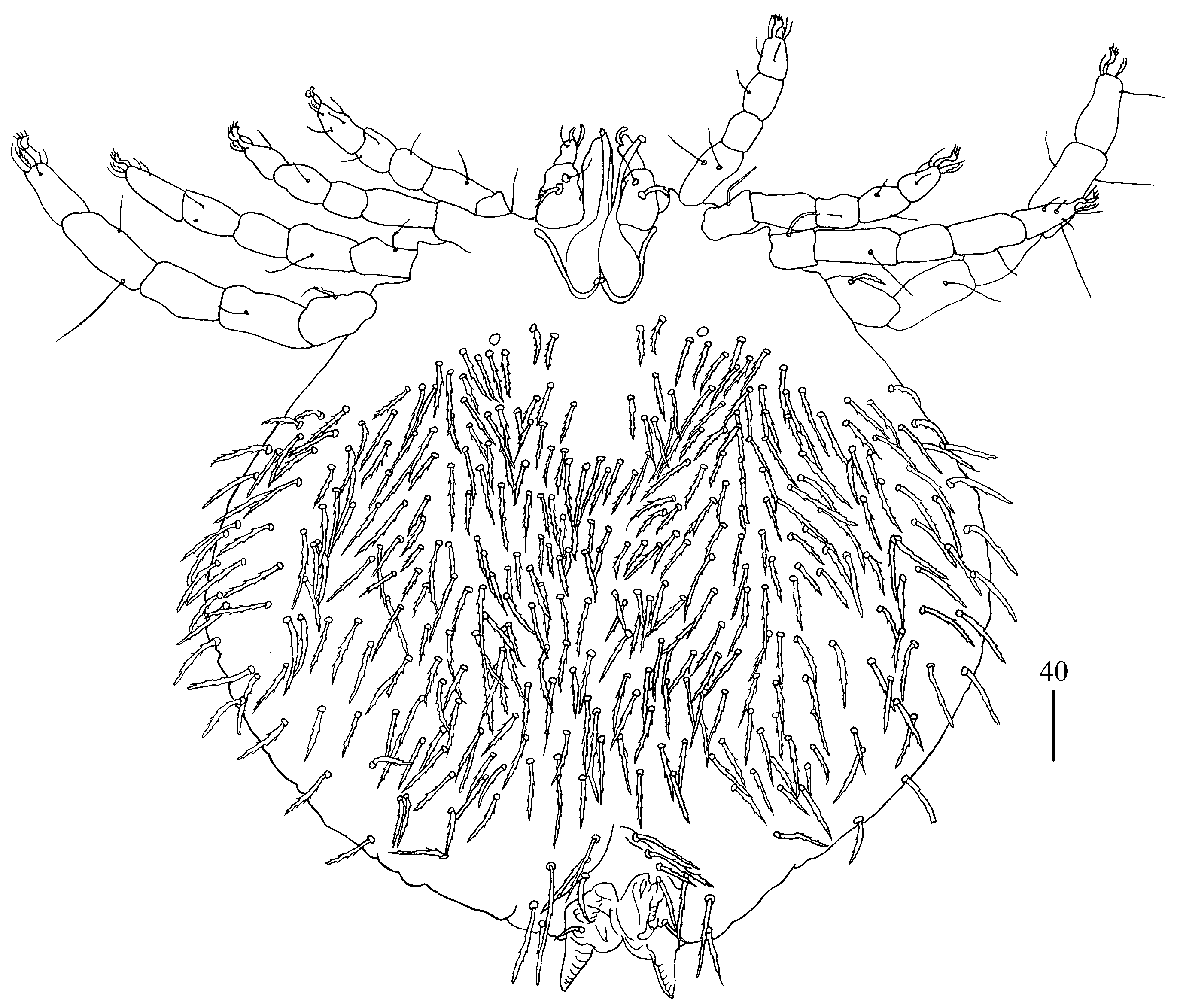
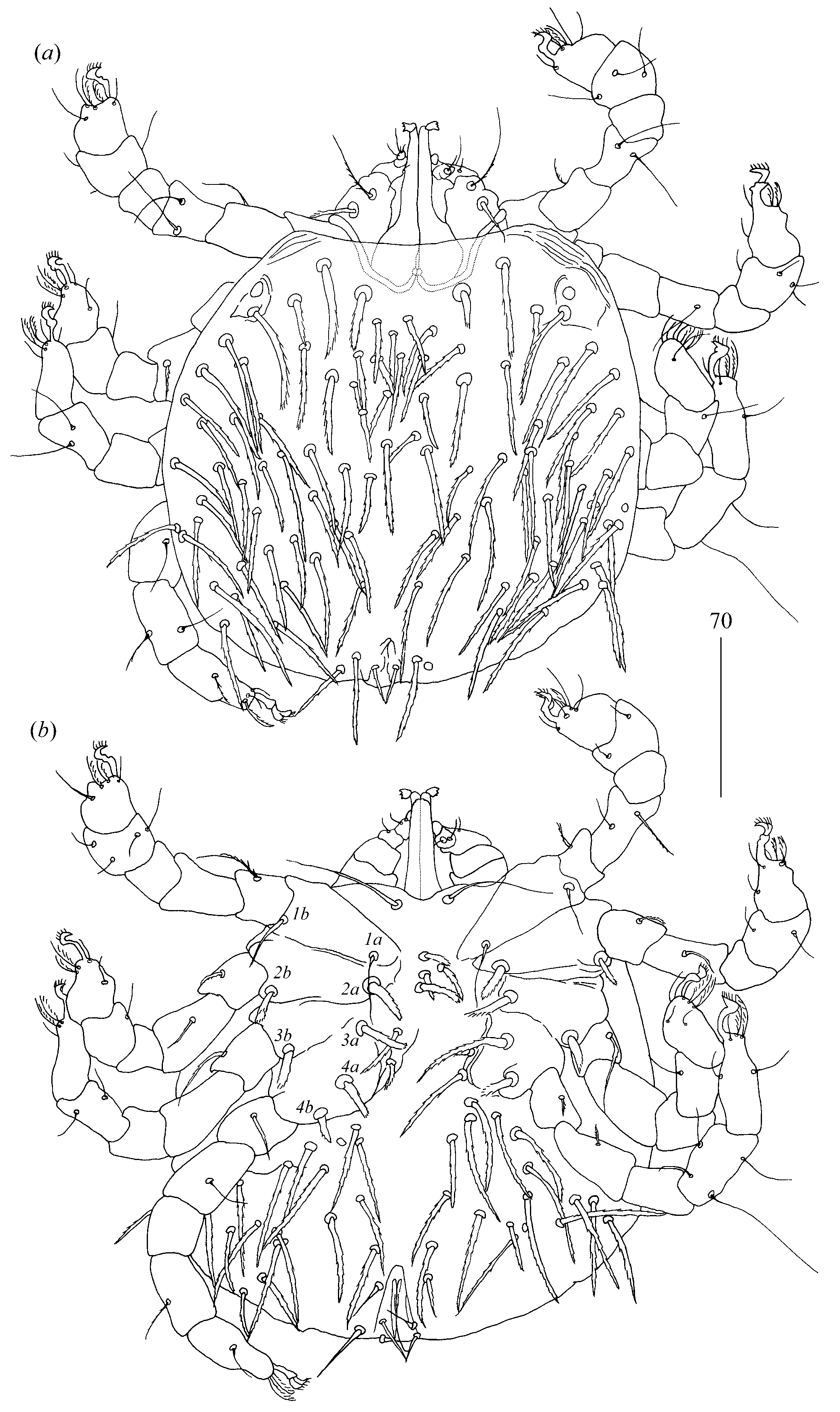
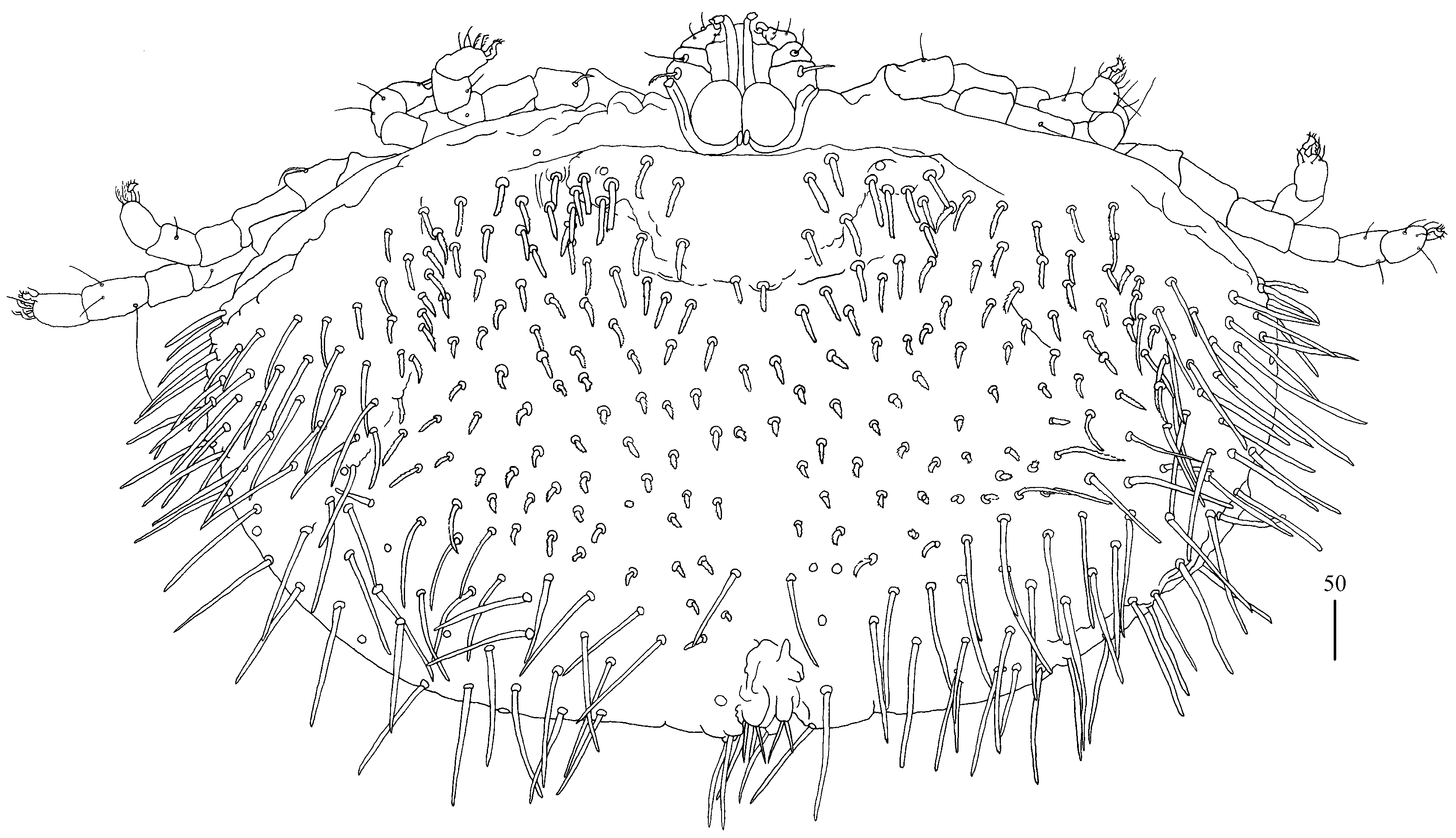
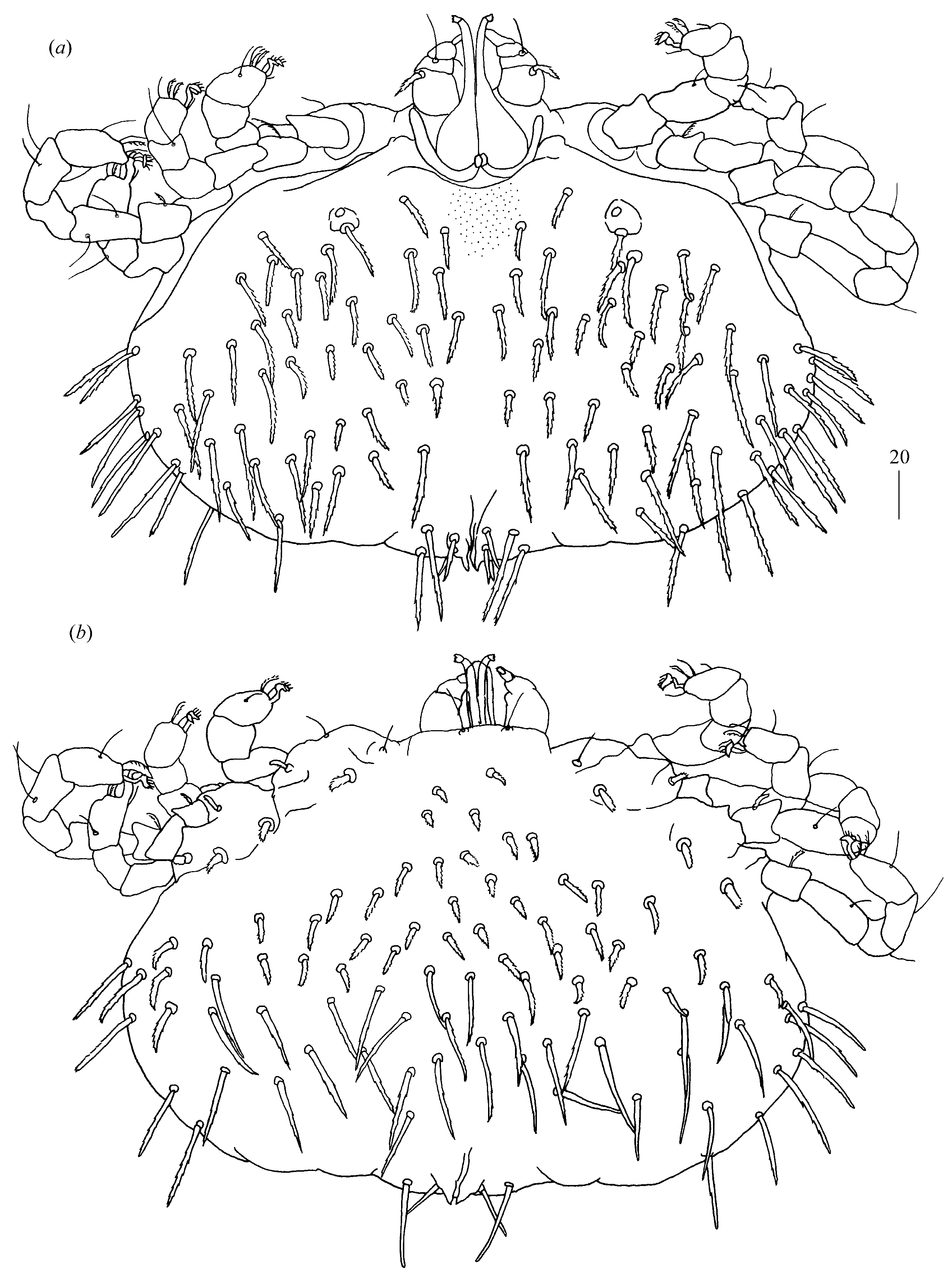
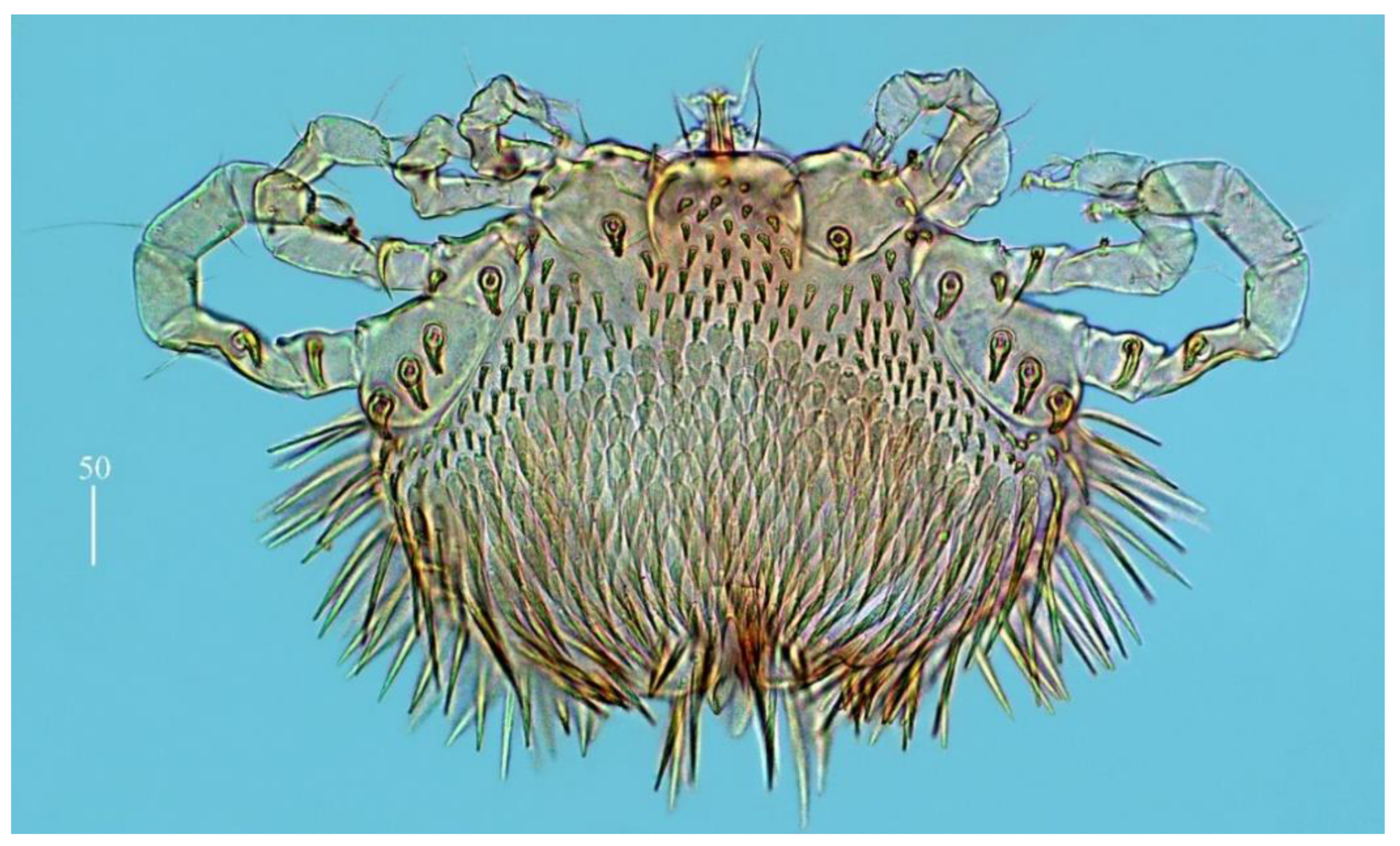
Idiosoma 210 (155–260) long and 305 (295–385) wide. Dorsum (
Figure 1). Propodonotal shield well outlined, 110 (85–115) long and 230 (220–270) wide, covered by minute punctations (
Figure 2a). On propodonotal shield, small eyes situated on lateral margins present and 73 (73–81) stout and plumose setae, 10–20 long. These setae decrease in length from anterior to posterior part of propodonotal shield. Posterior to propodonotal shield, four rows of numerous stout and plumose setae (about 15) long present. These setae increase in length from anterior to posterior part of idiosoma (10–15 long) and resemble setae situated posteriorly on propodonotal shield. Postero-lateral and most posterior part of idiosoma with numerous flattened longer setae (20–50 long) that increase in length from anterior to posterior part of idiosoma; these setae are slightly serrate only at tip (as in
Figure 2c). Venter (
Figure 1b). Anterior part with numerous thick and plumose setae (10–15 long) and posterior part with about 60 pairs of thick and flattened setae (40–50 long). Genital region (
Figure 1c). Genital setae represented by four pairs of slender blunt pointed setae
g1–
g4. Setae
g1 and
g2 about 20 long,
g3 about 10 long and
g4 about 30 long. Setae
g1–
g3 situated medially on genital valves and setae
g4 situated laterally. Pseudanal series represented by 11 pairs of blunt-pointed smooth and flattened setae
ps1–
ps11, about 50 (40–55) long.
