
Diversity, Seasonal and Diel Distribution Patterns of Anchovies (Osteichthyes) in a Protected Tropical Lagoon in the Southwestern Gulf of Mexico

Abstract
:1. Introduction
2. Materials and Methods
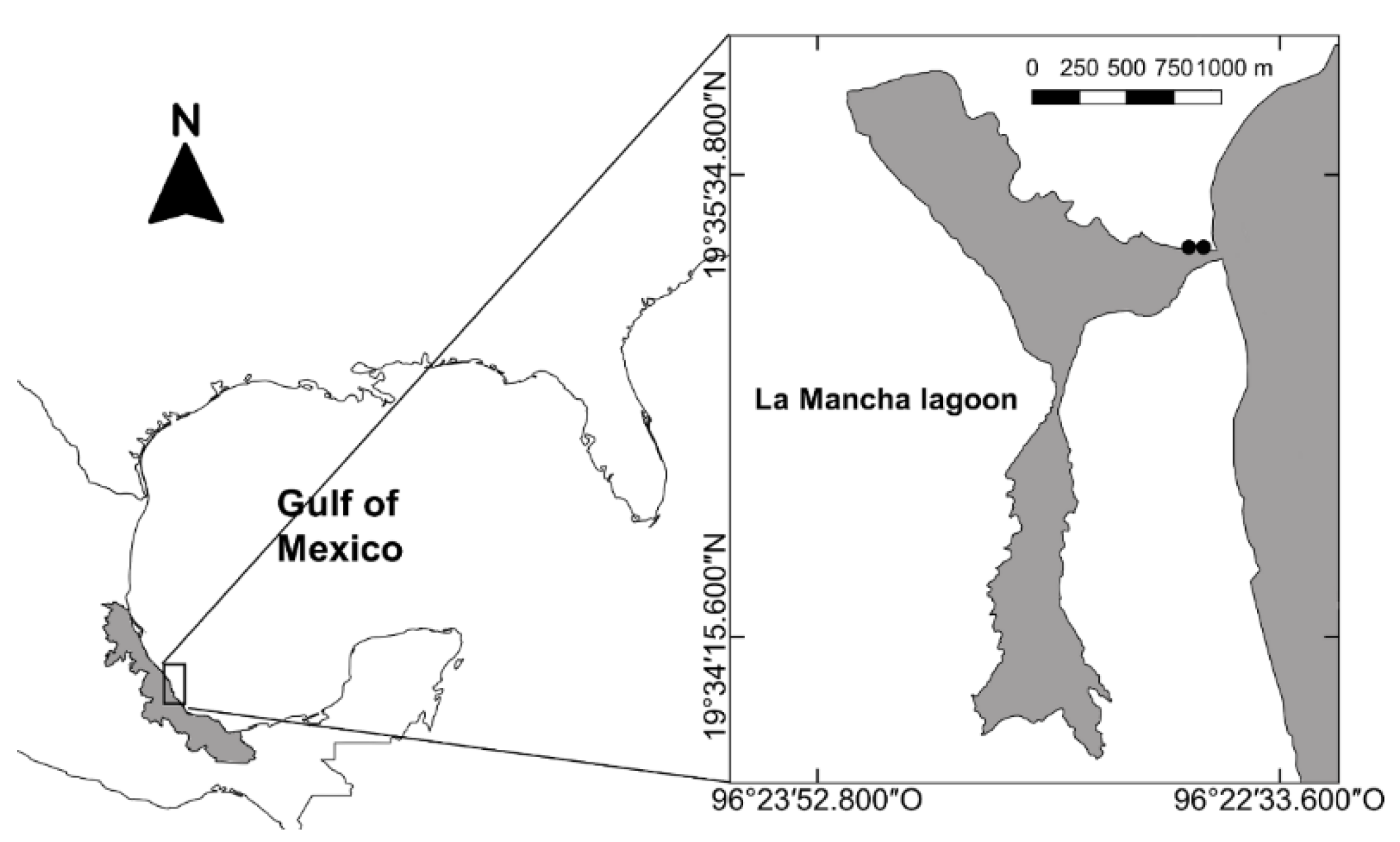
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
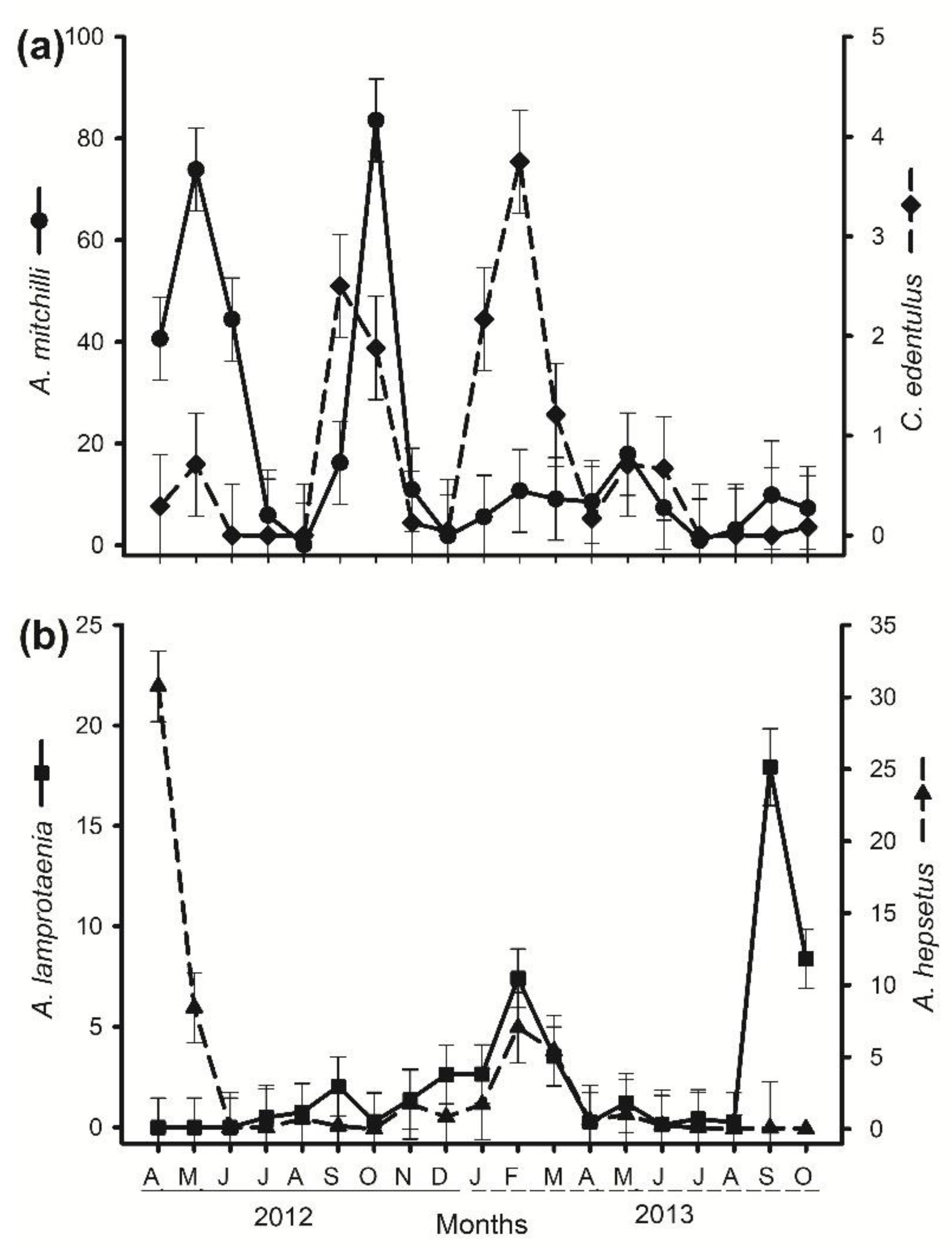
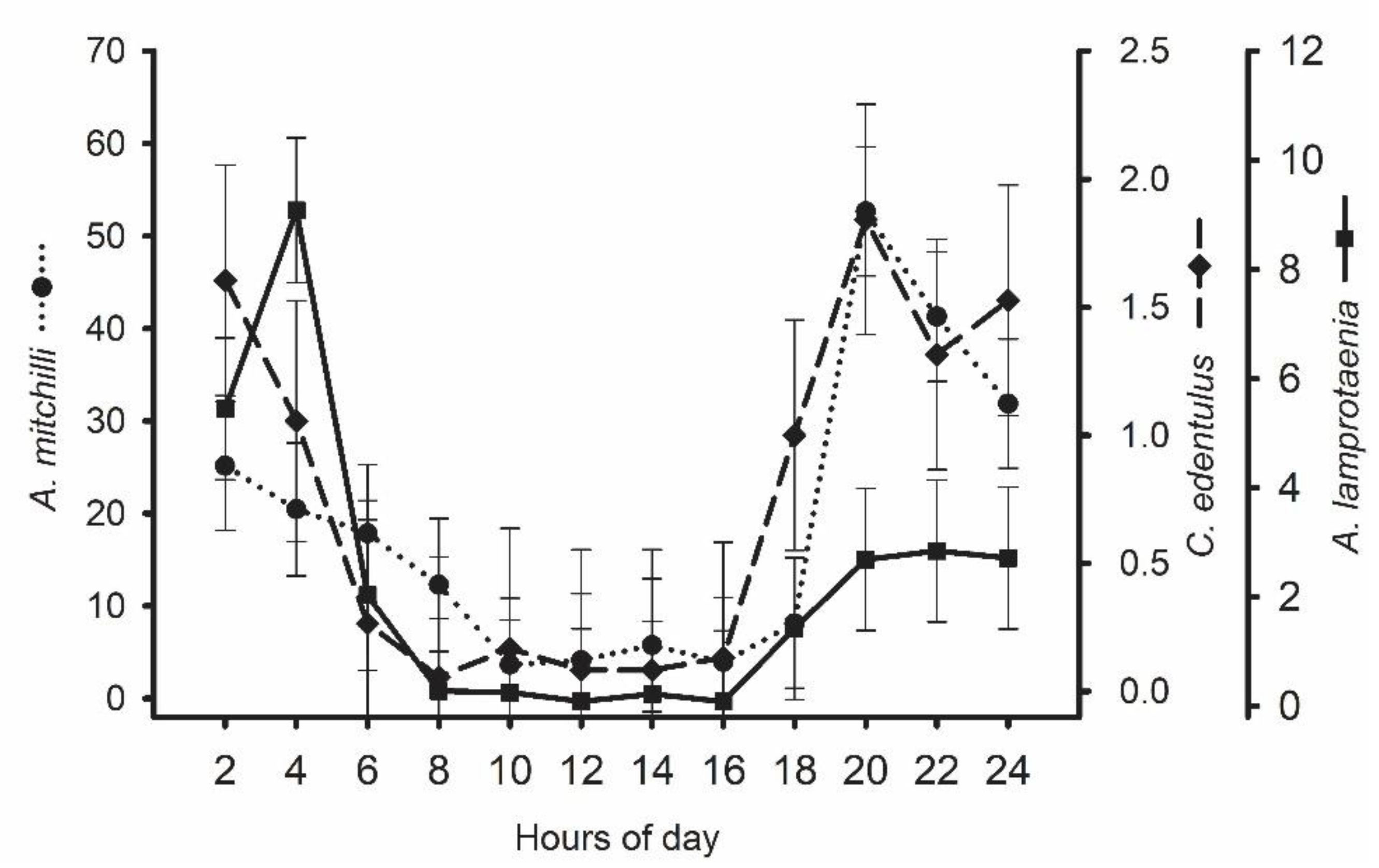
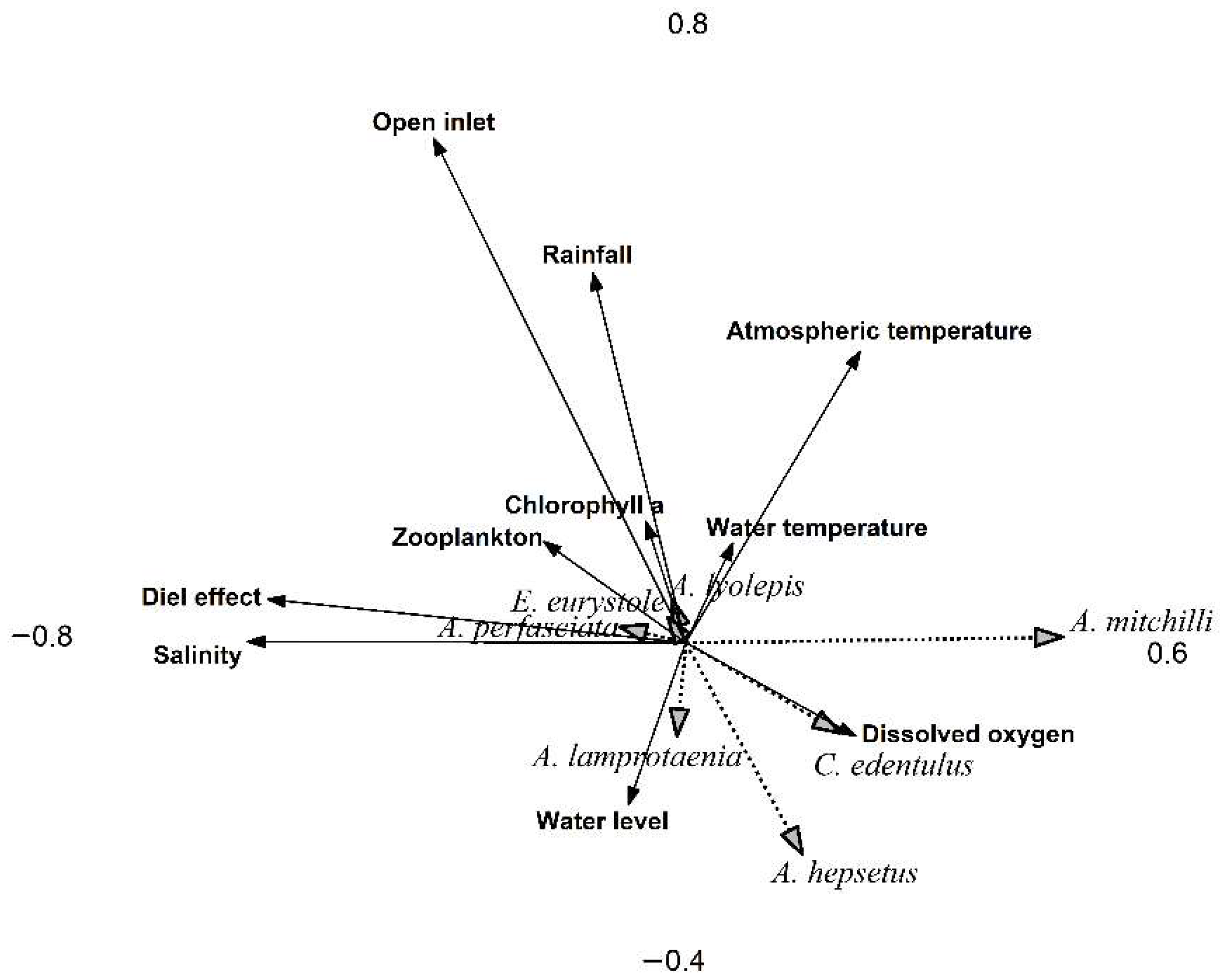
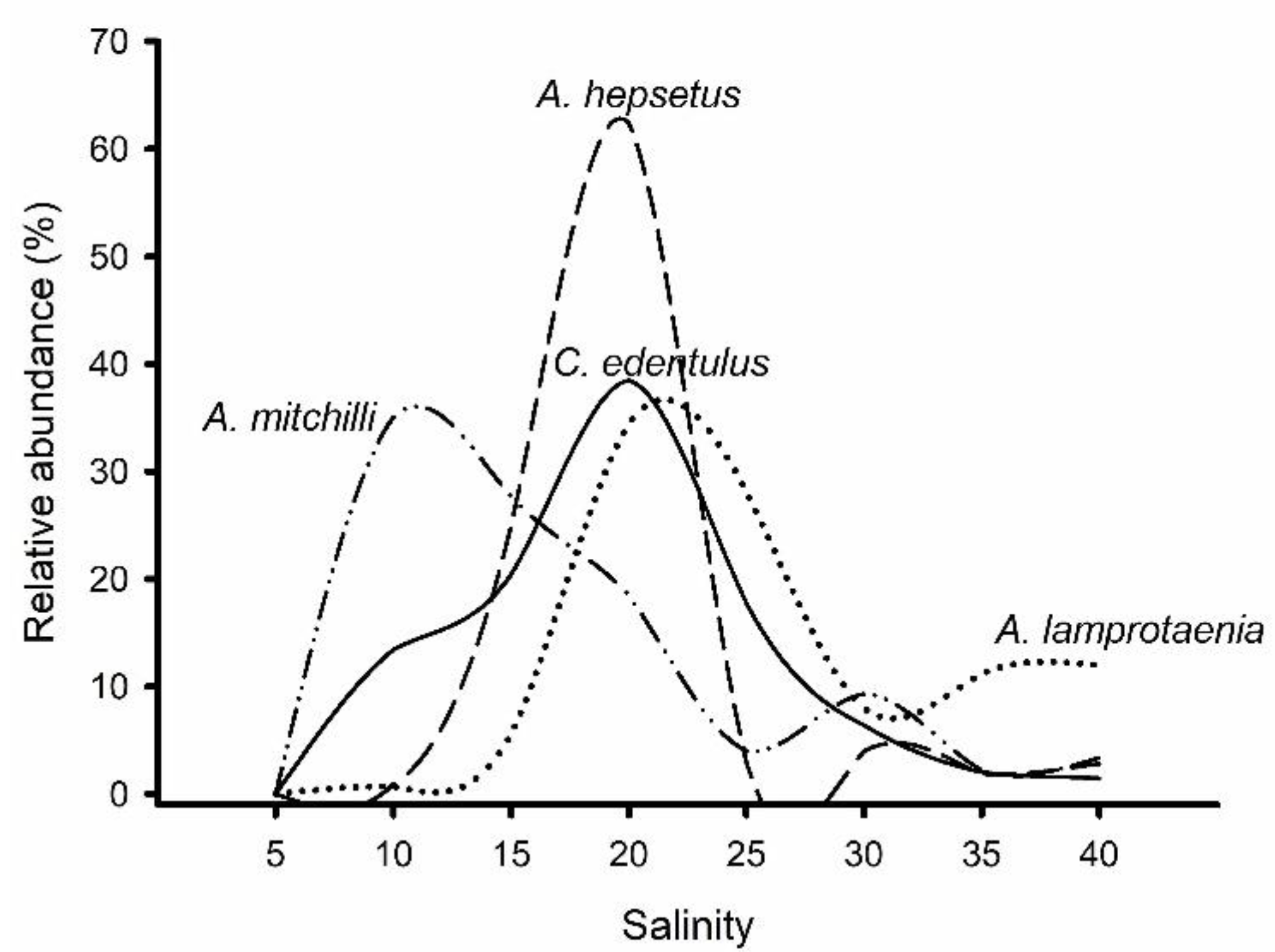
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, J.F.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; p. 752. [Google Scholar]
- Nizinski, M.; Munroe, T. Order Clupeiformes: Engraulidae. In The Living Marine Resources of The Western Central Atlantic Volume 2; Carpenter, K.E., Ed.; FAO Species Identification Guide for Fishery Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5; FAO: Rome, Italy, 2002; pp. 764–794. [Google Scholar]
- McEachran, J.D. Fishes (Vertebrata: Pisces) of the Gulf of Mexico. In Gulf of Mexico–Origins, Waters, and Biota. Biodiversity; Felder, D.L., Camp, D.K., Eds.; Texas A&M University Press: College Station, TX, USA, 2009; pp. 1223–1316. [Google Scholar]
