
Effect of Life-History Traits and Habitat Condition on Genetic Diversity between Invasive and Native Plant Populations

 , and
, and 
Abstract
:1. Introduction
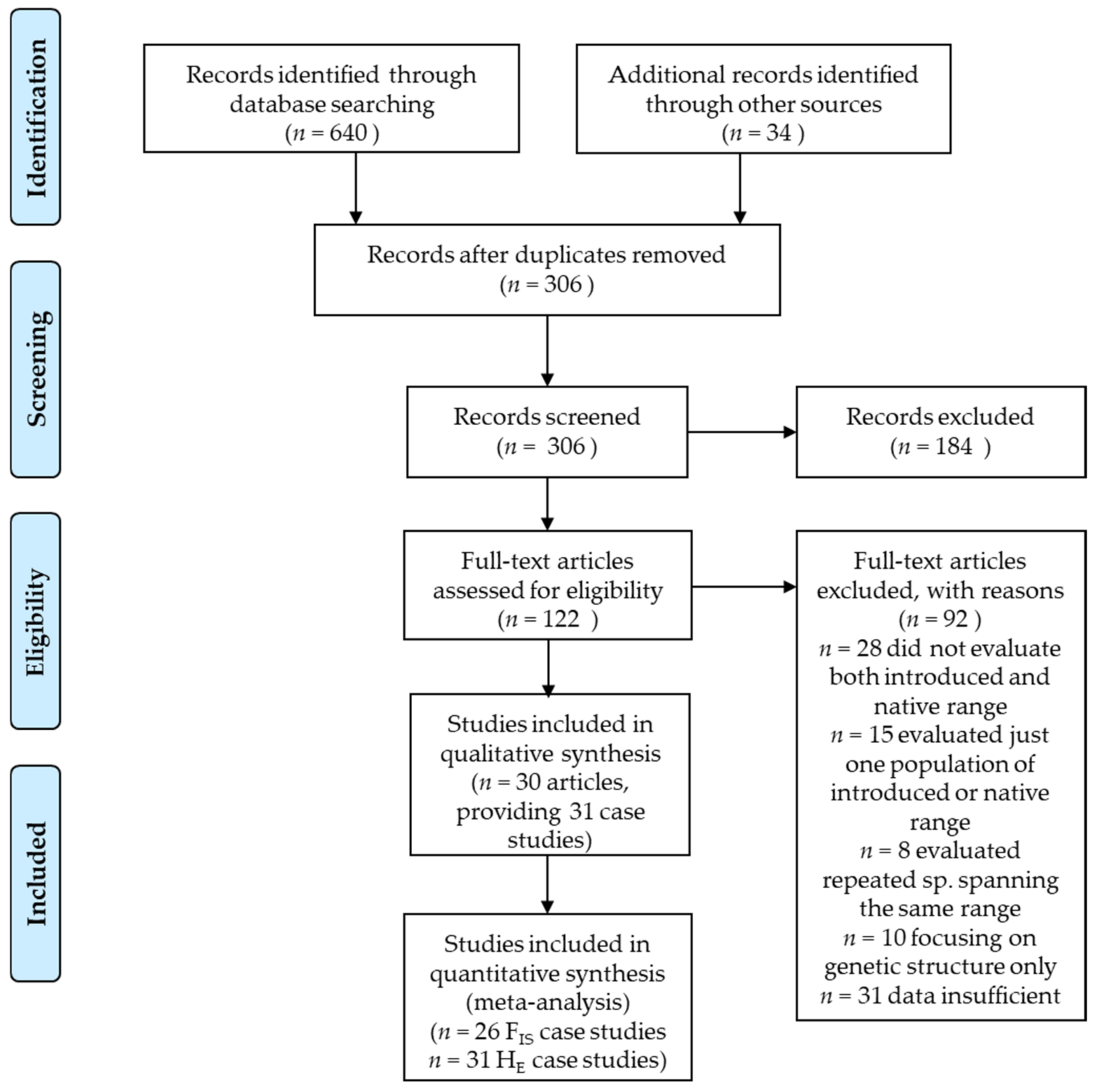
2. Materials and Methods
2.1. Study Selection and Coding
2.2. Effect Size and Moderators
2.3. Effect Model and Publication Bias
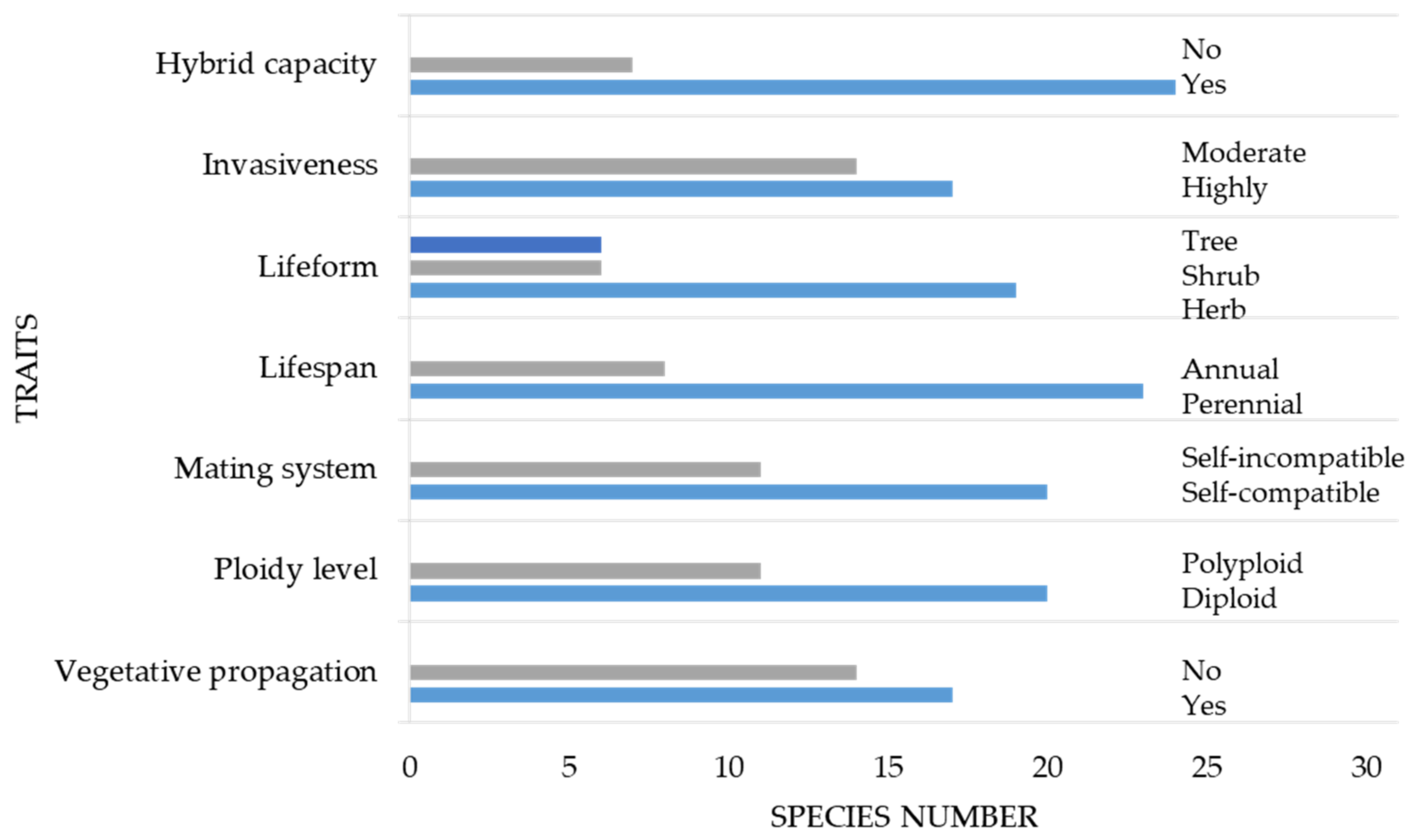
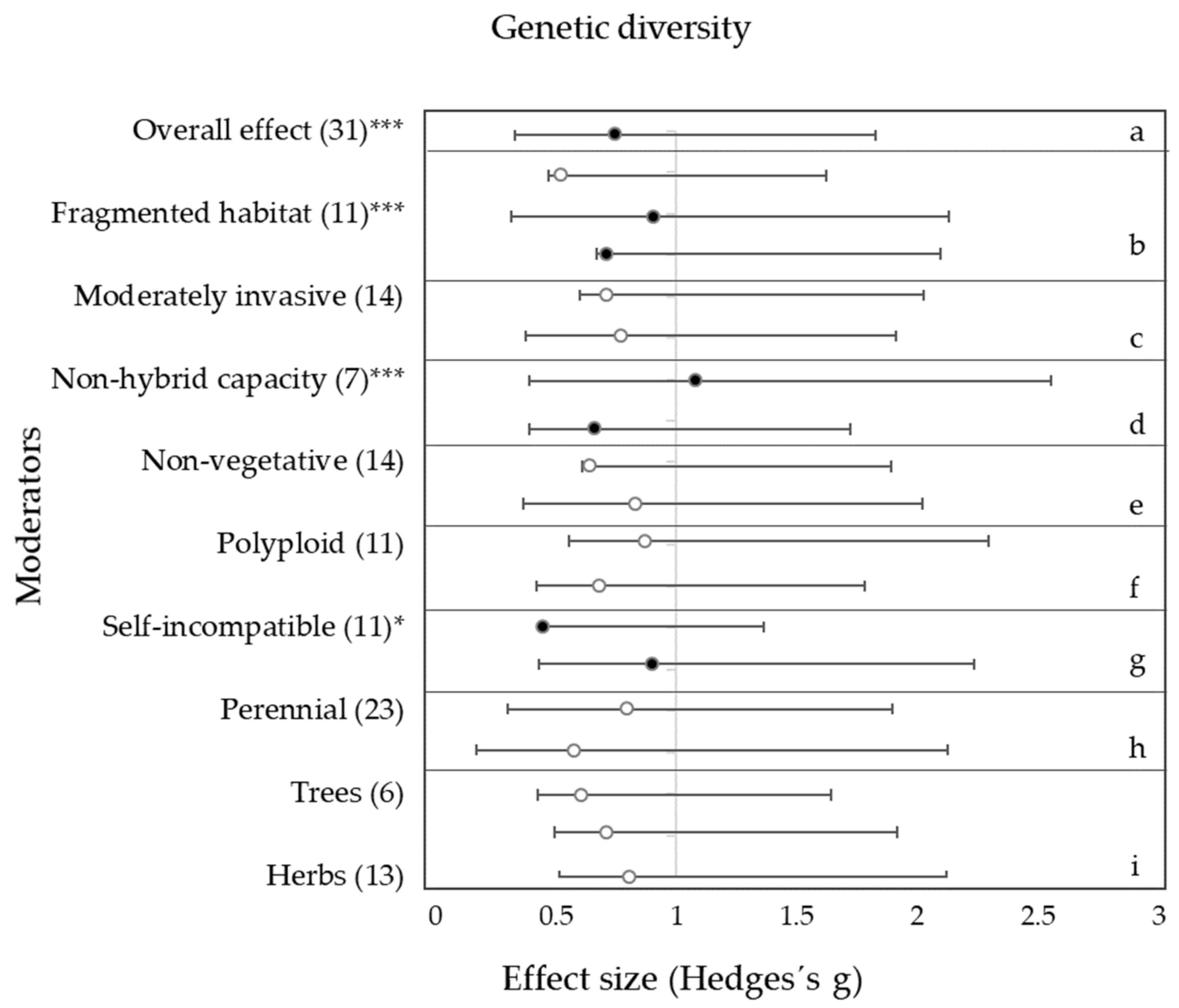
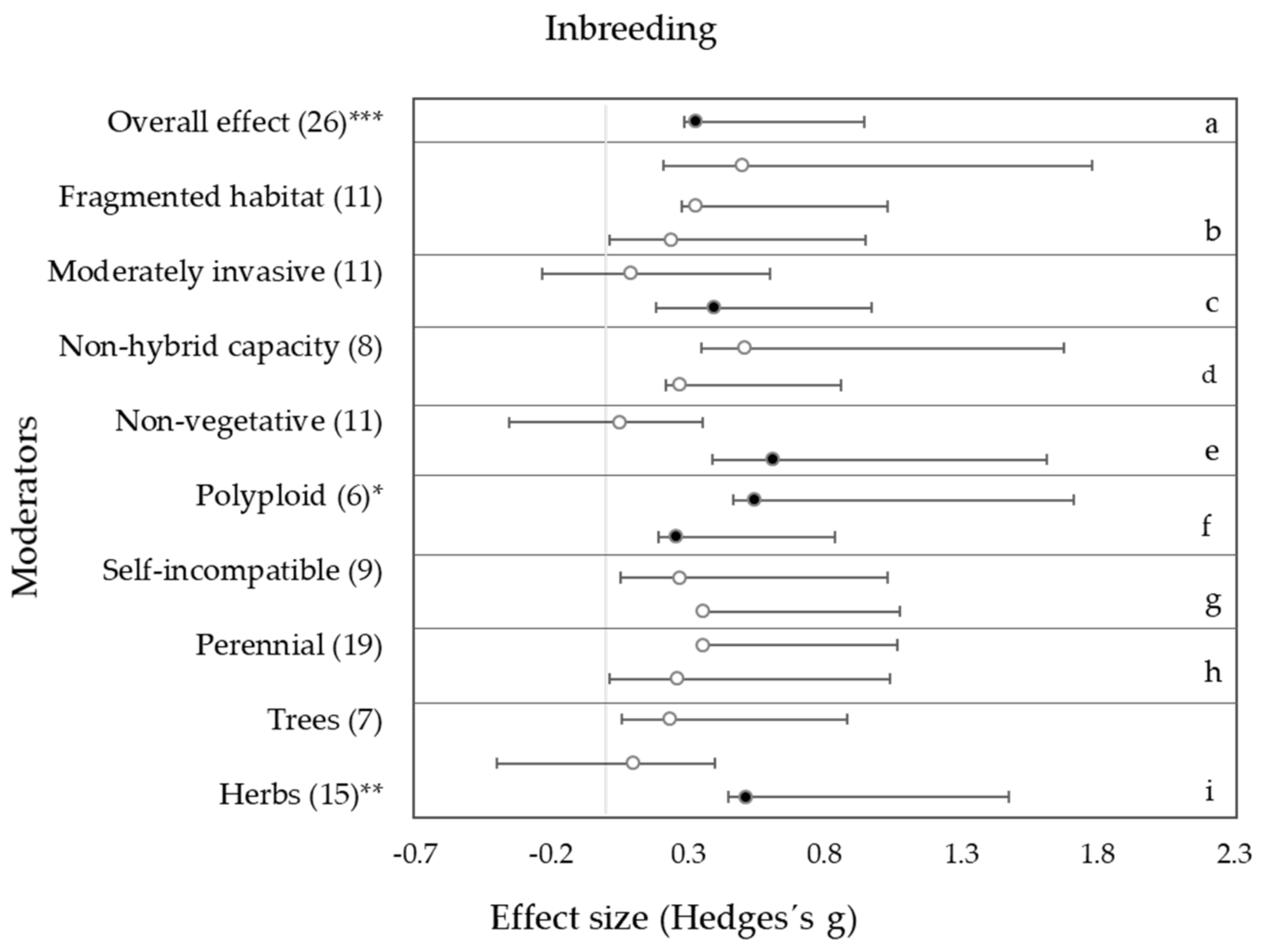
3. Results
3.1. Habitat Condition
3.2. Invasiveness
3.3. Hybrid Capacity and Ploidy Level
3.4. Life-History Traits
3.5. Publication Bias
4. Discussion
4.1. Effects of Habitat Condition on Genetic Diversity and Inbreeding
4.2. Effects of Interspecific Hybridization and Polyploidization on Genetic Diversity and Inbreeding
4.3. Effects of Life-History Traits on Genetic Diversity and Inbreeding
5. Future Research Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Díez, P.; Godoy, O.; Alonso, A.; Gallardo, A.; Saldaña, A. What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis. Ecol. Lett. 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.L.; Kimball, S.; Novak, S.J. Invasion of Plant Communities. In Global Plant Invasions; 2022; pp. 29–51. ISBN 9783030896843. [Google Scholar]
- Riera, M.; Pino, J.; Melero, Y. Impact of introduction pathways on the spread and geographical distribution of alien species: Implications for preventive management in Mediterranean ecosystems. Divers. Distrib. 2021, 27, 1019–1034. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Lockwood, J.L.; Cassey, P. The influence of numbers on invasion success. Mol. Ecol. 2015, 24, 1942–1953. [Google Scholar] [CrossRef] [Green Version]
- Estoup, A.; Ravigné, V.; Hufbauer, R.; Vitalis, R.; Gautier, M.; Facon, B. Is There a Genetic Paradox of Biological Invasion? Annu. Rev. Ecol. Evol. Syst. 2016, 47, 51–72. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Lundquist, L.L. Society for Conservation Biology Introduction: Population Biology, Evolution, and Control of Invasive Species. Biology 2003, 17, 24–30. [Google Scholar]
- Amsellem, L.; Noyer, J.L.; Hossaert-Mckey, M. Evidence for a switch in the reproductive biology of Rubus alceifolius (Rosaceae) towards apomixis, between its native range and its area of introduction. Am. J. Bot. 2001, 88, 2243–2251. [Google Scholar] [CrossRef] [Green Version]