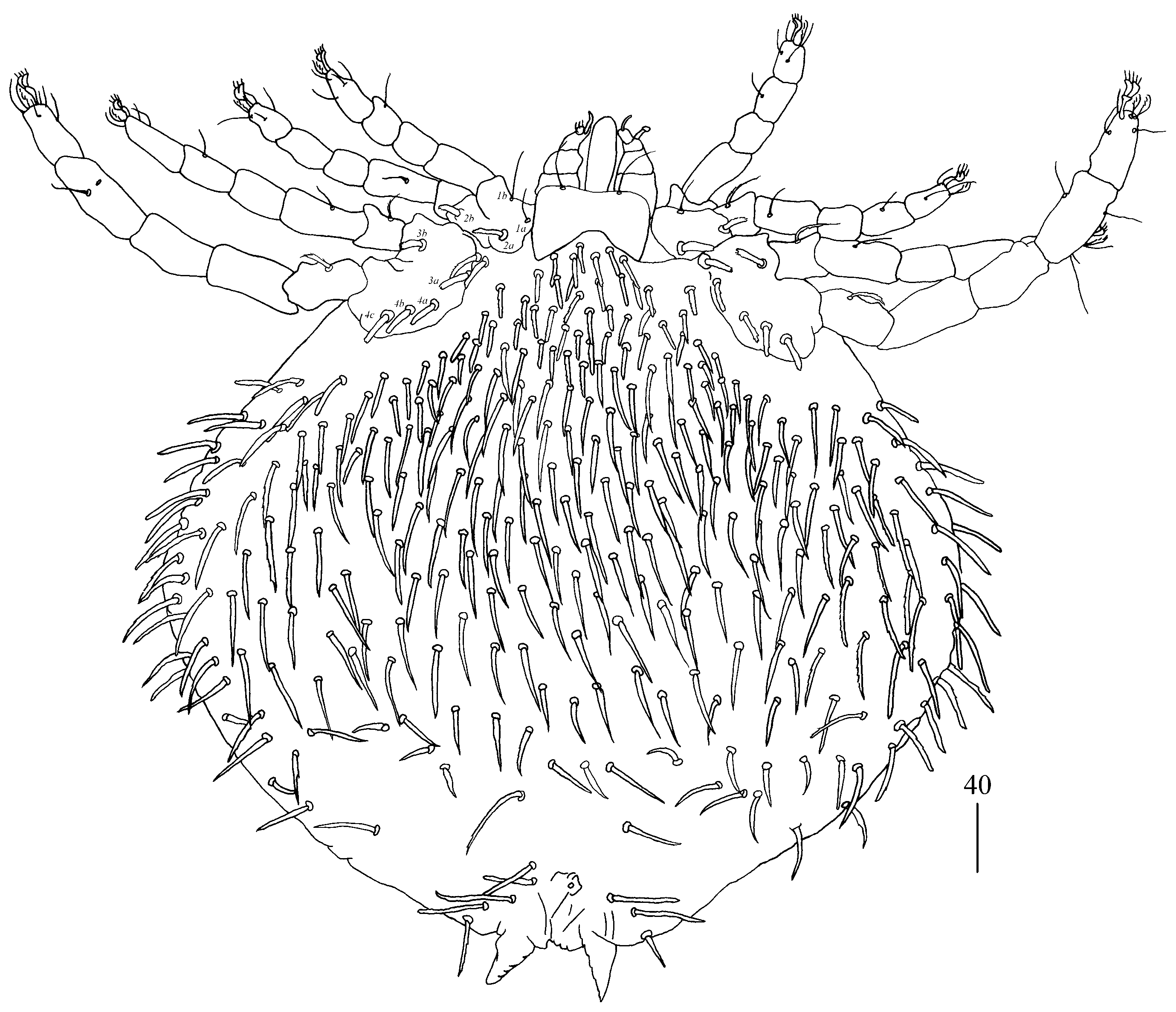
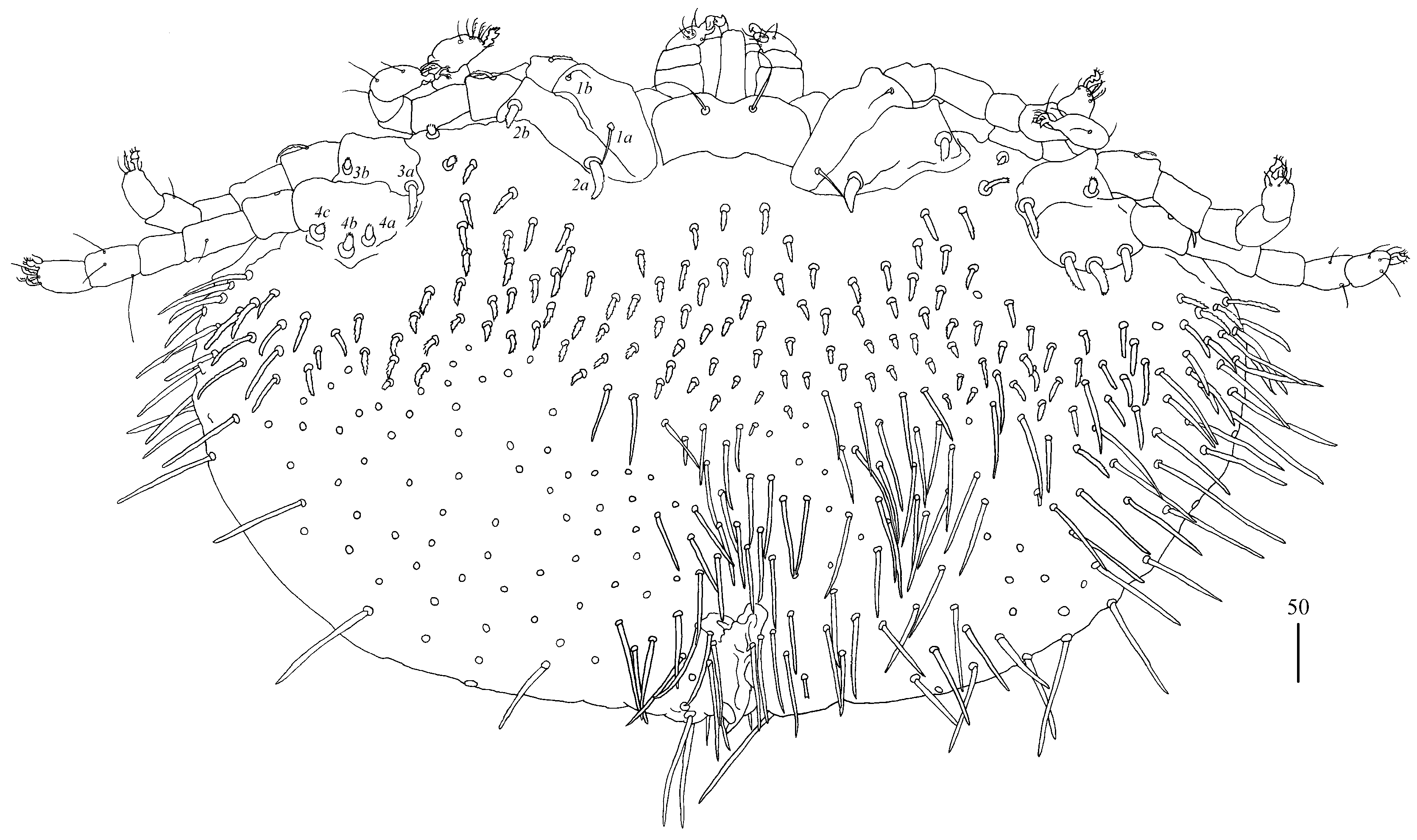
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b,
4a,
4b and
4c arranged in formula: 2–2–2–3. All coxal setae thick and plumose, except for filiform and smooth setae
1a and
1b. Two plumose setae present between coxal plates I and II. Leg chaetotaxy as follows: tibiae I–IV (5–5[4]–4–5), genua I–IV (1–0–0–1), femora I–IV (3–2–1–2) and trochanters I–IV (1–1–1–2). Setae
d′TiI–
IV,
d″TiI,
d′TiIV,
v′TiI–
IV,
v″TiI–
IV,
l′TiI–
IV,
lGI,
lGIV,
dlFI–
IV and
dFIV filiform and smooth; setae
d″FI,
vFI–
II,
vTrI–IV and
v″TrIV filiform and serrate. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Solenidion
ω1 (about 25 long) longer than seta
ft (about 5 long). Setae
tc′,
tc″,
it′ and
it″ of leg I represented by euphatidia;
tc′ and
tc″ of legs II–IV,
u′,
u″,
vs′,
vs″,
a′,
a″ and
pl′ of legs I–IV filiform.
MALE (range for two paratypes).
Gnathosoma as in female. Chelicerae 80 long; swollen proximal part and slender distal part subequal in length, about 40 long. Fixed cheliceral digit about 5 long. Setae
dF and
dG 15 and 45 long, respectively. Subcapitular setae
n 35 long. Each branch of peritremes 50 long.