- Lara-Domínguez, A.L.; Franco-López, J.; Bedia-Sánchez, C.; Abarca-Arenas, L.G.; Díaz-Ruiz, S.; Aguirre-León, A.; González-Gándara, C.; Castillo-Rivera, M. Diversidad de Peces en los Ambientes Costeros y Plataforma Continental. Volume II. In La Biodiversidad en Veracruz: Estudio de Estado; CONABIO, Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A.C.: Veracruz, Mexico, 2011; pp. 505–516. [Google Scholar]
- Hasan, M.R.; Halwart, M. Fish as Feed Inputs for Aquaculture. Practices, Sustainability and Implications; FAO Technical Paper 518; FAO: Rome, Italy, 2009; p. 426. [Google Scholar]
- Castillo-Rivera, M.; Moreno, G.; Iniestra, R. Spatial, seasonal and diel variation in abundance of the bay anchovy, Anchoa mitchilli (Teleostei: Engraulidae), in a tropical coastal lagoon on Mexico. Southwest Nat. 1994, 39, 263–268. [Google Scholar] [CrossRef]
- Chen, Y. Fish resources of the Gulf of Mexico. In Habitats and Biota of the Gulf of Mexico: Before the Deepwater Horizon Oil Spill; Ward, C.H., Ed.; Springer Nature: New York, NY, USA, 2017; Volume II, pp. 869–1038. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, X.; Yang, Q.; Zuo, T.; Quigg, A. Mesozooplankton dynamics in relation to environmental factors and juvenile fish in a subtropical estuary of the Gulf of Mexico. J. Coast. Res. 2017, 33, 1038–1050. [Google Scholar] [CrossRef]
- Wang, S.; Houde, E.D. Distribution, relative abundance, biomass and production of bay anchovy Anchoa mitchilli in the Chesapeake Bay. Mar. Ecol. Prog. Ser. 1995, 121, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Araújo, F.G.; Silva, M.A.; Santos, J.N.S.; Vasconcellos, R.M. Habitat selection by anchovies (Clupeiformes: Engraulidae) in a tropical bay at Southeastern Brazil. Neotrop. Ichthyol. 2008, 6, 583–590. [Google Scholar] [CrossRef]
- Hagan, S.M.; Able, K.W. Diel variation in the pelagic fish assemblage in a temperate estuary. Estuaries Coast. 2008, 31, 33–42. [Google Scholar] [CrossRef]