- Husband, B.C.; Barrett, S.C.H. Colonization history and population genetic structure of Eichornia paniculata in Jamaica. Heredity 1991, 66, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R. Resolving the genetic paradox in invasive species. Heredity 2005, 94, 385. [Google Scholar] [CrossRef]
- Zimmermann, H.; Ritz, C.M.; Hirsch, H.; Renison, D.; Wesche, K.; Hensen, I. Highly Reduced Genetic Diversity of Rosa rubiginosa L. Populations in the Invasive Range. Int. J. Plant Sci. 2010, 171, 435–446. [Google Scholar] [CrossRef]
- Castillo, J.M.; Gallego-Tévar, B.; Figueroa, E.; Grewell, B.J.; Vallet, D.; Rousseau, H.; Keller, J.; Lima, O.; Dréano, S.; Salmon, A.; et al. Low genetic diversity contrasts with high phenotypic variability in heptaploid Spartina densiflora populations invading the Pacific coast of North America. Ecol. Evol. 2018, 8, 4992–5007. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.; Harms, N.; Grodowitz, M.; Purcell, M. Genetic structure of Hydrilla verticillata L.f. Royle in eastern China and the Republic of Korea: Implications for surveys of biological control agents for the invasive monoecious biotype. Aquat. Bot. 2018, 149, 17–27. [Google Scholar] [CrossRef]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef]
- Rollins, L.A.; Moles, A.T.; Lam, S.; Buitenwerf, R.; Buswell, J.M.; Brandenburger, C.R.; Flores-Moreno, H.; Nielsen, K.B.; Couchman, E.; Brown, G.S.; et al. High genetic diversity is not essential for successful introduction. Ecol. Evol. 2013, 3, 4501–4517. [Google Scholar] [CrossRef]
- Helsen, K.; Hagenblad, J.; Acharya, K.P.; Brunet, J.; Cousins, S.A.O.; Decocq, G.; De Frenne, P.; Kimberley, A.; Kolb, A.; Michaelis, J.; et al. No genetic erosion after five generations for Impatiens glandulifera populations across the invaded range in Europe. BMC Genet. 2019, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Shang, L.; Li, L.-F.; Song, Z.-P.; Wang, Y.; Yang, J.; Wang, C.-C.; Qiu, S.-Y.; Huang, J.-X.; Nie, M.; Wolfe, L.M.; et al. High Genetic Diversity with Weak Phylogeographic Structure of the Invasive Spartina alterniflora (Poaceae) in China Front. Plant Sci. 2019, 10, 1467. [Google Scholar]