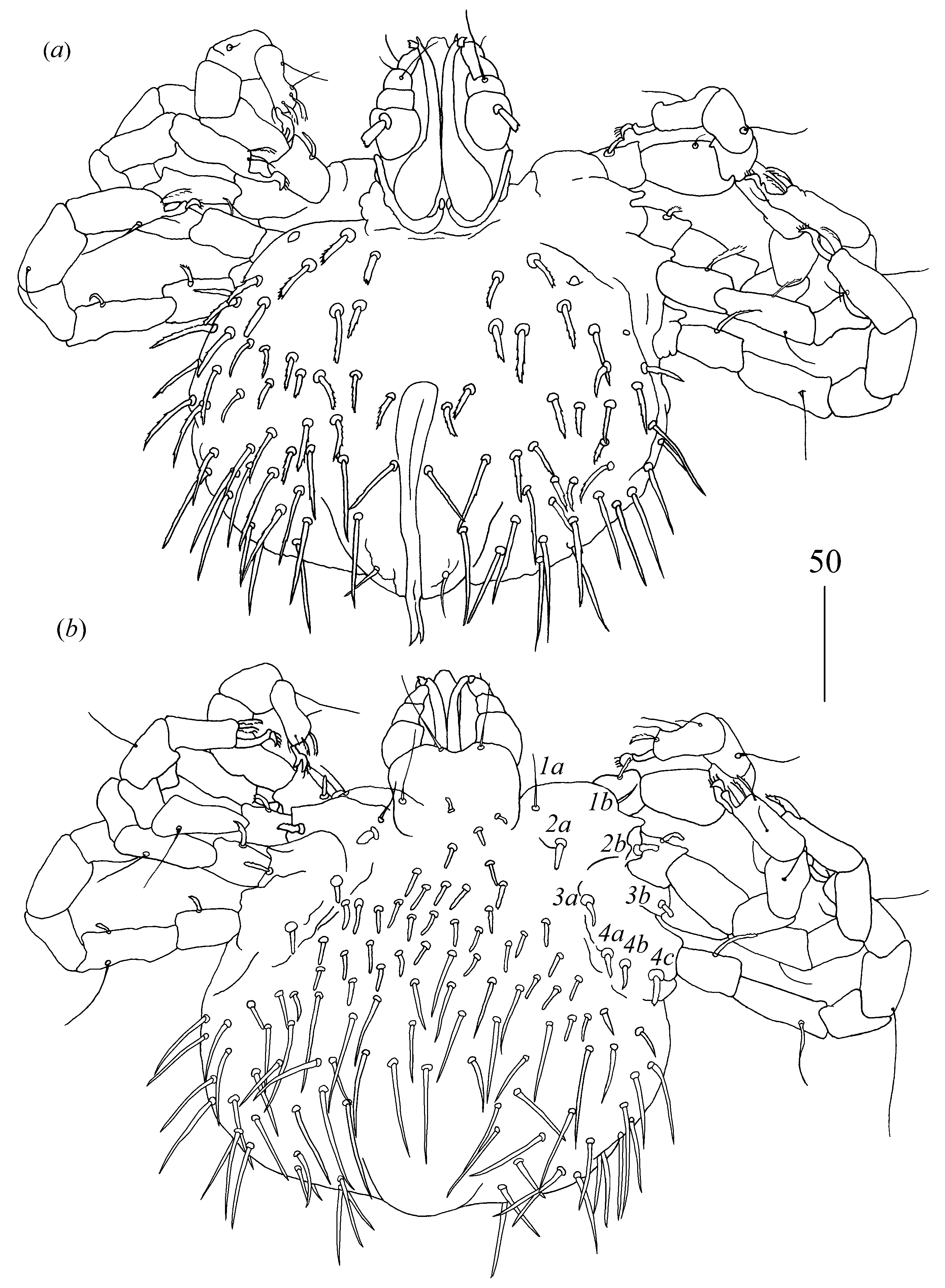
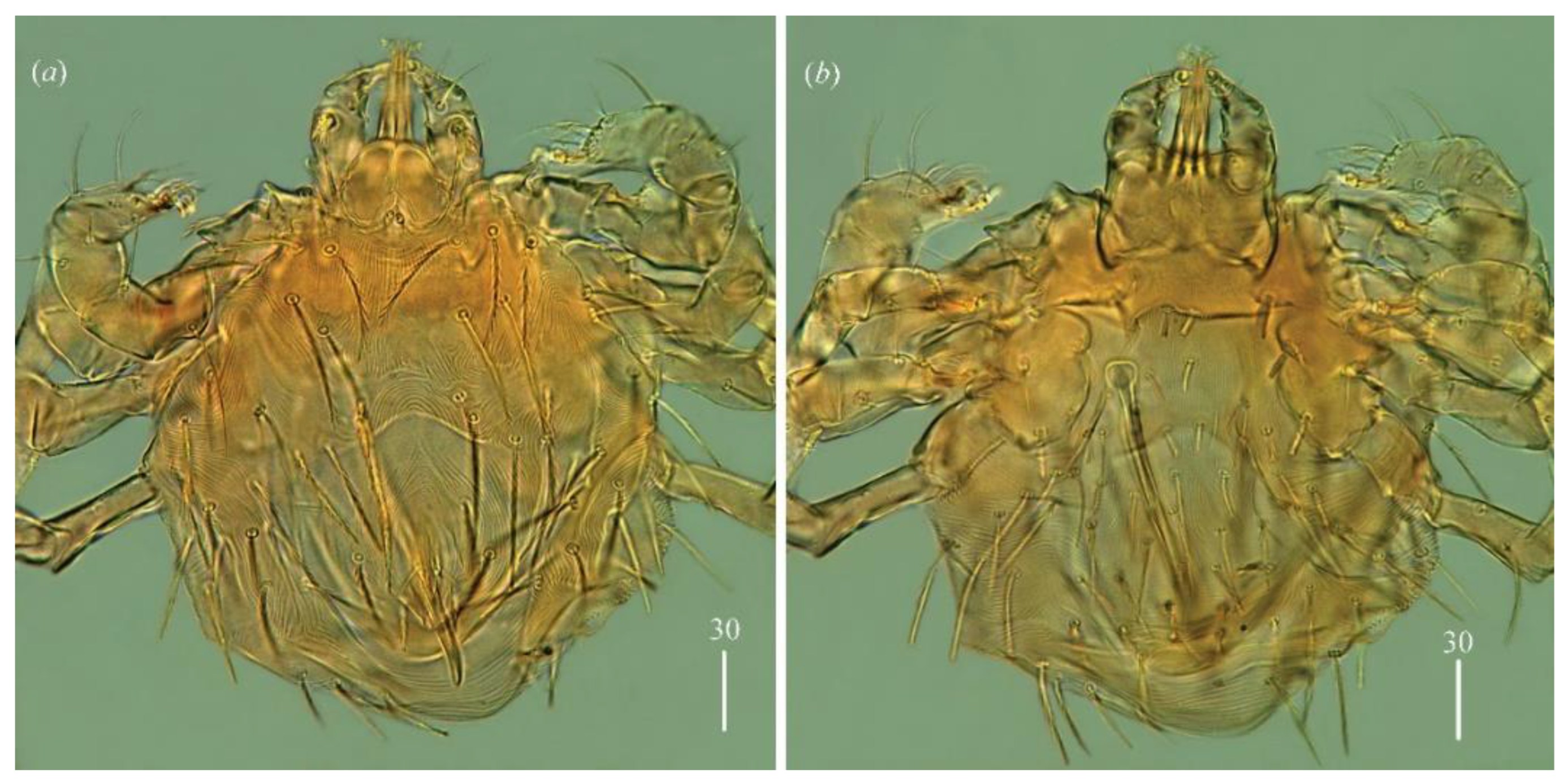
Idiosoma 150–155 long and 145–180 wide. Dorsum (
Figure 3a). Propodonotal shield reniform, 45–55 long and 70 wide, with small eyes present on lateral margins and six plumose and thick setae: two pairs (15 long) present antero-laterally and four pairs (10–20 long) present postero-laterally. Posterior to propodonotal shield, 21–23 pairs of slightly plumose setae (10–40 long) present. These setae increase in length from anterior to posterior part of idiosoma. Venter (
Figure 3b) with 18 short and plumose setae in antero-medial part, 5–10 long, and 29 thick and smooth setae in postero-lateral part, 20–40 long. Aedeagus 90 long, ended with spine-like structure. Most posteriorly situated one pair of spine-like setae about 10 long. Ano-genital opening covered with two curved spines, about 5 long. Laterally to aedeagus one pair of slightly serrate setae, 20 long, present.
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b and
4a arranged in formula: 2–2–2–1. All coxal setae thick and plumose, except for filiform and smooth setae
1a and
1b. Leg chaetotaxy: tibiae I–IV (5–5–5[4]–5), genua I–IV (1–0–1–1), femora I–IV (3–1–1–2) and trochanters I–IV (1–1–1–1). Setae
d’TiI–
IV,
d”TiI–IV,
lTiI–IV,
v’TiI–
IV,
v”TiI–IV,
l’GI,
lGIII–IV,
vFI,
l’FI and
vFIV filiform and smooth; setae
l”FI with barely discernible serration; setae
l”FII–FIV slightly serrate and setae
lTrI–IV filiform and smooth. Setation of tarsi I–IV as in female.
Deutonymph (one paratype).
Gnathosoma as in female. Chelicerae about 45 long. Swollen proximal part of chelicerae 20 long, slender distal part 25 long. Fixed cheliceral digit spinous, 5 long. Palpal tibia and tarsi with smooth setae. Each branch of peritremes 50 long.
Idiosoma 150 long and 200 wide. Dorsum. Propodonotal shield 150 wide and 40 long, punctuate, with 16 plumose setae, 15–20 long. Dorsum covered by setae resembling that of propodonotal shield, about 15 long. Posterior and posteo-lateral setae serrate and 35–40 long. Venter with 20 pairs of short (about 15 long) plumose setae situated in anterior half of idiosoma and 13 pairs of longer (about 40 long) slightly serrate setae situated in posterior half of idiosoma as in
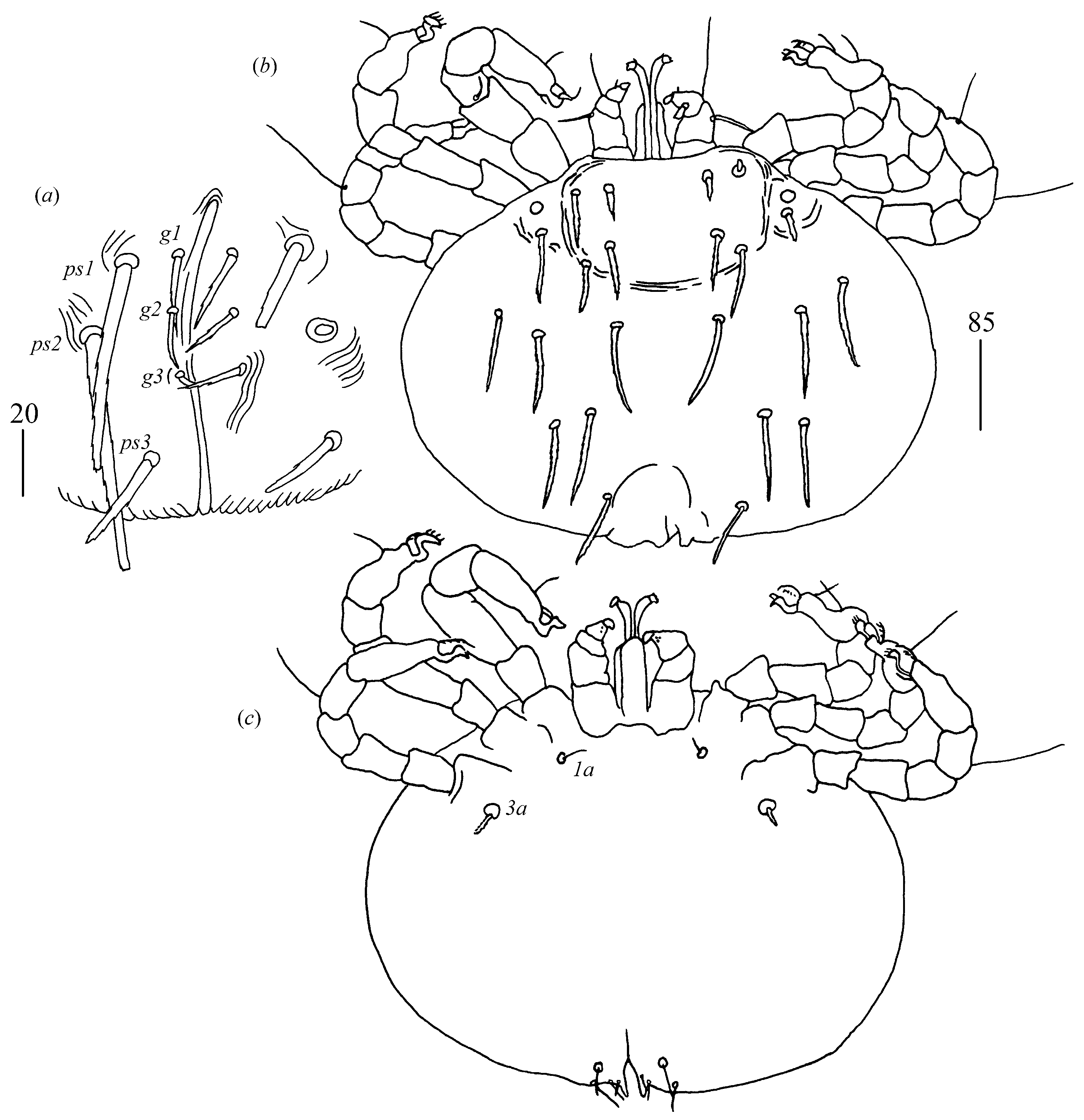
Figure 4. Genital region (
Figure 5a). Genital setae slightly serrate, setae
g1 20 long,
g2 and
g3 about 10 long each. Pseudanal setae
ps1–ps3 slightly serrate 30, 35 and 20 long, respectively. Additional unpaired seta
ps4 on one side of idiosoma present.
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b and
4a arranged in formula: 2–2–2–1. Coxal seta
1a and
1b filiform,
2a,
2b,
3a,
3b and
4a spur-like and serrate. All coxae punctate. Chaetotaxy of trochanters-tibiae I–IV as in female, except for lack of setae
v”TrIV. All setae of tibiae-trochanters I–IV filiform and smooth. Setation of tarsi I–IV as in female.
Larva (range for two paratypes).
Gnathosoma as in female. Chelicerae 35 long. Swollen proximal part of chelicerae 15 long and slender distal part 20 long. Fixed cheliceral digit about 5 long. Setae
dF slightly serrate, and 20 long and setae
dG filiform and smooth, 25 long. Each branch of peritremes 30 long. Hypostome 30 long.
Idiosoma 195–210 wide and 155–185 long. Dorsum with barely discernible punctate propodonotal shield 45 long and 70 wide and with 11 serrate setae (10–30 long) situated as in
Figure 5b. Eyes present laterally to propodonotal shield. Venter devoid of any setation (
Figure 5c). Genital region. Genital setal series represented by three pairs of filiform setae
g1–
g3 5–10 long; pseudanal setal series represented by two pairs of filiform pseudanal setae
ps1 and
ps2 with barely discernible serration. Setae
ps1 10–15 long and setae
ps2 about 20 long.
Legs. Coxal setation arranged in formulae: 2–0–1. Coxal setae
1a filiform and smooth;
1b filiform and slightly serrate; setae
3a thick and serrate. Chaetotaxy of legs I–III as follows: tibiae I–III (5–4–4), genua I–III (1–0–0), femora I–III (3–2–1) and trochanters I–III (0–0–0). Setation of tarsi I–III as in female, except for lack of setae
p” on tarsi I.
Type material.
Female holotype and paratypes: nine females, two males, one deutonymph and two larvae (CES19109) from Hemidactylus giganteus Stoliczka (Squamata: Gekkonidae) (no. CES19101), India, Karnataka, Yerramaranahalli, 13°32′55.4″ N, 77°39′18.5″ E, 12.11.2019, coll. P. Karanth.
Molecular data.