- Castillo-Rivera, M.; Ortiz-Burgos, S.; Zárate-Hernández, H. Temporal changes in species richness and fish composition in a submerged vegetation habitat in Veracruz, Mexico. Acta Ichthyol. Piscat. 2017, 47, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Pichler, H.A.; Gray, C.A.; Broadhurst, M.K.; Spach, H.L.; Nagelkerken, I. Seasonal and environmental influences on recruitment patterns and habitat usage among resident and transient fishes in a World Heritage Site subtropical estuary. J. Fish Biol. 2017, 90, 396–416. [Google Scholar] [CrossRef]
- Barletta, M.; Amaral, C.; Corrêa, M.F.; Guebert, F.M.; Dantas, D.V.; Lorenzi, L.; Saint-Paul, U. Factors affecting seasonal variations in demersal fish assemblages at an ecocline in a tropical–subtropical estuary. J. Fish Biol. 2008, 73, 1314–1336. [Google Scholar] [CrossRef]
- Whitfield, A.K. Estuaries—How challenging are these constantly changing aquatic environments for associated fish species? Environ. Biol. Fishes 2021, 104, 517–528. [Google Scholar] [CrossRef]
- Pessanha, A.L.M.; Araújo, F.G.; De Azevedo, M.C.C.; Dias Gomes, D. Diel and seasonal changes in the distribution of fish on southeast Brazil sandy beach. Mar. Biol. 2003, 143, 1047–1055. [Google Scholar] [CrossRef]