- Li, F.; van Kleunen, M.; Li, J.; Liu, X.; Gao, K.; Zhu, J.; Zhao, X.; Zhao, C.; Li, J. Patterns of genetic variation reflect multiple introductions and pre-admixture sources of common ragweed (Ambrosia artemisiifolia) in China. Biol. Invasions 2019, 21, 2191–2209. [Google Scholar] [CrossRef]
- Kirk, H.; Paul, J.; Straka, J.; Freeland, J.R. Long-distance dispersal and high genetic diversity are implicated in the invasive spread of the common reed, Phragmites australis (Poaceae), in northeastern North America. Am. J. Bot. 2011, 98, 1180–1190. [Google Scholar] [CrossRef]
- Bentley, K.E.; Mauricio, R. High degree of clonal reproduction and lack of large-scale geographic patterning mark the introduced range of the invasive vine, kudzu (Pueraria montana var. lobata), in North America. Am. J. Bot. 2016, 103, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Barrett, S.C.H. Why reproductive systems matter for the invasion biology of plants. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 195–210. [Google Scholar]
- Chung, M.Y.; Son, S.; Herrando-Moraira, S.; Tang, C.Q.; Maki, M.; Kim, Y.-D.; López-Pujol, J.; Hamrick, J.L.; Chung, M.G. Incorporating differences between genetic diversity of trees and herbaceous plants in conservation strategies. Conserv. Biol. J. Soc. Conserv. Biol. 2020, 34, 1142–1151. [Google Scholar] [CrossRef]
- Hamrick, J.; Godt, M. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Rodger, J.G.; Johnson, S.D. Self-pollination and inbreeding depression in Acacia dealbata: Can selfing promote invasion in trees? S. Afr. J. Bot. 2013, 88, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Rosche, C.; Hensen, I.; Schaar, A.; Zehra, U.; Jasieniuk, M.; Callaway, R.M.; Khasa, D.P.; Al-Gharaibeh, M.M.; Lekberg, Y.; Nagy, D.U.; et al. Climate outweighs native vs. nonnative range-effects for genetics and common garden performance of a cosmopolitan weed. Ecol. Monogr. 2019, 89, e01386. [Google Scholar] [CrossRef]