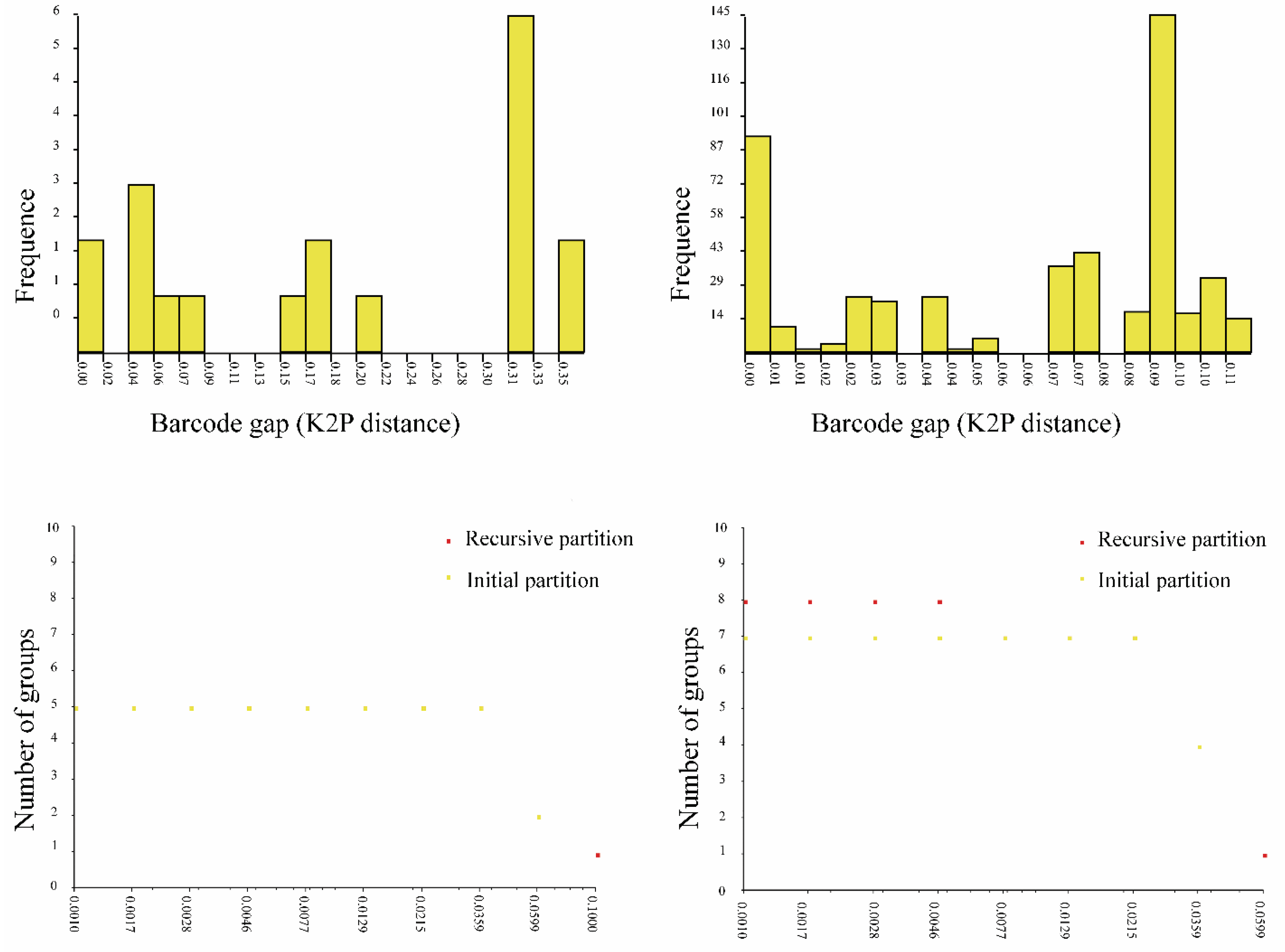
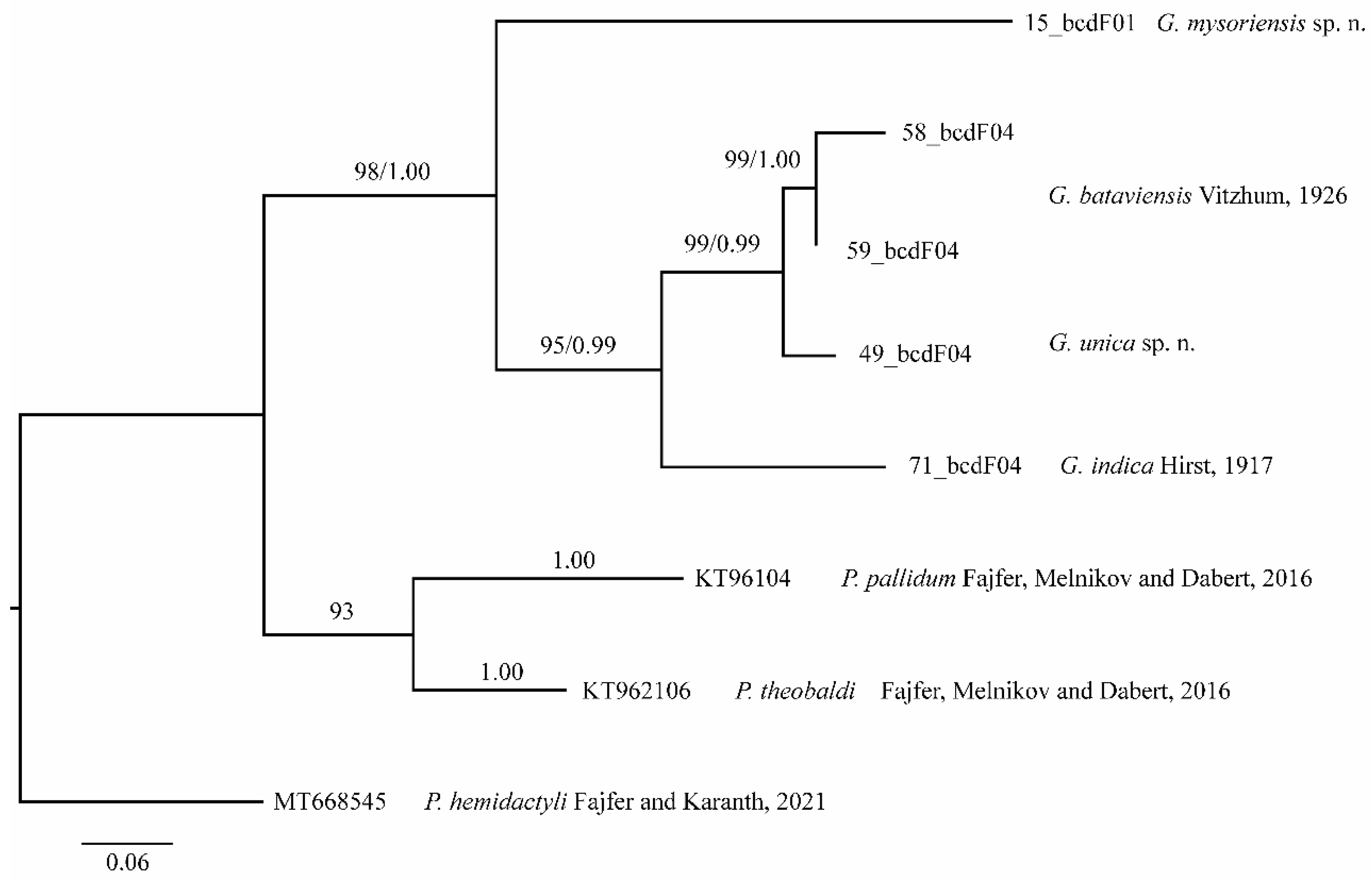
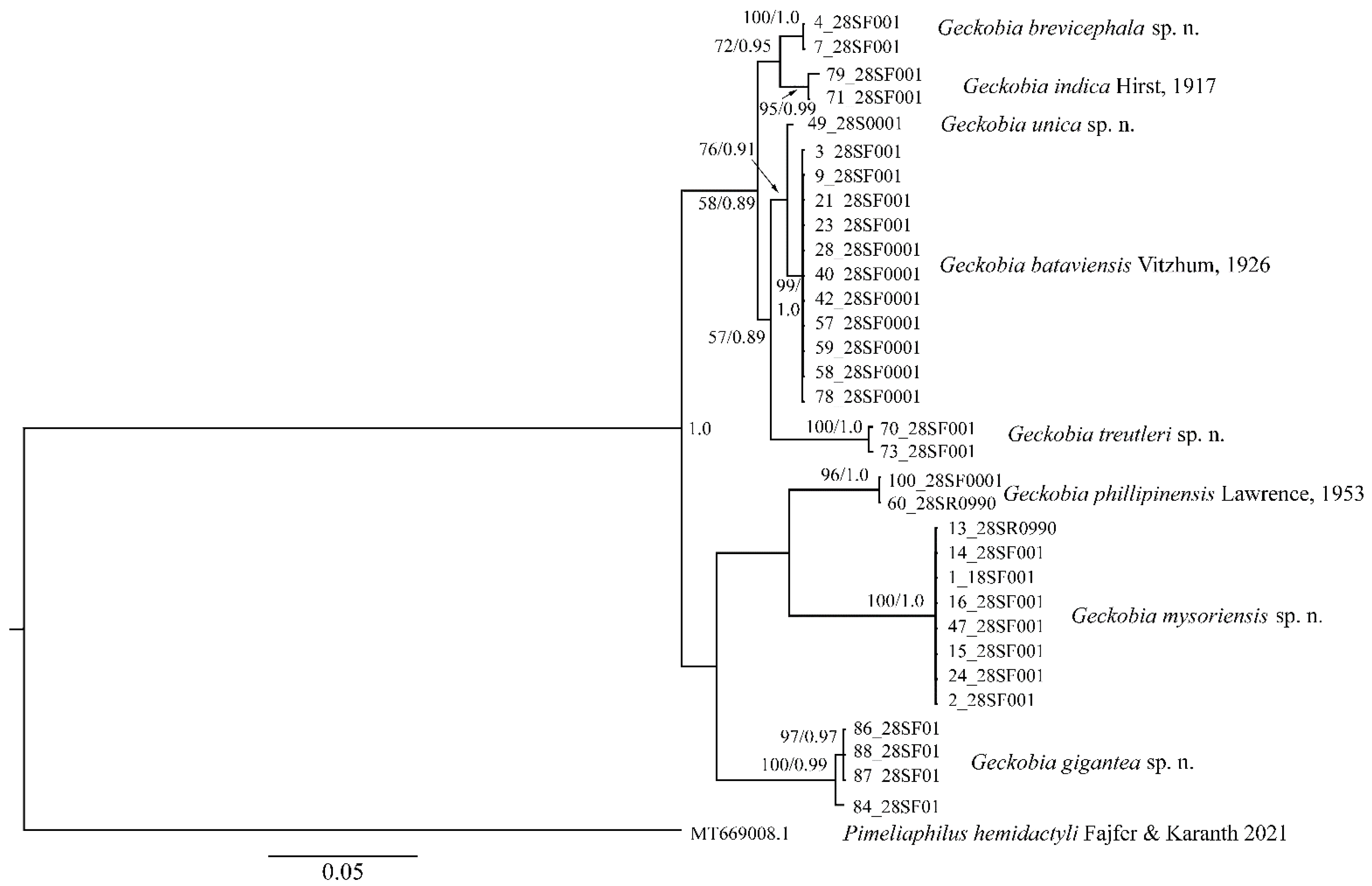
The D2 region of 28S rRNA of G. gigantea is 898 bp long and comprises four sequences represented by two haplotypes differing in terms of five nucleotide positions (0.04%, SD = 0.002, K2P). The 18S region of the rRNA is 1708 bp (two sequences) and 844 bp long (two sequences) and comprises four sequences represented by two haplotypes differing by two nucleotide positions (0.20%, SD = 0.001, K2P).
Etymology.
The species name is derived from the species name of the host.
Differential diagnosis.
This species is most similar to
Geckobia indica Hirst, 1917 from
Hemidactylus gleadowi Murray, India (“Upper Sind” according to original description of Hirst [
43]) [
10,
40]. In both species, the propodonotal shield and eyes are present, the shape and arrangement of the dorsal and ventral setae are the same, palpal setae
dF are thick and serrate and the setation of tarsi I–IV is the same. In
Geckobia gigantea sp. n., the propodonotal shield has a rounded posterior part and 73–81 setae on the shield, which decrease in length from the anterior to posterior part; leg seta
dFIII is absent, and setae
lGIV and
v”TrIV are present. In
G. indica, the propodonotal shield is concave in its posterior part and possesses 34–46 setae, which increase in length from the anterior to posterior part of the shield; leg setae
dFIII are present, and setae
lGIV and
v”TrIV are absent.
Female (holotype, range for eight paratypes).
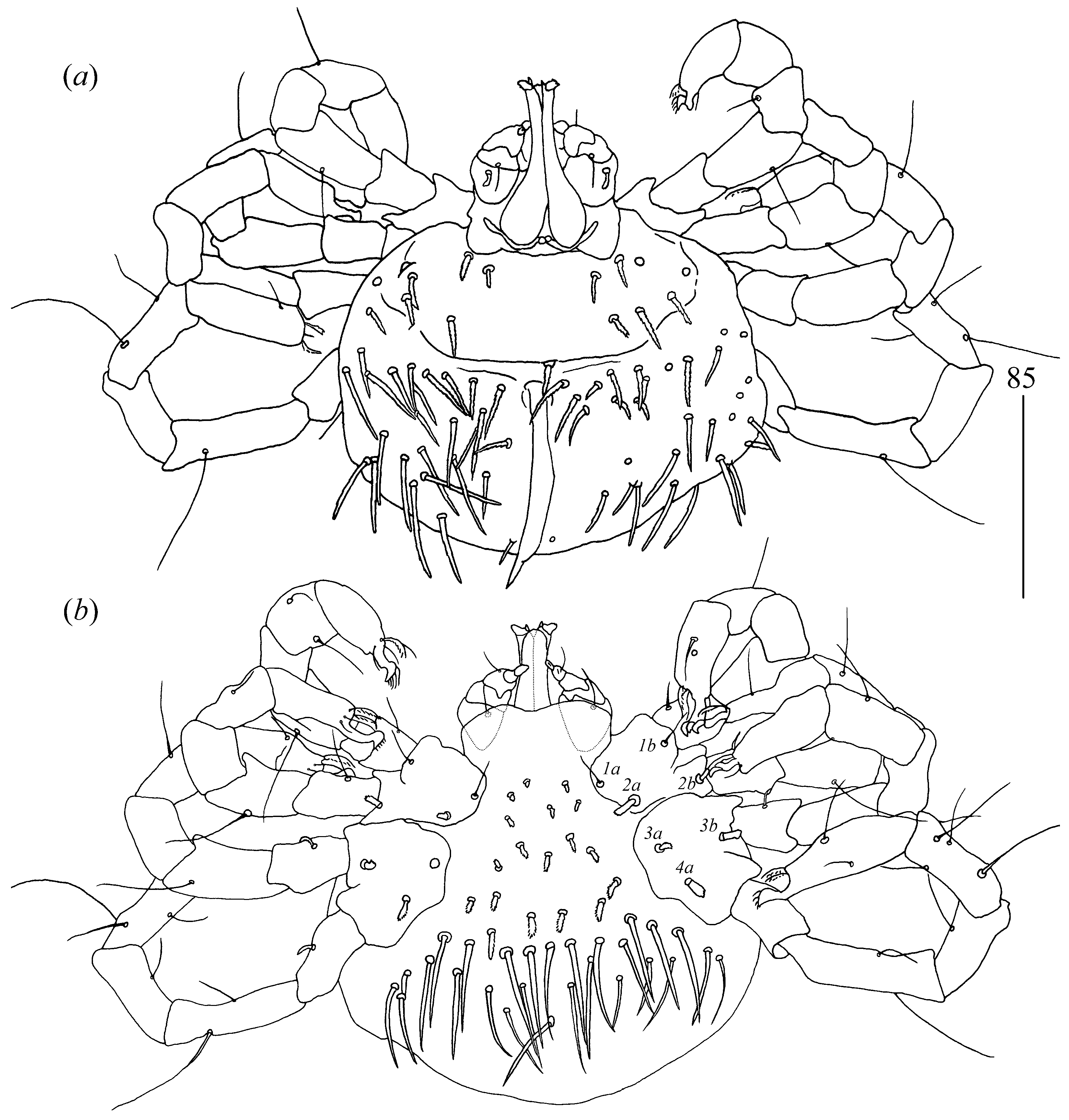
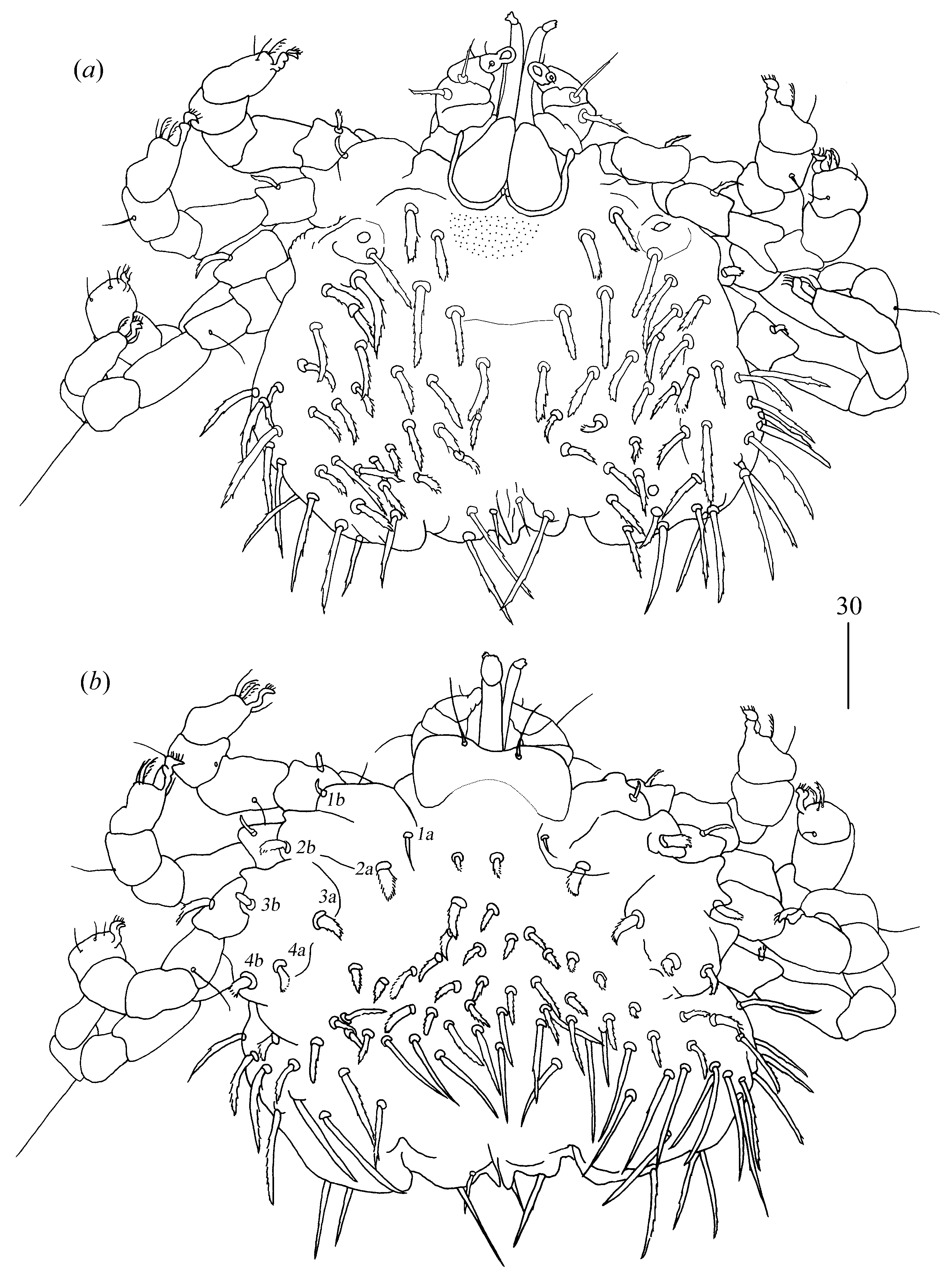
Gnathosoma (
Figure 8a). Chelicerae 110 (105–110) long. Swollen proximal cheliceral part 40 (40–45) long and slender distal part 70 (70) long. Movable cheliceral digit three-pronged. Fixed cheliceral digit spinous, about 5 long. Palpal femur with serrate seta
dF, 20 (20–25) long; palpal genu with smooth setae
dG, 50 (50–55) long. Palpal tibia with three smooth setae:
l′Ti,
l″Ti and
vTi. Palp tarsi with four filiform smooth setae. Supcapitular setae
n serrate and 50 (50) long. Each peritremal branch about 80 long.