- Garcia, A.F.; Pessanha, A.L. Temporal dynamics of tidal mudflat fish assemblages in north-eastern Brazil: Short and medium-term variations. J. Mar. Biol. Assoc. UK 2018, 98, 1745–1755. [Google Scholar] [CrossRef]
- Castillo-Rivera, M.; Zárate-Hernández, R.; Ortiz-Burgos, S.; Zavala-Hurtado, A. Diel and seasonal variability in the fish community structure of a mud-bottom estuarine habitat in the Gulf of Mexico. Mar. Ecol. 2010, 31, 633–642. [Google Scholar] [CrossRef]
- Lacerda, C.H.F.; Barletta, M.; Dantas, D.V. Temporal patterns in the intertidal faunal community at the mouth of a tropical estuary. J. Fish Biol. 2014, 85, 1571–1602. [Google Scholar] [CrossRef]
- Ribeiro, F.; Hale, E.; Hilton, E.J.; Clardy, T.R.; Deary, A.L.; Targett, T.E.; Olney, J.E. Composition and temporal patterns of larval fish communities in Chesapeake and Delaware Bays, USA. Mar. Ecol. Prog. Ser. 2015, 527, 167–180. [Google Scholar] [CrossRef]
- Auth, T.D.; Arula, T.; Houde, E.D.; Woodland, R.J. Spatial ecology and growth in early life stages of bay anchoy Anchoa mitchilli in Chesapeake Bay (USA). Mar. Ecol. Prog. Ser. 2020, 651, 125–143. [Google Scholar] [CrossRef]
- Santos, S.R.; Galvão, K.P.; Adler, G.H.; Andrade-Tubino, M.F.; Vianna, M. Spatiotemporal distribution and population biology aspects of Cetengraulis edentulus (Actinopterygii: Clupeiformes: Engraulidae) in a South-western Atlantic estuary, with notes on the local Clupeiformes community: Conservation implications. Acta Ichthyol. Piscat. 2020, 50, 139–150. [Google Scholar] [CrossRef]
- Silva, M.A.; Araújo, F.G.; Costa, M.C.; Santos, J. The nursery function of sandy beaches in a Brazilian tropical bay for 0-group anchovies (Teleostei, Engraulidae): Diel, seasonal and spatial patterns. J. Mar. Biol. Assoc. UK 2004, 84, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Google. (s.f.). Mapa de Veracruz, Mexico en Google Maps. Available online: https://www.google.com.co/maps/@4.6315748,-74.0699088,11.79z?hl=en (accessed on 3 December 2018).
- QGIS.org. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: https://qgis.org (accessed on 3 December 2018).