- Ferrero, V.; Barrett, S.C.H.; Castro, S.; Caldeirinha, P.; Navarro, L.; Loureiro, J.; Rodríguez-Echeverría, S. Invasion genetics of the Bermuda buttercup (Oxalis pes-caprae): Complex intercontinental patterns of genetic diversity, polyploidy and heterostyly characterize both native and introduced populations. Mol. Ecol. 2015, 24, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Te Beest, M.; Le Roux, J.J.; Richardson, D.M.; Brysting, A.K.; Suda, J.; Kubesová, M.; Pysek, P. The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot. 2012, 109, 19–45. [Google Scholar] [CrossRef] [Green Version]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Euphytica 2006, 148, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Rosche, C.; Durka, W.; Hensen, I.; Mráz, P.; Hartmann, M.; Müller-Schärer, H.; Lachmuth, S. The population genetics of the fundamental cytotype-shift in invasive Centaurea stoebe s.l.: Genetic diversity, genetic differentiation and small-scale genetic structure differ between cytotypes but not between ranges. Biol. Invasions 2016, 18, 1895–1910. [Google Scholar] [CrossRef]
- Sans, F.X.; Garcia-Serrano, H.; Afán, I. Life-history traits of alien and native Senecio species in the Mediterranean region. Acta Oecologica-Int. J. Ecol. 2004, 26, 167–178. [Google Scholar] [CrossRef]
- Lee, C.E.; Gelembiuk, G.W. Evolutionary origins of invasive populations. Evol. Appl. 2008, 1, 427–448. [Google Scholar] [CrossRef]
- Williams, D.A.; Overholt, W.A.; Cuda, J.P.; Hughes, C.R. Chloroplast and microsatellite DNA diversities reveal the introduction history of Brazilian peppertree (Schinus terebinthifolius) in Florida. Mol. Ecol. 2005, 14, 3643–3656. [Google Scholar] [CrossRef]
- Chabrerie, O.; Verheyen, K.; Saguez, R.; Decocq, G. Disentangling relationships between habitat conditions, disturbance history, plant diversity, and American black cherry (Prunus serotina Ehrh.) invasion in a European temperate forest. Divers. Distrib. 2008, 14, 204–212. [Google Scholar] [CrossRef]
- Henry, P.; Le Lay, G.; Goudet, J.; Guisan, A.; Jahodová, S.; Besnard, G. Reduced genetic diversity, increased isolation and multiple introductions of invasive giant hogweed in the western Swiss Alps. Mol. Ecol. 2009, 18, 2819–2831. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Siemann, E.; Rogers, W.E. Geographic distribution of genetic variation among native and introduced populations of Chinese tallow tree, Triadica sebifera (Euphorbiaceae). Am. J. Bot. 2011, 98, 1128–1138. [Google Scholar] [CrossRef]