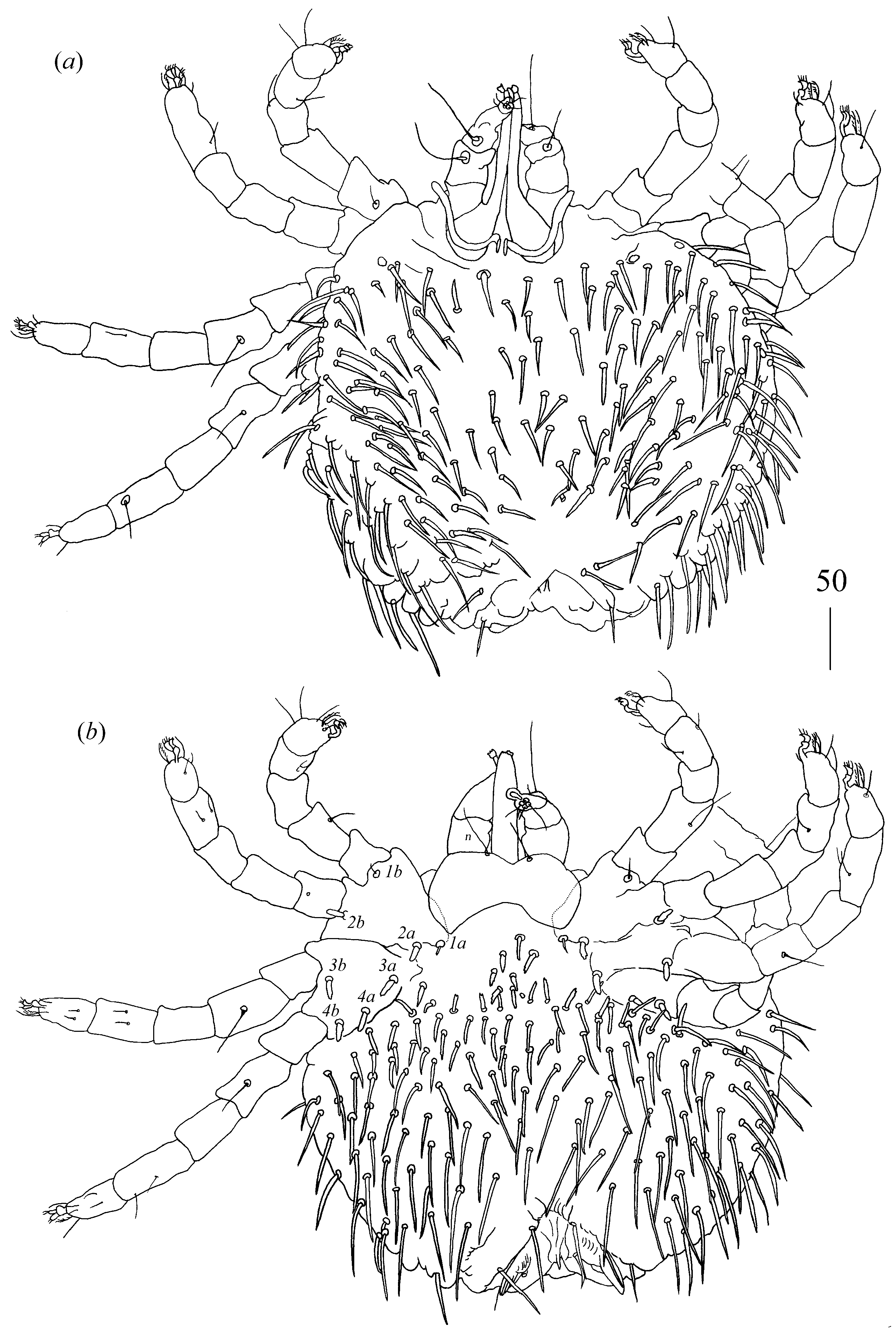
Idiosoma 480 (395–490) long and 515 (445–520) wide. Dorsum covered by numerous serrate setae, 20–35 long, arranged as in
Figure 6. These setae slightly increase from anterior to posterior part of idiosomal dorsum. Small eyes present. Venter (
Figure 7) with numerous setae that cover all idiosoma except for most posterior part. These setae are less serrate than those on dorsum and 30–45 long. Genital series with one pair of filiform slightly serrate genital setae
g1, about 20 long and 10–11 pairs of serrate pseudanal setae
ps (in holotype, 10 setae
ps present on left side and 11 pairs on right side of idiosoma), 35–55 long.
Legs. Coxal setae as follows: 2–2–2–3. Setae
1a and
1b filiform and smooth
2a,
2b,
3a,
3b,
4a,
4b and
4c thick and serrate apically. Setae of tibiae I–IV (5–5–5–5), genua I–IV (1–0–0–1), femora I–IV (3–2–2–2) and trochanters I–IV (1–1–1–1). Setae
vTrI–IV thick and serrate,
vFI–IV short and serrate,
dFI and
lFI–IV filiform and slightly serrate,
lGI and
lGIV with barely discernible serration. Setae
v′TiI–IV,
v″TiI–IV,
d′TiI–IVd″TiI–IV and
l′TiI–IV smooth. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Setae
it′,
it″,
tc′ and
tc″ of legs I in form of euphatidia. Setae
pl′ smooth, setae
a′,
a″,
u′,
u″,
vs′ and
vs″ serrate. Setae
ft smooth, about 5 long.
Deutonymph (one paratype).
Gnathosoma as in female. Chelicerae 70 long. Swollen cheliceral part 20 long and slender cheliceral part 50 long. Setae
dF thick and serrate, 25–30 long; setae
dG filiform with barely discernible serration, about 30 long. Each peritremal branch about 45 long. Subcapitular setae
n 20 long.
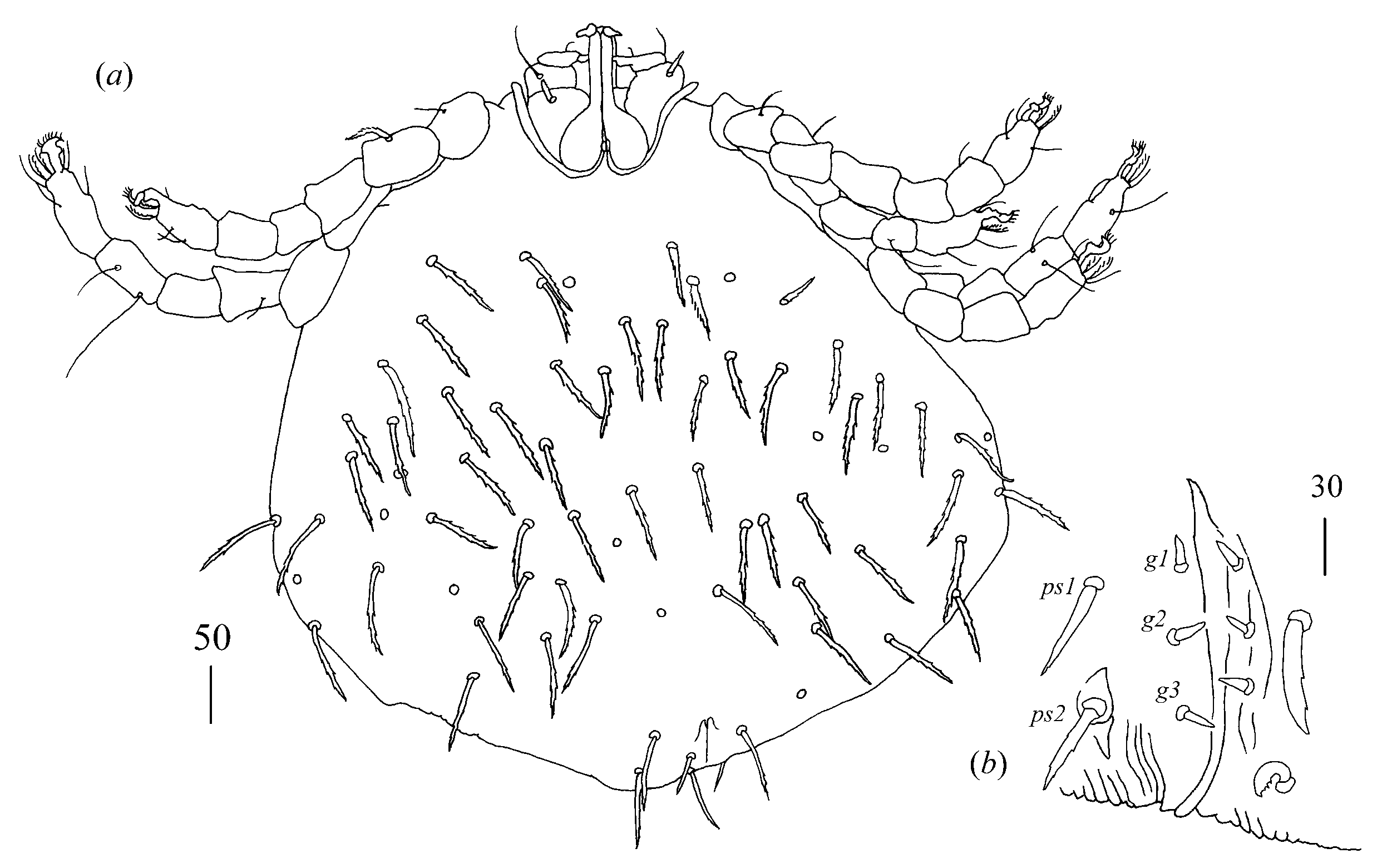
Idiosoma 310–335 wide and 300–315 long. Dorsum with about 30 pairs of dorsal serrate setae (30–40 long) arranged as in
Figure 9a. Venter with five–six short setae, 10–20 long, situated anteriorly and longer setae, about 35 long, situated on remaining part of idiosomal venter, except for most posterior part. Genital region (
Figure 9b) with three genital setae
g1–g3 10–15 long and three pseudanal setae
ps1–
ps3. Setae
ps1 35 long,
ps2 30 long and
ps3 20 long.
Legs. Coxal setae as follows: 2–2–2–2. Coxal setae
1a and
1b filiform with barely visible serration,
2a,
2b,
3a,
3b,
4a and
4b thick and serrate. Setae of tibiae-trochanters I–IV as in female except for lack of setae
lGI and
lFIII.
Protonymph (one paratype).
Gnathosoma as in female. Chelicerae 70 long. Swollen proximal part of chelicerae 30 long and slender distal part about 40 long. Setae
dF and
dG filiform and serrate, about 35 long. Subcapitular setae
n slightly serrate, about 45 long. Peritremes about 55 long.
Idiosoma 165–195 wide and 185–190 long. Dorsum with numerous serrate setae 25–30 long situated as in
Figure 10a. Venter (
Figure 10b) with four short serrate setae situated anteriorly (10–15 long) and numerous longer setae situated in posterior half of idosomal venter, 25–30 long. Genital region with three setae
g1–
g3, about 15 long, and three pseudanal setae
ps1–
ps3. Setae
ps1 15–20 long,
ps2 and
ps3 20–35 long.
Legs. Coxal setae as follows: 2–2–2–2. Setae
1a and
1b filiform and slightly serrate, setae
2a,
2b,
3a,
3b,
4a and
4b plumose, thick and spur-like. Setae of tibiae-trochanters I–IV as in female except for lack of setae
lGI,
lGIV and
lFII–III.
Larva (range for five paratypes). Gnathosoma as in female. Chelicerae about 55 long. Slender proximal part about 30 long and swollen distal part 25–30 long. Fixed cheliceral digit 5 long. Setae dF slightly serrate,15 long, setae dG filiform and smooth 20 long. Each branch of peritremes 65–70 long. Idiosoma 145 long and 190 wide. Dorsum with barely discernible propodonotal shield 50 long and 70 wide. On propodonotal shield, four pairs of setae present; setae situated most laterally 20 long, setae situated medially 15 long. Additional six serrate setae, 20–35 long on the remaining part of idiosoma present. Eyes present laterally to propodonotal shield. Genital area with two pairs of genital setae g1 and g2 and three pairs of pseudanal setae ps1–ps3. Setae g1 and g2 5–10 long, ps1–ps3 15–20 long. Legs. Coxae in formula: 2–1–1. Coxae 1a, 1b and 2b filiform and smooth, setae 3a spur-like and serrate. Setae of tibiae-femora I–III as in female, except for lack of setae v’TrI–III.
Type material examined.
Female holotype and paratypes: two females, one deutonymph, one protonymph and five larvae (CES19110) from Cnemaspis mysoriensis (Jerdon) (Squamata: Gekkonidae), India, Karnataka state, Bangalore, IISc, 19.09.2019, coll. Achyuthan Srikanthan; six female paratypes from same host species and locality, 17.10.2019, coll. Achyuthan Srikanthan, two females from the same host, India, Karnataka state, Bangalore, IISc, 09.10.2019, coll. Caleb Daniel.
Molecular data.
The COI sequence data of 646 bp were generated from two females. Both specimens shared the same COI haplotype. The alignment of the hypervariable D2 region of the nuclear 28S rRNA of G. mysoriensis is 909 bp long and comprises eight sequences represented by one haplotype. The 18S region od rRNA is 1681 bp long and comprises nine sequences represented by one haplotype.
Etymology.
The species name is derived from the species name of the host.
Differential diagnosis.
This species is most similar to
Geckobia uenoi from
Eublepharis splendens Nakamura and Ueno, 1959 (Squamata: Eublepharidae) from the Tokunoshima Island [
14,
44]. In both species, the body is almost circular, and the dorsal setae slightly increase in length from the anterior to posterior part of the idiosoma. The arrangement of the idiosomal setae and setation of tibia–coxae I–IV are the same.
Geckobia mysoriensis sp. n. differs from
G. uenoi in terms of the presence of serrate subcapitular setae
n, densely serrate dorsal setae with long ciliations, the absence of the propodonotal shield and lack of coxal setae
4c. In
G. uenoi, the subcapitular setae
n are smooth, the dorsal setae are slightly serrate with short ciliations, the propodonotal shield is present and coxal setae
4c are absent.
Remarks. For the first time, an active protonymph in a species of the genus
Geckobia has been observed. The protonymph (sample ID: 14_28_SF001 and 14_18SF) and deutonymph (sample ID: 24_SF001 and 24_SF001) of the species are represented by the same haplotype, whereas, simultaneously, the significant morphological differences between the mite stages are observed (e.g., size of idiosoma, shape of setae
dF or leg chaetotaxy pattern). These findings suggest that the protonymph can be either an active or inactive feeding stage depending on the
Geckobia species. So far, only in mites of the genus
Pterygosoma and
Neopterygosoma have active protonymphs been frequently found, e.g., [
17], whereas in the remaining pterygosomatids they are commonly represented by inactive forms.
Female (holotype, range for one paratype).
Gnathosoma. Chelicerae 130 (135) long; swollen cheliceral part 50 (55) long and slender distal part 80 (80) long. Movable cheliceral digit three-pronged. Fixed cheliceral digit spinous, 5 (5) long. Setae
dF and
dG filiform and smooth, 55 (50) and 65 (65) long, respectively. Subcapitular seta
n filiform and smooth, 60 (65) long. Each branch of peritremes 85 (85) long.