- Whitehead, P.J.P.; Nelson, G.; Wongratana, T. FAO Species Catalogue. Vol. 7. Clupeoid Fishes of the World (Suborder Clupeoidei). An Annotated and Illustrated Catalogue of the Herrings, Sardines, Pilchards, Sprats, Anchovies and Wolf-Herrings. Part 2. Engraulididae; FAO: Rome, Italy, 1988; Volume 7, pp. 305–579. [Google Scholar]
- Servicio Meteorológico Nacional (SMN). Normales Climatológicas por Estado. Available online: https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado?estado=ver (accessed on 17 March 2020).
- Constante-Pérez, L.G.; Castillo-Rivera, M.; Serrato-Díaz, A. Diversity, seasonal and diel distribution of snappers (Lutjanidae: Perciformes) in a tropical coastal inlet in the southwestern Gulf of Mexico. Neotrop. Ichthyol. 2022, 20, e220009. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; Massey University: Palmerston North, New Zealand, 2008; p. 214. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v7: User Manual/Tutorial; PRIMER-E: Plymouth, MA, USA, 2015; p. 296. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User´s Guide: Software for Canonical Community Ordination; Version 4.5; Microcomputer Power: Ithaca, NY, USA, 2002; p. 500. [Google Scholar]
- Helfman, G.S. Fish Behaviour by Day, Night and Twilight. In Behaviour of Teleost Fishes, 2nd ed.; Pitcher, T.J., Ed.; Chapman and Hall: London, UK, 1993; pp. 479–512. [Google Scholar]
- Torre, M.P.; Lifavi, D.M.; Targett, T.E. Diel differences in abundance and diversity of fish species and blue crab (Callinectes sapidus) in the sandy beach shore zone of lower Delaware Bay. Fish. Bull. 2017, 115, 556–565. [Google Scholar] [CrossRef]
- Nemerson, D.M.; Able, K.W. Diel and tidal influences on the abundance and food habits of four young-of-the-year fish in Delaware Bay, USA, marsh creeks. Environ. Biol. Fishes 2020, 103, 251–268. [Google Scholar] [CrossRef]
- Castillo-Rivera, M.; Kobelkowsky, A. Distribution and segregation of two sympatric Brevoortia species (Teleostei: Clupeidae). Estuaries Coast. Shelf Sci. 2000, 50, 593–598. [Google Scholar] [CrossRef]
- Kimura, R.; Secor, D.H.; Houde, E.D.; Piccoli, P.M. Up-estuary dispersal of young-of-the-year bay anchovy Anchoa mitchilli in the Chesapeake Bay: Inferences from microprobe analysis of strontium in otoliths. Mar. Ecol. Prog. Ser. 2000, 208, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Peebles, E.B.; Burghart, S.E.; Hollander, D.J. Causes of interestuarine variability in bay anchovy (Anchoa mitchilli) salinity at capture. Estuaries Coast. 2007, 30, 1060–1074. [Google Scholar] [CrossRef]
- James, N.C.; Cowley, P.D.; Whitfield, A.K.; Lamberth, S.J. Fish communities in temporarily open/closed estuaries from warm and cool-temperate regions of South Africa: A review. Rev. Fish Biol. Fish. 2007, 17, 565–580. [Google Scholar] [CrossRef]
- Whitfield, A.K.; Adams, J.B.; Bate, G.C.; Bezuidenhout, K.; Bornman, T.G.; Cowley, P.D.; Froneman, P.W.; Gama, P.T.; James, N.C.; Mackenzie, B.; et al. A multidisciplinary study of a small, temporarily open/closed South African estuary. Afr. J. Mar. Sci. 2008, 30, 453–473. [Google Scholar] [CrossRef]
- Mendoza, E.; Castillo-Rivera, M.; Zárate-Hernández, R.; Ortiz Burgos, S. Seasonal variations in the diversity, abundance, and composition of species in an estuarine fish community in the Tropical Eastern Pacific, Mexico. Ichthyol. Res. 2009, 56, 330–339. [Google Scholar] [CrossRef]
- Becker, A.; Lowry, M.B.; Taylor, M.D. Scales of spatial and temporal variation of small bodied nekton within intermittently closed/open lakes and lagoons (ICOLLs) in south-eastern Australia. Reg. Stud. Mar. Sci. 2020, 33, 100936. [Google Scholar] [CrossRef]