- Hernández, F.; Presotto, A.; Poverene, M.; Mandel, J.R. Genetic Diversity and Population Structure of Wild Sunflower (Helianthus annuus L.) in Argentina: Reconstructing Its Invasion History. J. Hered. 2019, 110, 746–759. [Google Scholar] [CrossRef]
- Banks, S.C.; Cary, G.J.; Smith, A.L.; Davies, I.D.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B.; Peakall, R. How does ecological disturbance influence genetic diversity? Trends Ecol. Evol. 2013, 28, 670–679. [Google Scholar] [CrossRef]
- Devictor, V.; Robert, A. Measuring community responses to large-scale disturbance in conservation biogeography. Divers. Distrib. 2009, 15, 122–130. [Google Scholar] [CrossRef]
- Daehler, C. Performance comparisons of co-occurring native and alien invasive plants: Implications for conservation and restoration. Annu. Rev. Ecol. Evol. Syst 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Pysek, P.; Richardson, D. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions; Nentwig, W., Ed.; Springer: New York, NY, USA, 2007; pp. 97–125. [Google Scholar]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Palacio-López, K.; Gianoli, E. Invasive plants do not display greater phenotypic plasticity than their native or non-invasive counterparts: A meta-analysis. Oikos 2011, 120, 1393–1401. [Google Scholar] [CrossRef]
- Oduor, A.M.O.; Leimu, R.; van Kleunen, M. Invasive plant species are locally adapted just as frequently and at least as strongly as native plant species. J. Ecol. 2016, 104, 957–968. [Google Scholar] [CrossRef]
- Tarin, D.; Pepper, A.E.; Goolsby, J.A.; Moran, P.J.; Arquieta, A.C.; Kirk, A.E.; Manhart, J.R. Microsatellites Uncover Multiple Introductions of Clonal Giant Reed (Arundo donax). Invasive Plant Sci. Manag. 2013, 6, 328–338. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. The Genetical Structure of Populations. Ann. Hum. Genet. 1950, 15, 323–354. [Google Scholar] [CrossRef]
- González, A.V.; Gómez-Silva, V.; Ramírez, M.J.; Fontúrbel, F.E. Meta-analysis of the differential effects of habitat fragmentation and degradation on plant genetic diversity. Conserv. Biol. J. Soc. Conserv. Biol. 2020, 34, 711–720. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2013; ISBN 159726606X. [Google Scholar]
- CABI Invasive Species Compendium. Available online: www.cabi.org/isc (accessed on 12 December 2021).