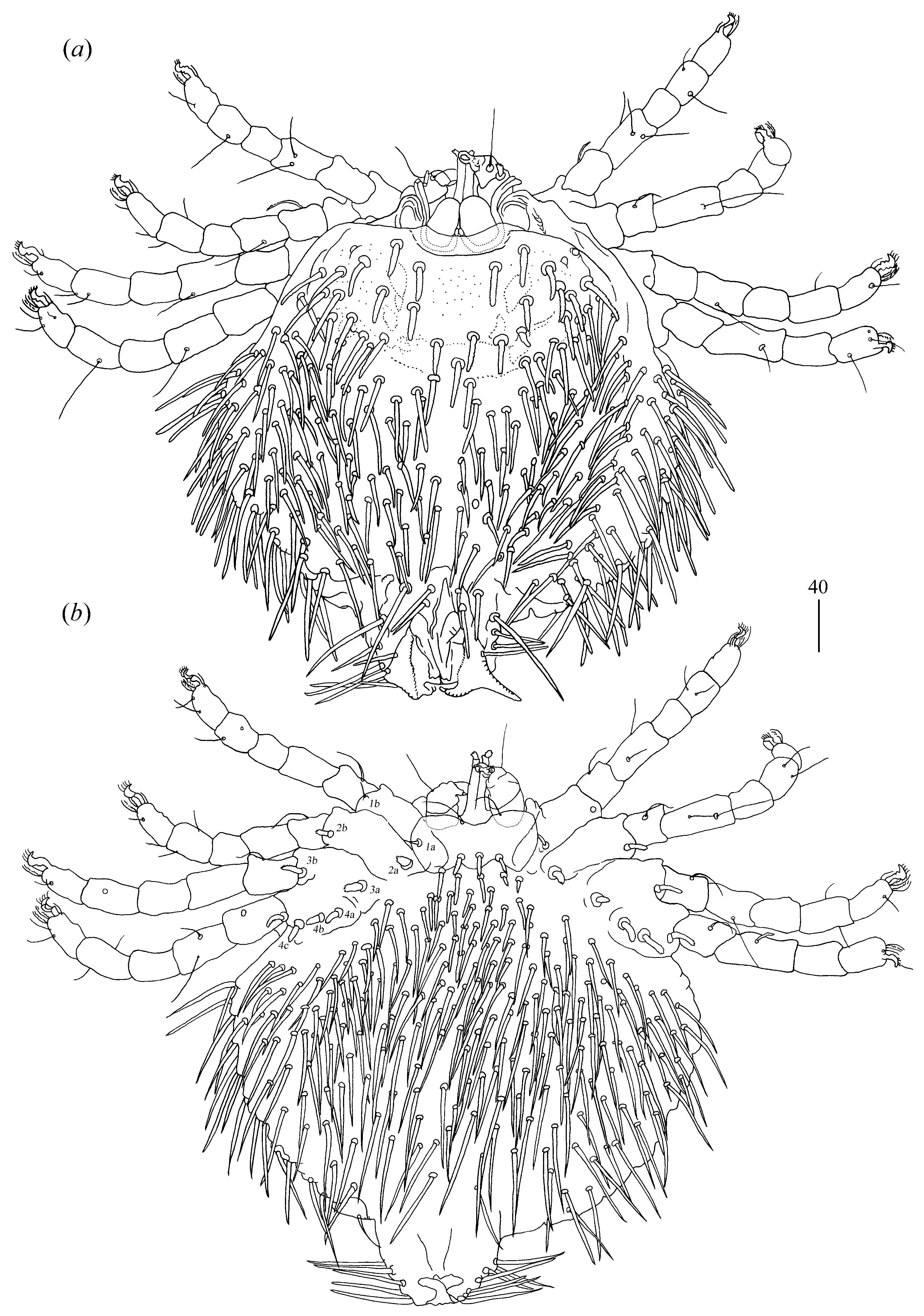
Idiosoma 320 (295) long and 350 (325) wide. Propodonotal shield absent. Small eyes present laterally. Dorsum with numerous serrate setae, 30–50 (25–50) long, distributed as in
Figure 12a. Venter (
Figure 12b) with about 28 short and plumose setae in anterior part, 10–15(10–20) long and longer serrate setae, 25–50 (30–50) long in posterior part. Genital series represented by one slightly serrate seta
g1 and seven serrate pseudanal setae
ps1–ps7. Seta
g1 20–25 (25) long; setae
ps1–ps7 45 (50), 40 (40), 30 (35), 20 (30), 20 (25), 30 (30) and 25 (25) long, respectively. Coxae in formula: 2–2–2–2. Setae
1a and
1b filiform and smooth, setae
2a,
2b,
3a,
3b,
4a,
4b spur-like and serrate at tip. Setae of tibiae I–IV (5–5–5–5), genua I–IV (0–0–0–1), femora I–IV (2–1–1–1), trochanters I–IV (1–1–1–1[0]). All setae of trochanter–genua I–IV filiform and smooth. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Setae
it′,
it″,
tc′ and
tc″ of legs I in form of euphatidia. Setae
pl′ smooth, setae
tc′ and
tc″ of legs II–IV and all setae
vs′,
vs″,
a′,
a″ slightly serrate. Setae
ft smooth and about 5 long. Solenidion
ω1 of legs I about 25 long.
Type material.
Female holotype and one female paratype (CES19111) from Hemidactylus treutleri Mahony, 2009 (under toepads), (Squamata: Gekkonidae) (CES19102), India, Karnataka, Yerramaranahalli, 13°32′55.4″ N, 77°39′18.5″ E, 12.11.2019, coll. P. Karanth.
Molecular data.
The alignment of the D2 region of 28rRNA of G. treutleri is 927 bp long and comprises two sequences represented by two haplotypes differing in terms of three nucleotide positions (0.02%, SD = 0.001, K2P). The 18S region of rRNA was approximately 850 bp long and comprises two sequences represented by two haplotypes differing in one nucleotide position (0.12%, SD = 0.001).
Etymology.
The species name is derived from the species name of the host.
Differential diagnosis.
This species is most similar to
Geckobia keegani Lawrence 1953 from
Hemidactylus frenatus on Philippine Island [
45], Australia [
46] and Costa Rica [
47]. In both species, the idiosoma is circular; the dorsal and ventral setae have the same general shape; the setation of tibiae I–IV, genua I–III, femora I–IV and trochanters I–IV is the same and legs I–IV are subequal in length.
G. treutleri differs from
G. keegani by the presence of smaller idiosoma (320 long and 350 wide), setae in the posterior part of the idiosoma, one pair of genital setae, seta
lGIV and the absence of the propodonotal shield. In
G. keegani, the idiosoma is bigger (513–616 long and 496–630 wide), setae in the posterior part of idiosoma are absent, four pairs of genital setae are present, seta
lGIV is absent and the propodonotal shield is present.
Female (holotype).
Gnathosoma. Chelicerae 80 long. Swollen, proximal part of cheliceral base and slender distal part subequal in length, 40 long. Movable cheliceral digit three-pronged. Fixed cheliceral digit spinous, about 5 long. Palpal femur with filiform serrate setae
dF and
dG 40 and 55 long, respectively. Setae
dF only slightly thicker than setae
dG. Palpal tibia with three smooth setae (
dTi,
l′Ti and
l″Ti) and curved claw. Palpal tarsi with four smooth setae. Subcapitular seta
n filiform and serrate, about 35 long. Each branch of peritremes with barely visible chambers about 75 long. Hypostome with ornamented apex.
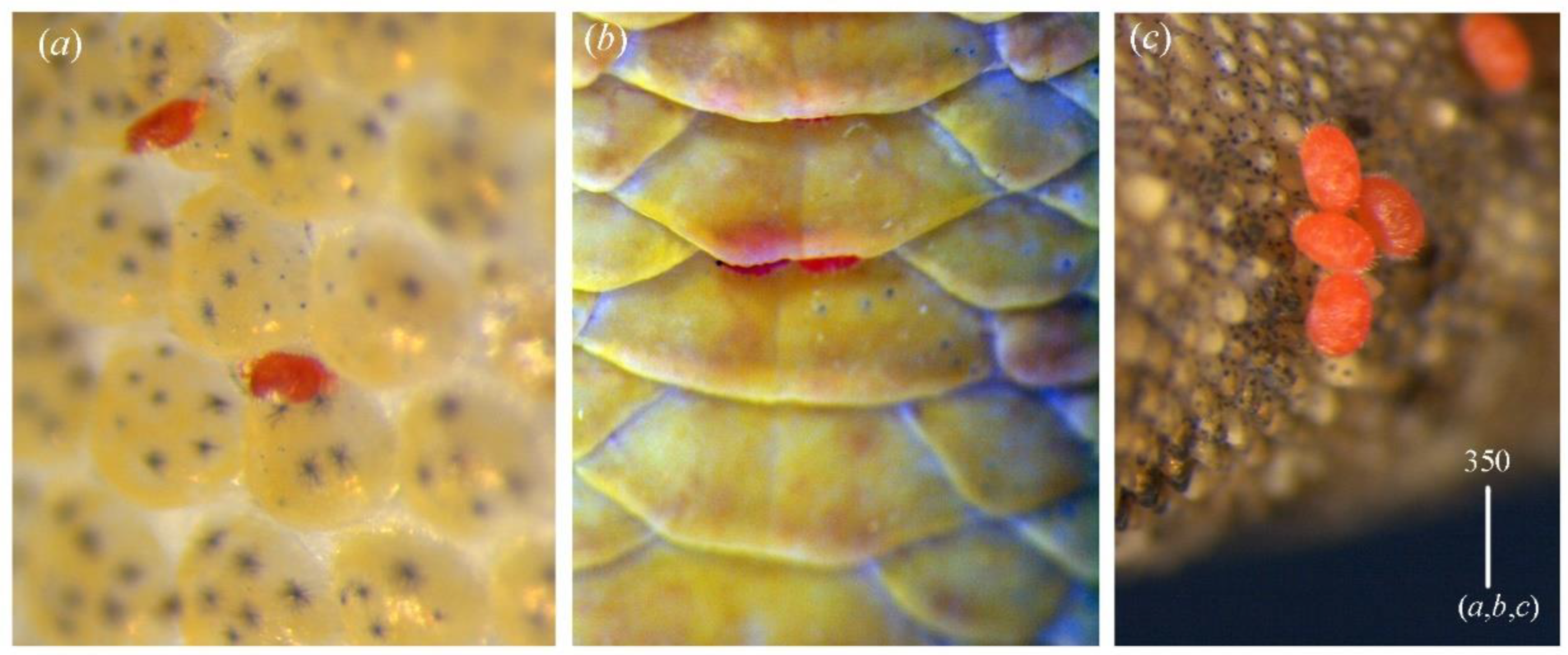
Idiosoma 340 long and 335 wide. Dorsum. Propodonotal shield well outlined, reniform, 110 long and 195 wide, sparsely punctate but in some parts covered by small unsclerotized lacunae (as in
Figure 13a). On propodonotal shield, small eyes situated on lateral margins and 34 stout and plumose setae present. Setae situated antero-medially 25–30 long; setae situated posteriorly and laterally 35–40 long. Posterior to propodonotal shield longer setae (40 long) that decrease in length to median part of idiosoma (25–35 long) and then increase in length in the posterior part (40–55 long). Postero-lateral parts and most posterior part with numerous slightly serrate (50–60 long) and blunt-pointed setae. Venter (
Figure 13b) with 26 short and plumose setae (10–15 long) situated anteriorly and numerous slightly flattened setae (60 long) with slightly serrated tip situated posteriorly. Genital region. Genital setae represented by four pairs of setae
g1–g4. Setae
g1–g3 slightly serrate and 50, 45 and 20 long, respectively. Setae
g4 (30 long) thinner than
g1–g3 and situated on valvae. Pseudanal setal series represented by nine setae
ps about: 75, 35, 40, 55, 60, 55, 45, 60 and 55 long, respectively,
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b,
4a,
4b and
4c arranged in formula: 2–2–2–3. All coxal setae thick and plumose, except for filiform, slightly serrate setae
1a and
1b. Leg chaetotaxy as follows: tibiae I–IV (5–5–5–5), genua I–IV (1 + k–0–1[0]–0), femora I–IV (3–2–2–2) and trochanters I–IV (1–1–1–2). Setae
d′TiI–
IV,
d″TiI,
v′TiI–
IV,
v″TiI–
IV,
l′TiI–
IV,
vFI–II,
lGI and
lGIII filiform and smooth; setae
l′FI–IV filiform and slightly serrate; setae
vFIII and
vFIV thick and serrate, and
TrI–IV filiform and serrate. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Solendion
ω1 (about 25 long) of legs I longer than seta
ft (about 5 long). Setae
tc′,
tc″,
it′,
it″ of leg I represented by euphatidia;
tc′ and
tc″ of legs II–IV,
u′, u″,
vs′,
vs″,
a′,
a″ and
pl′ of legs I–IV filiform.
Type material.
Female holotype (CES19114) from Hemidactylus cf. parvimaculatus India, Karnataka state, Bangalore, NCBS campus, 30 October 2019, coll. Chaitanya R.
Molecular data.
The COI sequence data of 639 bp is generated from the holotype female. The D2 alignment of G. unica is 918 bp long, and the 18S region of rDNA is 1680 bp.
Etymology.
The species name is derived from the Latin adjective “unique” which means “one, uncommon” and refers to unique setation of mite’s trochanter IV.
Differential diagnosis
This species is most similar to
G. bataviensis Vitzthum, 1926 from
Hemidactylus frenatus [
48]. In both species, the idiosoma is almost circular; the propodonotal shield and the eyes are present; dorsal and ventral setae are plumose; palpal setae
dF and
dG are serrate; the chaetotaxy of coxae I–IV, tibiae I–IV and femora I–IV is the same, and four genital setae are present. In
G. unica sp. n., the propodonotal shield is punctate, reniform and convex in its posterior margin; 34 setae are present on the propodonotal shield; dorsal setae decrease in length to the median part of the idiosoma and then increase in length in the posterior part of the idiosoma; subcapitular setae
n are serrate, and leg setae
lGIII and
lTrIV are present. In
G.
bataviensis, the propodonotal shield is smooth and concave in its posterior margin, 40 setae are present on the propodonotal shield, the dorsal setae increase in length posteriorly, subscapular setae
n has barely discernible serration and setae
lGIII and
lTrIV are absent.
Female (holotype, range for one paratype).
Gnathosoma. Chelicerae 90 (90) long; swollen cheliceral part 40 (40) long and slender distal part 50 (55) long. Fixed cheliceral digit rounded. Setae
dF and
dG serrate, 25 (30) and 40 (40) long, respectively. Setae
dF slightly thicker than
dG. Subcapitular setae
n with barely discernible serration, about 60 (60) long. Tibial setae (
v,
l′ and
l″) smooth. Palp tarsi with four slightly serrate setae. Each branch of peritremes 70 (75) long.
Idiosoma 405 (415) long and 665 (670) wide. Dorsum. Propodonotal shield 190 (200) wide and 85 (90) long, punctuate (as in
Figure 14) and with 18 plumose setae, subequal in length, 25–30 (25–30) long. Small eyes present laterally on propodonotal shield. Posteriorly to propodonotal shield, two rows of plumose setae, about 20 (20) long, resembling those on propodonotal shield, present. Medial part of idiosoma with short plumose setae, about 10 (15) long. Most posterior part of idiosoma and lateral parts with serrate setae 70–75 (70–80) long. Venter. Antero-medial part with numerous plumose setae that slightly decrease in length from anterior, 15 (15) long, to posterior part of idiosoma 10 (10) long (
Figure 15). Posterior and postero-lateral parts of idiosomal venter with slightly serrate flattened setae 40–55 long. Genital region. Setae
g1 and
g2 30 (30) long, setae
g3 20 (25) long. Pseudanal setae
ps1–ps10 65–80 (70–80) long.
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b,
4a,
4b and
4c arranged in formula: 2–2–2–3. All coxal setae thick and plumose, except for filiform, slightly serrate setae
1a and
1b. Between coxae I and II, two thick plumose setae present. Leg chaetotaxy as follows: tibiae I–IV (5–5–5–5), genua I–IV (1–0–0–0), femora I–IV (3–2–1–2) and trochanters I–IV (1–1–1–1). Setae
d′TiI–
IV,
d″TiI,
v′TiI–
IV,
v″TiI–
IV,
l′TiI–
IV,
vFI–IV,
dFI–IV,
dGI,
vGI long filiform and smooth,
vFI–
II,
vFIV short and slightly serrate,
lFI short and slightly serrate and setae
lTrI–IV densely serrate. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
P′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Solendion
ω1 of legs I (about 25 long) longer than seta
ft (about 5 long). Setae
tc′,
tc″,
it′ and
it″ of leg I represented by euphatidia;
tc′ and
tc″ of legs II–IV,
u′,
u″,
vs′ and vs. of legs I–IV,
pl′,
a′,
a″ of legs I–IV filiform.
Type material.
Female holotype and one female paratype (CES 19113) from Hemidactylus frenatus, India, Karnataka state, Bangalore, IISc campus, 19.09.2019, coll. Caleb Daniel.
Molecular data.
The D2 alignment of G. brevicephala is 918 bp long and comprises two sequences represented by one haplotype. The 18S region of rRNA is 1679 bp long and comprises two sequences represented by one haplotype.
Etymology.
The species name is derived from the Latin word brevis which means “short” and cephale which means “head” and refers to the short gnathosoma of the species.
Differential diagnosis.
This species is very similar to
Geckobia gibbonsi Bertand and Ineich, 1987 taken from a gecko
Lepidodactylus sp. from Eua Island of Tonga [
49]. In both species, the idiosoma is much wider than long, the propodonotal shield is present, the eyes are present and the chaetotaxy of legs I–IV is the same. This new species differs from
G. gibbonsi in terms of the shape of the propodonotal shield, which is straight in its anterior and posterior part; 18 setae and eyes are present on the shield and short setae in the middle of the idiosomal dorsum. In
G. gibbonsi, the propodonotal shield is concave in the anterior and posterior parts, 14 setae are present on the shield, the eyes are situated outside the shield and the dorsal setae increase in length from the anterior to posterior part of the idiosomal dorsum.
3.2.2. New Data of Already Described Species
G. indica Hirst, 1917: 139; 1926: 185
Figure 8; Haitlinger 2005: 96.
Redescription.
Female (range for five specimens). Chelicerae 90–95 long. Swollen cheliceral part 45–50 long and slender distal part 45–50 long. Movable cheliceral digit three-pronged. Fixed cheliceral digit spinous, about 5 long. Setae
dF thick and densely serrate, serrate
dG slender and slightly serrate (as in
Figure 16c). Setae
dF and
dG subequal in length and 25–30 long. Subcapitular setae
n slightly serrate and about 40 long. Hypostomal apex smooth. Each branch of peritremes 75–85 long.