- Mecalco-Hernández, A.; Castillo-Rivera, M.; Sanvicente-Añorve, L.; Flores-Coto, C.; Álvarez-Silva, C. Variación estacional y nictímera en la distribución del zooplancton dominante en una laguna costera tropical. Rev. Biol. Mar. Oceanogr. 2018, 53, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Rivera, M. Influence of rainfall pattern in the seasonal variation of fish abundance in a tropical estuary with restricted marine communication. J. Water Resour. Prot. 2013, 5, 311–319. [Google Scholar] [CrossRef]
- Souza, C.D.; Batista, V.S.; Fabré, N.N. What are the main local drivers determining richness and fishery yields in tropical coastal fish assemblages? Zoologia. 2018, 35, e12898. [Google Scholar] [CrossRef] [Green Version]
- Peebles, E.B. Temporal resolution of biological and physical influences on bay anchovy Anchoa mitchilli egg abundance near a river-plume frontal zone. Mar. Ecol. Prog. Ser. 2002, 237, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Oghenekaro, E.U.; Chigbu, P.; Oseji, O.F.; Tang, K.W. Seasonal Factors Influencing Copepod Abundance in the Maryland Coastal Bays. Estuaries Coast. 2018, 4, 495–506. [Google Scholar] [CrossRef]
- Ellis, J.T.; Dean, B.J. Gulf of Mexico Processes. J. Coast. Res. 2012, Special Issue No. 60, 6–13. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-1. 2022. Available online: https://www.iucnredlist.org (accessed on 29 September 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | df | MS | Pseudo-F | P (Permuted) |
|---|---|---|---|---|
| Anchoa mitchilli | ||||
| Months | 18 | 14,046 | 8.405 | 0.0001 |
| Diel | 1 | 39,377 | 23.563 | 0.0001 |
| Months x Diel | 18 | 1360.2 | −0.81396 | 0.0959 |
| Residual | 404 | 1671.1 | ||
| Anchoa hepsetus | ||||
| Months | 18 | 1220.3 | 6.8875 | 0.0001 |
| Diel | 1 | 208.57 | 11772 | 0.2104 |
| Months x Diel | 18 | 219.08 | −1.2365 | 0.2347 |
| Residual | 404 | 177.18 | ||
| Anchoa lamprotaenia | ||||
| Months | 18 | 336.83 | 5.8138 | 0.0001 |
| Diel | 1 | 878.04 | 15.155 | 0.0001 |
| Months x Diel | 18 | 64.768 | −1.1179 | 0.2749 |
| Residual | 404 | 57.937 | ||
| Cetengraulis edentulus | ||||
| Months | 18 | 27.303 | 4.4635 | 0.0001 |
| Diel | 1 | 87.311 | 14.274 | 0.0005 |
| Months x Diel | 18 | 5.1598 | 0.84353 | 0.0222 |
| Residual | 404 | 6.1169 |
| Correlations | Axis 1 | Axis 2 | P | VIF |
|---|---|---|---|---|
| Salinity | −0.300 | 0.0006 | 0.002 | 1.911 |
| Diel effect | −0.286 | 0.019 | 0.002 | 1.035 |
| Open inlet | −0.172 | 0.221 | 0.026 | 2.707 |
| Atmospheric temperature | 0.119 | 0.127 | 0.002 | 2.143 |
| Rainfall | −0.063 | 0.162 | 0.014 | 3.142 |
| Water temperature | 0.032 | 0.043 | 0.030 | 2.395 |
| Zooplankton | −0.097 | 0.044 | 0.392 | 1.257 |
| Chlorophyll a | −0.027 | 0.053 | 0.258 | 1.288 |
| Dissolved oxygen | 0.116 | −0.041 | 0.162 | 2.368 |
| Water level | −0.039 | −0.070 | 0.750 | 1.072 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgado-Dueñas, G.; Castillo-Rivera, M. Diversity, Seasonal and Diel Distribution Patterns of Anchovies (Osteichthyes) in a Protected Tropical Lagoon in the Southwestern Gulf of Mexico. Diversity 2022, 14, 852. https://doi.org/10.3390/d14100852
Morgado-Dueñas G, Castillo-Rivera M. Diversity, Seasonal and Diel Distribution Patterns of Anchovies (Osteichthyes) in a Protected Tropical Lagoon in the Southwestern Gulf of Mexico. Diversity. 2022; 14(10):852. https://doi.org/10.3390/d14100852
Chicago/Turabian StyleMorgado-Dueñas, Guadalupe, and Manuel Castillo-Rivera. 2022. "Diversity, Seasonal and Diel Distribution Patterns of Anchovies (Osteichthyes) in a Protected Tropical Lagoon in the Southwestern Gulf of Mexico" Diversity 14, no. 10: 852. https://doi.org/10.3390/d14100852