- Bonett, D.G. Confidence intervals for standardized linear contrasts of means. Psychol. Methods 2008, 13, 99–109. [Google Scholar] [CrossRef]
- Bonett, D.G. Meta-analytic interval estimation for standardized and unstandardized mean differences. Psychol. Methods 2009, 14, 225–238. [Google Scholar] [CrossRef]
- Koricheva, J.; Gurevitch, J.; Mengersen, K. Handbook of Meta-Analysis in Ecology and Evolution; Koricheva, J., Gurevitch, J., Mengersen, K., Eds.; Princeton University Press: Princeton, NJ, USA, 2013; ISBN 9780691137292. [Google Scholar]
- Hedges, L. V Distribution theory for Glass’s estimator of effect size and related estimators. J. Educ. Stat. 1981, 6, 107–128. [Google Scholar] [CrossRef]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. Introduction to Meta-Analysis; John Wiley & Sons: Chichester, UK, 2009. [Google Scholar]
- Egger, M.; Smith, G.D.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. Br. Med. J. 1997, 315, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Jennions MD, M.A. Publication bias in ecology and evolution: An empirical assessment using the ‘trim and fill’ method. Biol. Rev. 2002, 77, 211–222. [Google Scholar] [CrossRef]
- Borenstein, M.; Hedges, L.; Higgins, J.; Rothstein, H. Comprehensive Meta-Analysis; Biostat: Englewood, NJ, USA, 2005. [Google Scholar]
- Alexander, J.M.; Poll, M.; Dietz, H.; Edwards, P.J. Contrasting patterns of genetic variation and structure in plant invasions of mountains. Divers. Distrib. 2009, 15, 502–512. [Google Scholar] [CrossRef]
- Zhu, B.R.; Barrett, S.C.H.; Zhang, D.Y.; Liao, W.J. Invasion genetics of Senecio vulgaris: Loss of genetic diversity characterizes the invasion of a selfing annual, despite multiple introductions. Biol. Invasions 2017, 19, 255–267. [Google Scholar] [CrossRef]
- Vilá, M.; Valladares, F.; Traveset, A.; Santamaría, L.; Castro, P. Invasiones Biológicas; Cyan, S.A., Ed.; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2008; ISBN 9788400085452. [Google Scholar]
- Honnay, O.; Jacquemyn, H. Susceptibility of common and rare plant species to the genetic consequences of habitat fragmentation. Conserv. Biol. J. Soc. Conserv. Biol. 2007, 21, 823–831. [Google Scholar] [CrossRef]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrerias-Diego, Y.; Lobo, J. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and methodological approaches. Mol. Ecol. 2008, 17, 5177–5188. [Google Scholar] [CrossRef]
- Breed, M.F.; Ottewell, K.M.; Gardner, M.G.; Marklund, M.H.K.; Dormontt, E.E.; Lowe, A.J. Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics. Heredity 2015, 115, 108–114. [Google Scholar] [CrossRef] [Green Version]
- DiBattista, J.D. Patterns of genetic variation in anthropogenically impacted populations. Conserv. Genet. 2008, 9, 141–156. [Google Scholar] [CrossRef]
- Fontúrbel, F.E.; Bruford, M.W.; Salazar, D.A.; Cortés-Miranda, J.; Vega-Retter, C. The hidden costs of living in a transformed habitat: Ecological and evolutionary consequences in a tripartite mutualistic system with a keystone mistletoe. Sci. Total Environ. 2019, 651, 2740–2748. [Google Scholar] [CrossRef] [Green Version]
- González-Varo, J.P.; Carvalho, C.S.; Arroyo, J.M.; Jordano, P. Unravelling seed dispersal through fragmented landscapes: Frugivore species operate unevenly as mobile links. Mol. Ecol. 2017, 26, 4309–4321. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Luo, X.; Han, L.; Yang, J.; Jin, J.; Yang, J. Genetic patterns reveal differences between the invasion processes of common ragweed in urban and non-urban ecosystems. Glob. Ecol. Conserv. 2022, 38, e02214. [Google Scholar] [CrossRef]
- García-Verdugo, C.; Calleja, J.A.; Vargas, P.; Silva, L.; Moreira, O.; Pulido, F. Polyploidy and microsatellite variation in the relict tree Prunus lusitanica L.: How effective are refugia in preserving genotypic diversity of clonal taxa? Mol. Ecol. 2013, 22, 1546–1557. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.A.; Ni, Z.; Scheen, A.-C.; Koh, J.; Gilbert, C.A.; Lefkowitz, D.; Chen, Z.J.; Soltis, P.S.; Soltis, D.E. Evolution and expression of homeologous loci in Tragopogon miscellus (Asteraceae), a recent and reciprocally formed allopolyploid. Genetics 2006, 173, 1599–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, A.; Ainouche, M.L.; Wendel, J.F. Genetic and epigenetic consequences of recent hybridization and polyploidy in Spartina (Poaceae). Mol. Ecol. 2005, 14, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Parisod, C.; Salmon, A.; Zerjal, T.; Tenaillon, M.; Grandbastien, M.-A.; Ainouche, M. Rapid structural and epigenetic reorganization near transposable elements in hybrid and allopolyploid genomes in Spartina. New Phytol. 2009, 184, 1003–1015. [Google Scholar] [CrossRef]