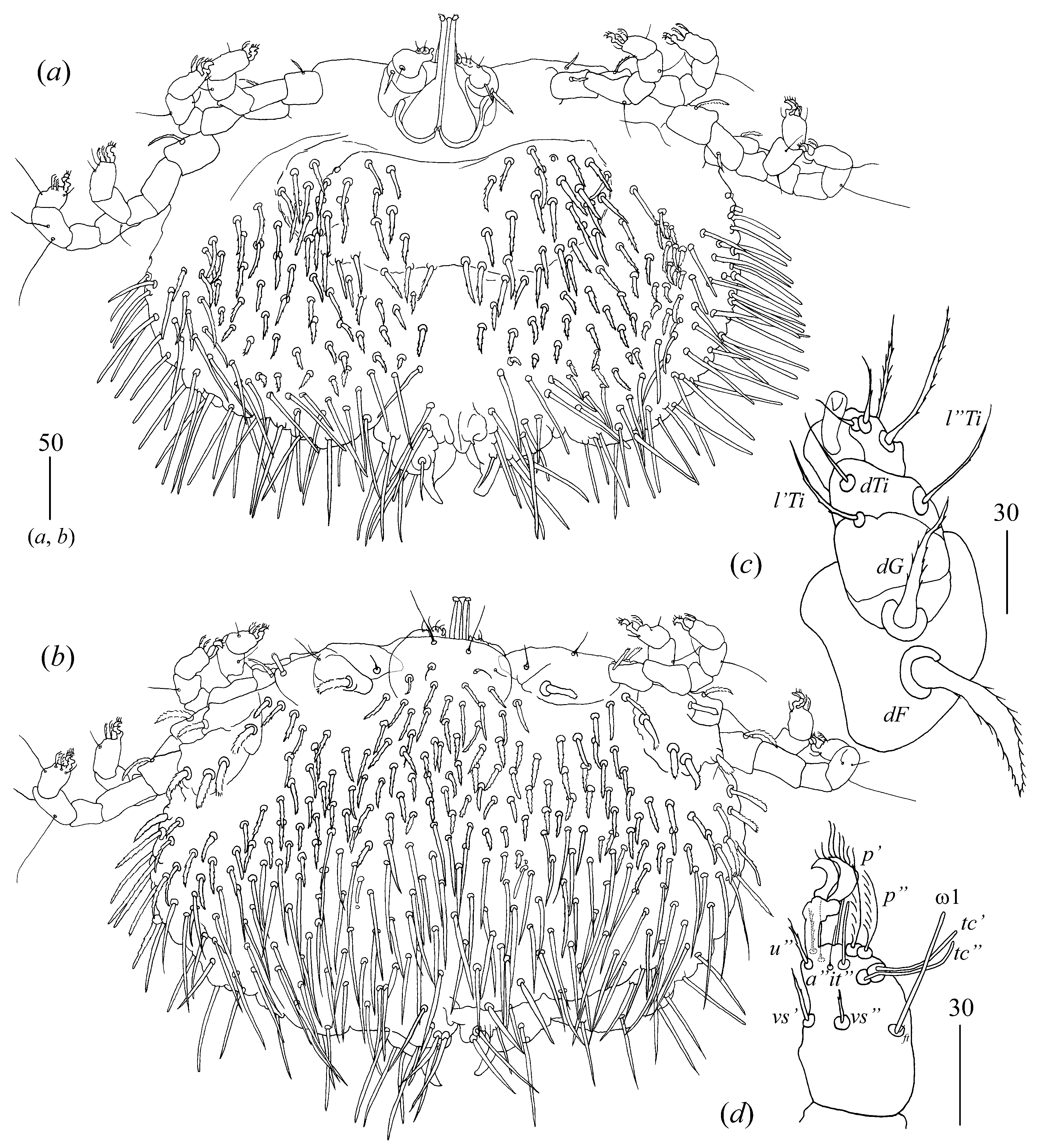
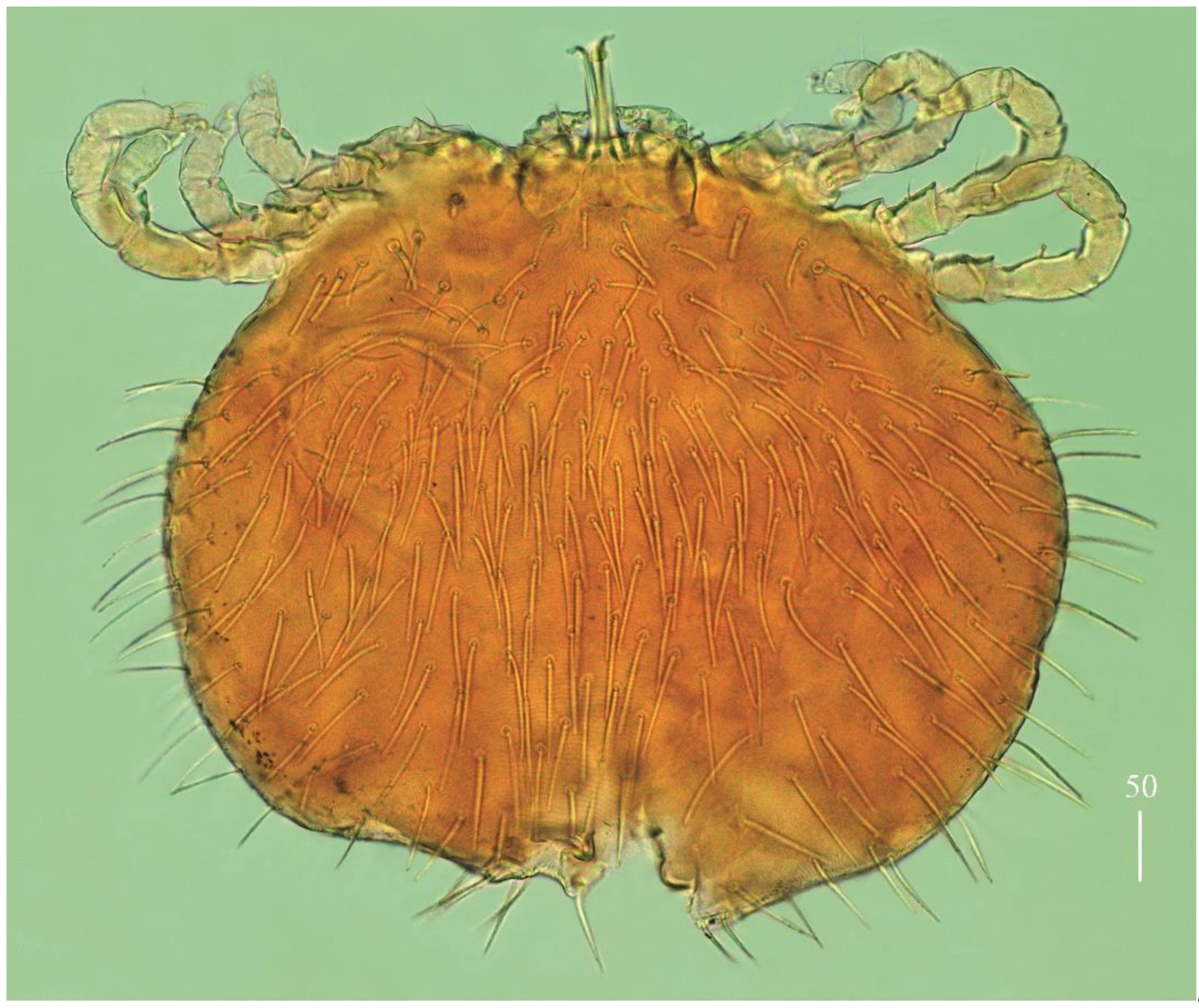
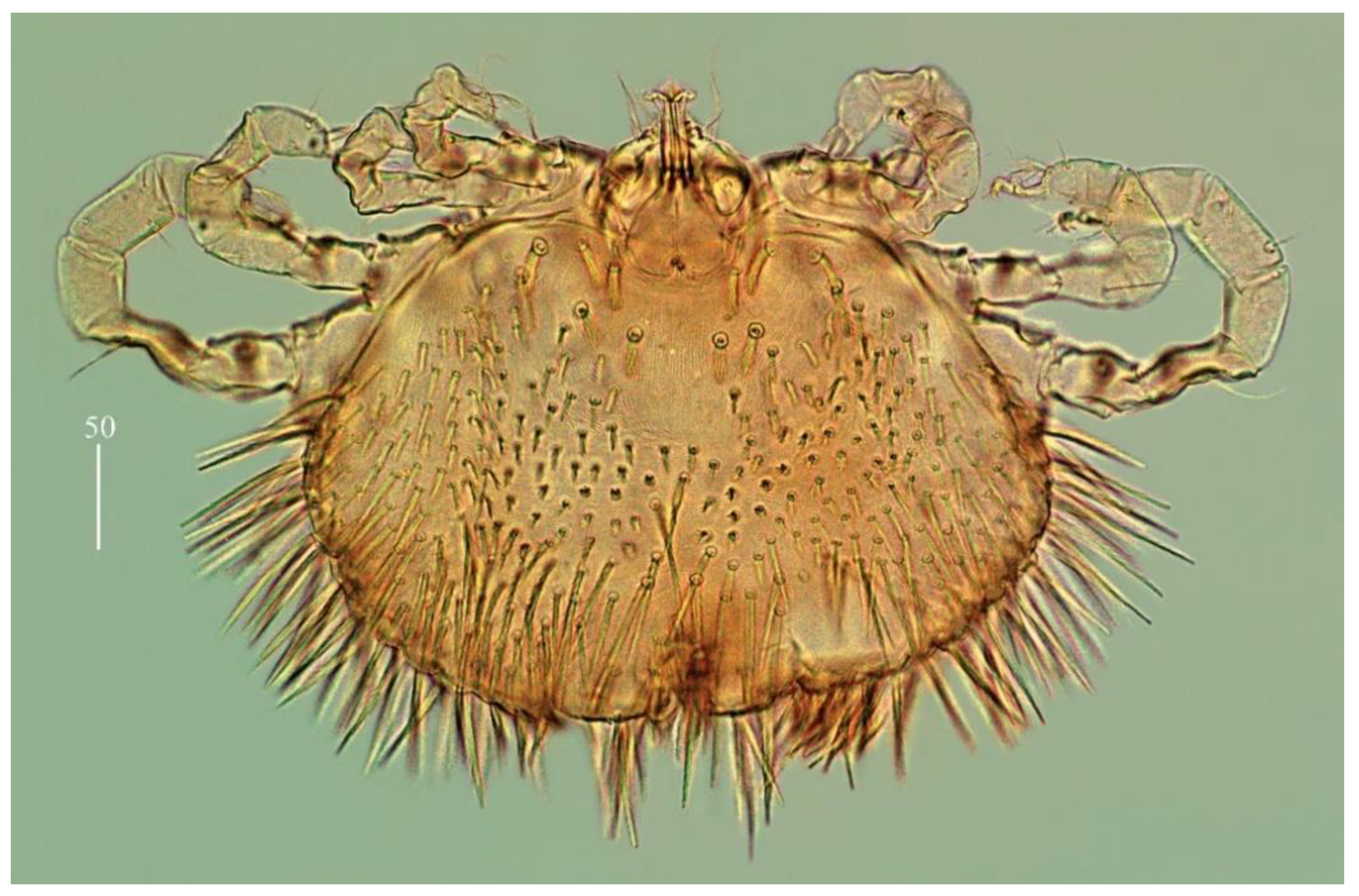
Idiosoma 290–325 long and 430–460 wide. Dorsum (
Figure 16a). Propodonotal shield almost smooth, 95 long and 240 wide. Inconspicuous eyes present laterally between most anterior setae. About 23–26 pairs of plumose setae (20–40 long) present on propodonotal shield. Posterior to propodonotal shield row of setae resembling setae on propodonotal shield (20–30 long) present. Below shorter plumose setae (about 10–15 long) present. Most posteriorly slender slightly serrate setae 50–70 long present. Venter (
Figure 16b) with short plumose setae about 15–20 long situated antero-laterally. Longer numerous setae (60–70 long) situated posteriorly and laterally. Genital area. Genital setal series represented by four pairs of slightly serrate setae
g1–g4, about 30 long. Pseudanal setal series represented by eight blunt-pointed serrate setae
ps1–ps8 60–75 long.
Legs. Coxae in formula: 2–2–2–3. Setae
1a and
1b filiform and smooth, setae
2a and
2b serrate, setae
3a,
3b,
4a,
4b and
4c spur-like and serrate. Setae of tibiae I–IV (5–5–5–5), genua I–IV (1–0–0–0), femora I–IV (3–2–2–2) and trochanters I–IV (1–1–1–1). Setae
l′TiI–IV,
l″TiI–IV,
v′TiI–IV,
v″TiI–IV,
dTiI–IV,
vFI and
vGI smooth; setae
dFIIV and
lFI–IV slightly serrate and setae
vTrI–IV serrate. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1 (
Figure 16d); tarsi II: 10 setae (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Setae
it′,
it″,
tc′ and
tc″ of legs I in form of euphatidia. Setae
a′,
a″,
u′ and
u″ of legs I–IV serrate. Setae
pl′,
tc′ and
tc″ of legs II–IV and setae
vs′ and
vs″ of legs I–IV smooth. Setae
ft smooth and about 5 long. Solenidion
ω1 of legs I about 25 long.
Description.
Male (range for three specimens).
Gnathosoma. Chelicerae about 75 long; swollen cheliceral part about 35 long and slender distal part about 40 long. Setae
dF thick and serrate, about 15 long; setae
dG filiform and smooth, about 30 long. Fixed cheliceral digit spinous, 5 long. Subcapitular seta
n filiform with barely discernible serration about 40 long. Each branch of peritremes about 50 long.
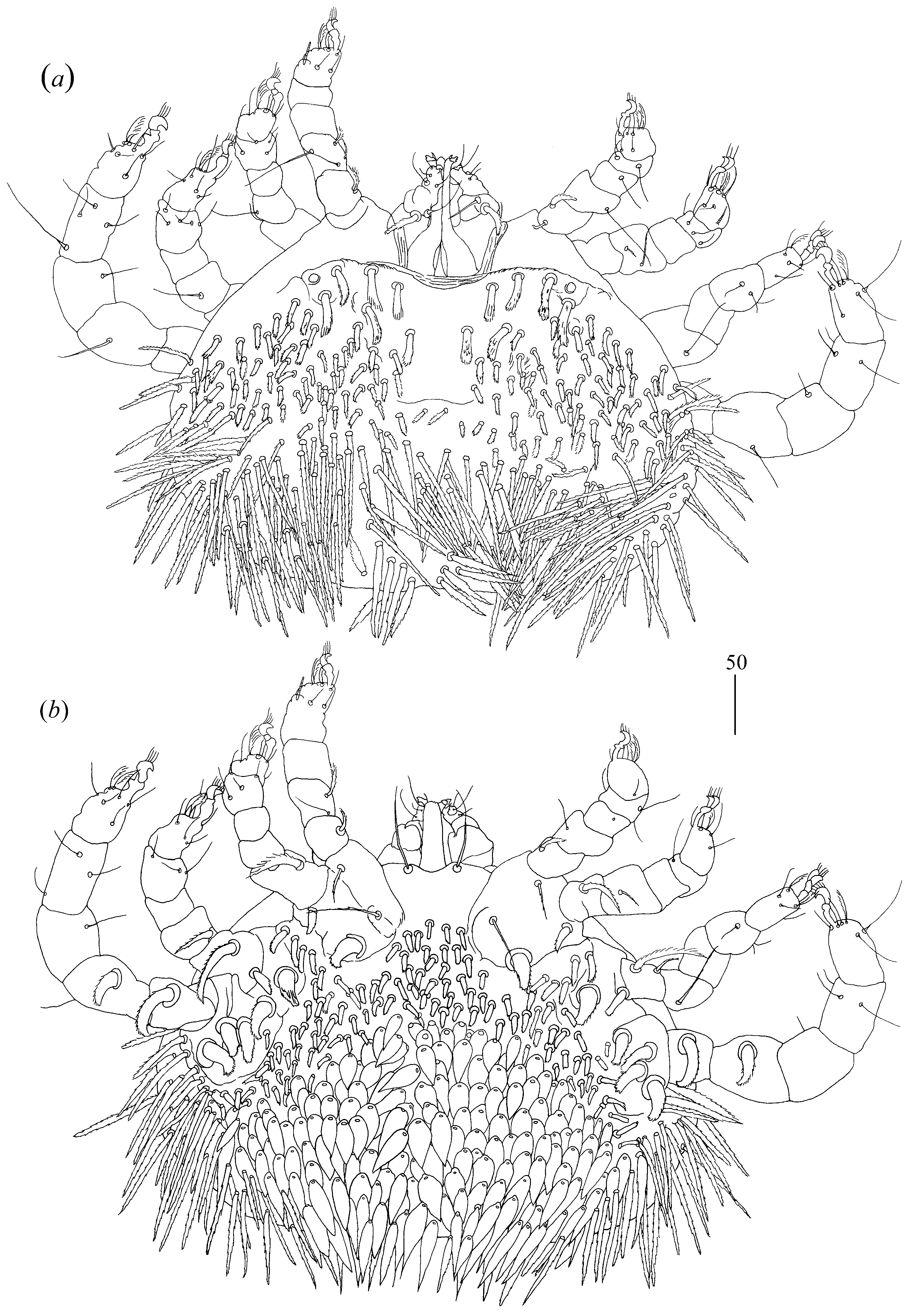
Idiosoma 180–205 wide and 145–185 long with weakly outlined propodonotal shield in anterior part. Dorsum (
Figure 17a) with about 20 pairs of short serrate setae (15–25 long) in anterior part of idiosoma and with 23 pairs of longer serrate setae (about 35 long) in posterior part. Venter (
Figure 17b) with 20–34 pairs of short serrate setae (5–15 long) situated antero-medially and 20–25 pairs of longer setae (20–45 long) situated posteriorly. Aedeagus 90–120 long, bifurcated at the end. Genital cone with one pair of smooth and slightly serrate setae (about 15 long) situated most anteriorly, two pairs of smooth spine-like setae (about 10 long) situated medially and one pair of longer smooth setae (15–20 long) situated most posteriorly.