- Chen, Z.J. Genetic and epigenetic mechanisms for gene expression and phenotypic variation in plant polyploids. Annu. Rev. Plant Biol. 2007, 58, 377–406. [Google Scholar] [CrossRef] [Green Version]
- Baker, H.G. Characteristics and modes of origin of weeds. In Characteristics and Modes of Origin of Weeds; Academic Press Inc.: New York, NY, USA, 1965; pp. 147–172. [Google Scholar]
- Levin, D.A. Mating system shifts on the trailing edge. Ann. Bot. 2012, 109, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Herrera, I.; Nassar, J.M. Reproductive and recruitment traits as indicators of the invasive potential of Kalanchoe daigremontiana (Crassulaceae) and Stapelia gigantea (Apocynaceae) in a Neotropical arid zone. J. Arid Environ. 2009, 73, 978–986. [Google Scholar] [CrossRef]
- Lemus-Jiménez, L.J.; Ramírez, N. Breeding systems of the Paraguaná coastal plain plants, Venezuela. Rev. Biol. Trop. 2005, 53, 415–430. [Google Scholar] [CrossRef]
- Duminil, J.; Hardy, O.J.; Petit, R.J. Plant traits correlated with generation time directly affect inbreeding depression and mating system and indirectly genetic structure. BMC Evol. Biol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Dick, C.W.; Bermingham, E.; Lemes, M.R.; Gribel, R. Extreme long-distance dispersal of the lowland tropical rainforest tree Ceiba pentandra L. (Malvaceae) in Africa and the Neotropics. Mol. Ecol. 2007, 16, 3039–3049. [Google Scholar] [CrossRef]
- Lobo, J.A.; Quesada, M.; Stoner, K.E. Effects of pollination by bats on the mating system of Ceiba pentandra (Bombacaceae) populations in two tropical life zones in Costa Rica. Am. J. Bot. 2005, 92, 370–376. [Google Scholar] [CrossRef]
- Mullarkey, A.A.; Byers, D.L.; Anderson, R.C. Inbreeding depression and partitioning of genetic load in the invasive biennial Alliaria petiolata (Brassicaceae). Am. J. Bot. 2013, 100, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Durka, W.; Bossdorf, O.; Prati, D.; Auge, H. Molecular evidence for multiple introductions of garlic mustard (Alliaria petiolata, Brassicaceae) to North America. Mol. Ecol. 2005, 14, 1697–1706. [Google Scholar] [CrossRef]
- Ross, C.A.; Auge, H.; Durka, W. Genetic relationships among three native North-American Mahonia species, invasive Mahonia populations from Europe, and commercial cultivars. Plant Syst. Evol. 2008, 275, 219–229. [Google Scholar] [CrossRef]
- Lachmuth, S.; Durka, W.; Schurr, F.M. The making of a rapid plant invader: Genetic diversity and differentiation in the native and invaded range of Senecio inaequidens. Mol. Ecol. 2010, 19, 3952–3967. [Google Scholar] [CrossRef]
- Pairon, M.; Petitpierre, B.; Campbell, M.; Guisan, A.; Broennimann, O.; Baret, P.V.; Jacquemart, A.L.; Besnard, G. Multiple introductions boosted genetic diversity in the invasive range of black cherry (Prunus serotina; Rosaceae). Ann. Bot. 2010, 105, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.-Y.; Hong, L.; Wang, Z.-F.; Shen, H.; Ye, W.-H.; Mu, H.-P.; Cao, H.-L.; Wang, Z.-M.; Bradshaw, C.J.A. Inferring the invasion history of coral berry Ardisia crenata from China to the USA using molecular markers. Ecol. Res. 2012, 27, 809–818. [Google Scholar] [CrossRef]
- Kelager, A.; Pedersen, J.S.; Bruun, H.H. Multiple introductions and no loss of genetic diversity: Invasion history of Japanese Rose, Rosa rugosa, in Europe. Biol. Invasions 2013, 15, 1125–1141. [Google Scholar] [CrossRef]
- Molins, M.P.; Corral, J.M.; Aliyu, O.M.; Koch, M.A.; Betzin, A.; Maron, J.L.; Sharbel, T.F. Biogeographic variation in genetic variability, apomixis expression and ploidy of St. John’s wort (Hypericum perforatum) across its native and introduced range. Ann. Bot. 2014, 113, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.L.; Fishman, L. Genetic evidence for founder effects in the introduced range of houndstongue (Cynoglossum officinale). Biol. Invasions 2014, 16, 205–216. [Google Scholar] [CrossRef]
- Kang, M.; Buckley, Y.M.; Lowe, A.J. Testing the role of genetic factors across multiple independent invasions of the shrub Scotch broom (Cytisus scoparius). Mol. Ecol. 2007, 16, 4662–4673. [Google Scholar] [CrossRef] [PubMed]
- Prentis, P.J.; Sigg, D.P.; Raghu, S.; Dhileepan, K.; Pavasovic, A.; Lowe, A.J. Understanding invasion history: Genetic structure and diversity of two globally invasive plants and implications for their management. Divers. Distrib. 2009, 15, 822–830. [Google Scholar] [CrossRef]
- Gaudeul, M.; Giraud, T.; Kiss, L.; Shykoff, J.A. Nuclear and chloroplast microsatellites show multiple introductions in the worldwide invasion history of common ragweed, Ambrosia artemisiifolia. PLoS ONE 2011, 6, e17658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornoy, B.; Atlan, A.; Roussel, V.; Buckley, Y.M.; Tarayre, M. Two colonisation stages generate two different patterns of genetic diversity within native and invasive ranges of Ulex europaeus. Heredity 2013, 111, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Mandak, B.; Hadincová, V.; Mahelka, V.; Wildova, R. European invasion of North American Pinus strobus at large and fine scales: High genetic diversity and fine-scale genetic clustering over time in the adventive range. PLoS ONE 2013, 8, e68514. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Solís-Montero, L.; Lou, A.; Vallejo-Marín, M. Population structure and genetic diversity of native and invasive populations of Solanum rostratum (Solanaceae). PLoS ONE 2013, 8, e79807. [Google Scholar] [CrossRef] [Green Version]
- Rijal, D.P.; Alm, T.; Jahodová, Š.; Stenøien, H.K.; Alsos, I.G. Reconstructing the invasion history of Heracleum persicum (Apiaceae) into Europe. Mol. Ecol. 2015, 24, 5522–5543. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.Y.; Tang, S.; Fatemi, M.; Gross, C.L.; Julien, M.H.; Curtis, C.; Van Klinken, R.D. Population structure and genetic diversity of invasive Phyla canescens: Implications for the evolutionary potential. Ecosphere 2015, 6, 1–21. [Google Scholar] [CrossRef]
- Mukherjee, A.; Williams, D.; Gitzendanner, M.A.; Overholt, W.A.; Cuda, J.P. Microsatellite and chloroplast DNA diversity of the invasive aquatic weed Hygrophila polysperma in native and invasive ranges. Aquat. Bot. 2016, 129, 55–61. [Google Scholar] [CrossRef]
- Thompson, G.D.; Richardson, D.M.; Wilson, J.R.U.; Bellstedt, D.U.; Le Roux, J.J. Genetic diversity and structure of the globally invasive tree, Paraserianthes lophantha subspecies lophantha, suggest an introduction history characterised by varying propagule pressure. Tree Genet. Genomes 2016, 12, 1–9. [Google Scholar] [CrossRef]
- Závada, T.; Malik, R.J.; Kesseli, R. V Population structure in chicory (Cichorium intybus): A successful US weed since the American revolutionary war. Ecol. Evol. 2017, 7, 4209–4219. [Google Scholar] [CrossRef]
- Xia, L.; Geng, Q.; An, S. Rapid genetic divergence of an invasive species, Spartina alterniflora, in China. Front. Genet. 2020, 11, 284. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.; Richardson, D.M.; Pauchard, A.; Le Roux, J.J. Genetic analyses reveal complex introduction histories for the invasive tree Acacia dealbata Link around the world. Divers. Distrib. 2021, 27, 360–376. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Description |
|---|---|
| Habitat fragmentation | Continuous areas subdivided into smaller fragments that increase habitat isolation and edge effects, usually accompanied by habitat loss. |
| Habitat degradation | Areas with a reduction in habitat quality due to changes in vegetation structure, resource availability, and microclimate conditions but without experiencing fragmentation or loss. |
| Undisturbed habitat | Areas unaltered by humans. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Espinosa, R.; González-Astorga, J.; Rico, Y.; Gallego-Fernández, J.B. Effect of Life-History Traits and Habitat Condition on Genetic Diversity between Invasive and Native Plant Populations. Diversity 2022, 14, 1025. https://doi.org/10.3390/d14121025
Hernández-Espinosa R, González-Astorga J, Rico Y, Gallego-Fernández JB. Effect of Life-History Traits and Habitat Condition on Genetic Diversity between Invasive and Native Plant Populations. Diversity. 2022; 14(12):1025. https://doi.org/10.3390/d14121025
Chicago/Turabian StyleHernández-Espinosa, Raquel, Jorge González-Astorga, Yessica Rico, and Juan B. Gallego-Fernández. 2022. "Effect of Life-History Traits and Habitat Condition on Genetic Diversity between Invasive and Native Plant Populations" Diversity 14, no. 12: 1025. https://doi.org/10.3390/d14121025