Legs as in female.
Deutonymph (range for two specimens). Chelicerae 70–75 long. Swollen cheliceral part 30–35 long and slender distal part about 40 long. Fixed cheliceral digit spinous and 5 long. Setae
dG filiform and smooth, 35 long; setae
dF slightly serrate and 20 long. Each branch of peritremes 45 long.
Idiosoma 220–255 long and 320–330 wide. Dorsum (
Figure 18a) propodonotal shield (about 90 wide and 65 long) weakly outlined, with minute punctations in anterior part and five serrate setae, 20–35 long. Laterally to propodonotal shield, small platelets (15 long and 15 wide) with eye and one pair of serrate setae about 20 long present. Posterior to propodonotal shield short plumose setae (about 15 long) present in medial part of idiosoma. These setae increase in size from medial to posterior and lateral parts of idiosoma, 20–40 long. Venter (
Figure 18b) with about 35 short plumose setae in antero-medial part (10–30 long) and about 45 longer serrate setae (40–70 long) in posterior part. Genital setae series represented by three pairs of slightly serrate setae
g1–g3 25, 20 and 15 long, respectively. Setae
ps1–ps3 serrate and 45, 40 and 25 long, respectively.
Legs as in female.
Protonymph (range for three specimens).
Gnathosoma as in female (
Figure 20a). Chelicerae 55–70 long. Swollen cheliceral part 30 long and slender cheliceral part 40 long. Setae
dG filiform, 25–30 long; setae
dF serrate, 20–25 long. Subcapitular setae
n 30 long. Each branch of peritremes 45 long.
Idiosoma 150–195 long and 185–225 wide. Dorsum (
Figure 19a). Propodonotal shield about 85 wide and 55 long; punctate only in its anterior part and with five pairs of serrate setae situated laterally. Small eyes present on lateral platelets (20 wide 15 long), accompanied by one serrate seta, about 25 long. Posterior to propodonotal shield serrate, longer setae (30–35 long) present. These setae longer than setae in posterior (20–25 long) part. Venter (
Figure 19b) with about 20 pairs of plumose short setae 5–15 long. These setae increase in size from anterior to medial part of idiosoma. From medial to posterior part of idiosomal venter, 21 pairs of longer serrate setae (20–40 long) present. Genital setal series represented by slightly serrate setae
g1–g3. Setae
g1 15 long; setae
g2 and
g3 10 long. Pseudanal setal series represented by serrate setae
ps1–
ps3 40, 30–35 and 20 long, respectively.
Legs as in female.
Nymphchrysalis (range for one specimen). Idiosoma 165 long and 360 wide with barely discernible coxae.
Larva (ranges for five specimens).
Gnathosoma. Chelicerae 45–50 long. Swollen cheliceral part and slender distal part subequal in length, 20–25 long. Setae
dF serrate and 15 long; setae
dG filiform and smooth 20 long. Subcapitular seta
n absent. Each peritremal branch about 25 long.
Idiosoma 145–190 long and 180–225 wide. Dorsum with 11 serrate setae. Propodonotal shield 65 wide and 50 long with four pairs of setae: setae situated medially shorter, 15–20 long; setae situated laterally longer, about 25 long (
Figure 20b). Eyes present on barely discernible lateral platelets with accompanied serrate seta about 30 long. Six pairs of setae situated in posterior half of idiosoma, 30–35 long. Genital area with three genital setae
g1–g3 (10–15 long) and two pseudanal setae
ps1 and
ps2 (20–25 long).
Legs. Coxae in formula: 2–1–1. Coxae
1a,
1b and
2b filiform and smooth; setae
3a spur-like and serrate. Setae of tibiae I–III (5–5–4) genua I–III (1–0–0) femora I–III (3–2–2) and trochanters I–III (0–0–0). Setation of tarsi I–III as in female, except for lack of setae
p” on tarsi I.
Type material (not examined).
Types from Hemidactylus gleadowi Murray (Squamata: Gekkonidae), ASIA: India, the northernmost portion of Sind (“upper Sind” according to Hirst 1917).
Type material deposition.
Unknown (it is not stated in the paper, i.e., [
10,
43]).
Non-type material (examined).
Five females, three males, two deutonymphs, three protonymphs, three males, five larvae and one nymphchrysalis (CES19112) from Hemidactylus treutleri Mahony (Squamata: Gekkonidae) (CES19104), India, Karnataka, Yerramaranahalli 13°32′55.4″ N, 77°39′18.5″ E, 12 November 2019, coll. P. Karanth.
Molecular data.
The COI sequence data of 661 bp were generated from one female. The D2 alignment of G. indica is 918 bp long and comprises two sequences represented by two haplotypes differing in terms of two nucleotide positions (0.32%, SD = 0.002, K2P). The 18S region of rRNA is 1688 bp and 844 long and comprises two sequences represented by two haplotypes differing in terms of two nucleotide positions (0.36%, SD = 0.002, K2P).
Host and distribution.
This species was collected from
Hemidactylus gleadowi Murray [
43] and “
Hemidactylus gleadowi (=
Hemidactylus brookii Gray)” from India [
10], the undetermined
Hemidactylus sp. from Sri Lanka [
7] and
Hemidactylus treutleri Mahony (new host) (present study) from India, Karnataka state, Yerramaranahalli (new locality) (present study).
Remarks.
This species was originally described by Hirst [
43] based on females collected from the ventral scales of a gecko
Hemidactylus gleadowi Murray, 1884 from India. The species description was incomplete; only detailed chaetotaxy of idiosomal dorsum and venter was presented. The author neither provides the drawing of the species nor the description of the chaetotaxy of gnathosoma, legs and genital region. Later on, in 1926, Hirst presented a figure with a dorsal view of
Geckobia indica and mentioned that the mites were collected from “
Hemidactylus gleadowi (=
H. brooki)” [
10]. In 1964, Jack, based on loaned-type specimens, described the chaetotaxy of trochanters-tibiae I–IV of
Geckobia indica; however, the chaetotaxy of coxae I–IV and tarsi I–IV was not presented [
16]. Then, Haitlinger [
7] collected
G. indica from undetermined
Hemidactylus sp. from Sri Lanka, exceeding the species distribution. Moreover, the measurements of many structures of the species were presented for the first time [
7]. Here, we present a full description of the species and provide detailed figures based on the material collected from
Hemidactylus treutleri. Additionally, males and immature stages are described for the first time. However, the neotype is not designated for this species because the specimens are not taken from the type of host and locality (see Article 75.3.6 of ICZN [
50,
51]).
G. bataviensis Vitzthum, 1926: 122 Figure 76; Haitlinger 1998: 161
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
Figure 12 and
Figure 13; Prawasti Farajallahrika and Raffiudin 2013: 83
Figure 3; Jacinavicius, Bassini-Silva, Oda, Kaiser 2021: 1
Figure 1 and
Figure 2.
G. gleadoviana Hirst, 1926: 185
Figure 9; Haitlinger 2005: 96.
G. nepalii Hiregaudar, Joshee and Soman, 1959: 66
Figure 2; Haitlinger 2005: 96.
Material examined.
Two females from Hemidactylus frenatus Duméril and Bibron (Gekkonidae) (CES/08/035) India, Kerala state, Peechi, 24.01.2008, coll. P. Karanth; one female from the same host species from India, Karnataka, Bangalore, IISc, 14 September 2019, coll. Caleb Daniel; one female from the same host species and locality, 19 September 2019, coll. Caleb Daniel; one female from the same host species and locality, 19 September 2019, coll. Caleb Daniel; one female from the same host species and locality, 9 October 2019, coll. Caleb Daniel; one specimen from the same host species and locality, 19 October 2019, coll. Caleb Daniel; one female from the same host species, India, Karnataka, Bangalore, NCBS campus, 29 October 2019, coll. Caleb Daniel, one female from the same host species and locality, 29 October 2019, coll. Caleb Daniel; one female from the same host species and locality, 29 October 2019, coll. Caleb Daniel; one female from the same host species, India, Karnataka, Yerramaranahalli, 13°32′55.4″ N 77°39′18.5″ E, 12.11.2019, coll. P. Karanth; one female from H. parvimaculatus India, Karnataka, Bangalore, NCBS campus, 29 October 2019, coll. Chaitanya R.
Molecular data.
The COI sequence data of 660–668 bp were generated from three females of G. bataviensis and represented by two haplotypes. The pairwise comparison of the two COI haplotypes is high and amounts to 4.2% (SE = 0.009). The nuclear data, including 918 positions for the D2 region of 28S rRNA, are obtained for 11 specimens of G. bataviensis and represented by one haplotype. The 18S region of rRNA is 1679 bp long and comprises 12 sequences represented by 7 haplotypes. Intraspecific K2P divergence of 18S region of rDNA in G. bataviensis is 0.28% (SD = 0.001), and the pairwise distance between the haplotypes ranges from 0.1 to 1.05% (SD = 0.002).
Remarks.
We adopt the convention of Domrow [
46] regarding
G. gleadoviana Hirst, 1926,
G. nepalii Hiregauder, Joshee and Soman, 1959 and
G. cosymboti Cuy, 1973 as junior synonyms of
G. bataviensis Vitzthum, 1926. We reject the view of Haitlinger that these might be subspecies of
G. bataviensis [
15] or separate species [
7]. The author in his latter paper [
7] shows in
Table 1 the differences between the of
G. gleadoviana and
G. nepalii (e.g., the species differs in length of the idiosomal and gnathosomal structures) all of which are exclusively quantitative characteristics which cannot be the only differences between species. However, we observe significant variability in size between engorged and non-engorged females of
G. bataviensis (compare
Figure 21 and
Figure 22 with, for example, Figure 76 in the original description or
Figure 2 in [
11]); therefore, quantitative characteristics cannot be the only differences between the species.
Diagnosis.
Female (range for five specimens).
Gnathosoma. Swollen cheliceral part shorter than slender distal part. Subcapitular setae
n slightly serrate. Setae
dF serrate, setae
dG filiform and smooth. Fixed cheliceral digit spinous. Hypostome with small depression present at apex.
Idiosoma 230–270 long and 360–405 wide. Dorsum (
Figure 23) with six thick setae situated antero-laterally. In median and lateral parts short plumose setae present. Posterior, lateral and peripheral setae numerous and serrate. Venter (
Figure 24). Antero-medial and lateral part of idiosoma with short plumose setae. Posterior half of idiosoma with lanceolate setae with minute serration on the surface. Most posterior peripheral setae more elongate and narrower than setae in medial part. Genital area represented by four slender genital setae
g1–g4 and eight densely serrate pseudanal setae
ps1–ps8. Coxal setae
1a and
1b filiform; setae
2b and
3b thick and serrate; setae
2a,
3a,
4a,
4b and
4c spur-like and serrate.
Description.
Male (range for six specimens).
Gnathosoma. Chelicerae 60–70 long. Slender cheliceral part 30–40 long and swollen basal part 30 long. Fixed cheliceral digit about 5 long. Dorsal palpal setae
dF thick and serrate, 10–15 long. Setae
dG filiform and smooth, 25–40 long. Subcapitular setae
n filiform and smooth, 30–50 long. Each branch of peritremes about 45 long.
Idiosoma 130–175 long and 170–190 wide. Dorsum with about 22 pairs of setae situated as in
Figure 25a. Setae situated anteriorly longer (40–45 long) than setae situated posteriorly (about 35 long). Eyes present. Venter (
Figure 25b) with 14 short serrate setae (10–15 long) situated medially and 32 longer serrate setae (15–30 long) situated in posterior half of idiosoma. Aedeagus 100–125 long. Genital cone with three setae 15, 10 and 5 long, respectively. One serrate seta situated laterally to genital cone, about 15 long, present.
Legs. Coxal setation:
1a,
1b,
2a,
2b,
3a,
3b,
4a and
4b arranged in formula: 2–2–2–2. Setae
1a and
1b filiform with barely discernible serration, setae
2b serrate, setae
3b slightly serrate, setae
2a,
3a,
4a and
4b spur-like, thick and serrate. Setae of tibiae I–IV (5–5–5–5), genua I–IV (1–0–0–1), femora I–IV (3–2–2–12) and trochanters I–IV (1–1–1–1). Setae
l′TiI–IV,
l″TiI–IV,
v′TiI–IV,
v″TiI–IV,
dTiI–IV,
dFI–IV,
vFI–III,
lGI and
lGIV filiform and smooth; setae
vFIV slender and slightly serrate; setae
lFI,
lTrI–IV thick and serrate. Setae
dFI–IV much longer than
vFI–IV. Setation of tarsi I: 14 setae (
ft,
tc′,
tc″,
p′,
p″,
a′,
a″,
it′,
it″,
u′,
u″,
vs′,
vs″ and
pl′) and solenidion
ω1; tarsi II: 10 setae (
tc′,
tc″,
P′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″) and
ω1; tarsi III and IV with 10 setae each (
tc′,
tc″,
p′,
p″,
a′,
a″,
u′,
u″,
vs′ and
vs″). Setae
tc′,
tc″,
it′ and
it″ of legs I in form of euphatidia. Setae
pl′ smooth, setae
tc′ and
tc″ of legs II–IV and all setae
vs′,
vs″,
a′ and
a″ slightly serrate. Setae
ft smooth, about 5 long. Solenidion
ω1 of legs I about 25 long. Length of legs I–IV as follows: 120, 130, 160 and 195 long, respectively.
Deutonymph (range for three specimens).
Gnathosoma. Chelicerae 85 long; slender cheliceral part 50 long and swollen basal part 35 long. Palpal femora with dorsal densely serrate seta
dF, 20 long; palpal genua with filiform and smooth dorsal seta
dG 35 long. Subcapitular setae
n filiform and serrate 50 long. Each branch of peritremes 50 long.
Idiosoma 220–245 long and 380–395 wide. Dorsum (
Figure 26a). Propodonotal shield (about 85 wide and 75 long) barely discernible, smooth and slightly concave in its anterior part. Four pairs of serrate thick setae (20–25 long) situated antero-medially on propodonotal shield. Anterior setae situated near propodonotal shield plumose and 10–15 long. Posterior half of idiosoma with thick densely serrate setae 50–70 long. Eyes present on small lateral platelets and accompanied by two densely serrate setae, 20 long. Lateral setae densely serrate and 55–70 long. Venter (
Figure 26b). Anterior part with numerous slightly serrate setae, about 10 long. Medial part with lanceolate setae, about 25 long and posterior part with longer, narrower and serrate setae, 50–60 long. Genital area with five pseudanal thick, flattened and densely serrate setae 35–30 long.
Legs as in female.
Material examined.
Five females, six males and three deutonymphs (CES19115) from Hemidactylus frenatus, India, Karnataka state, Bangalore, NCBS campus, 29.10.2019, coll. Chaitanya R., Karanth P.
Molecular data.
The D2 alignment of
G. phillipinensis is 910 bp long and comprises two sequences represented by one haplotype. The 18S region of rDNA is 1709 long and comprises four sequences represented by three haplotypes differing in one–two nucleotides (0.09%, SE = 0.0001). The amplification of
COI was unsuccessful using both, universal and specific primers [
35,
52]. In all cases,
Wolbachia endosymbiont was detected.
Host and distribution.
G. phillipinensis was described from
Hemidactylus frenatus from the Philippine Islands [
45] and India (new record).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






























