

Odonata from Iberá Wetland System (Corrientes, Argentina) Are Regional Biogeographic Schemes Useful to Assess Odonata Biodiversity and Its Conservation?
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
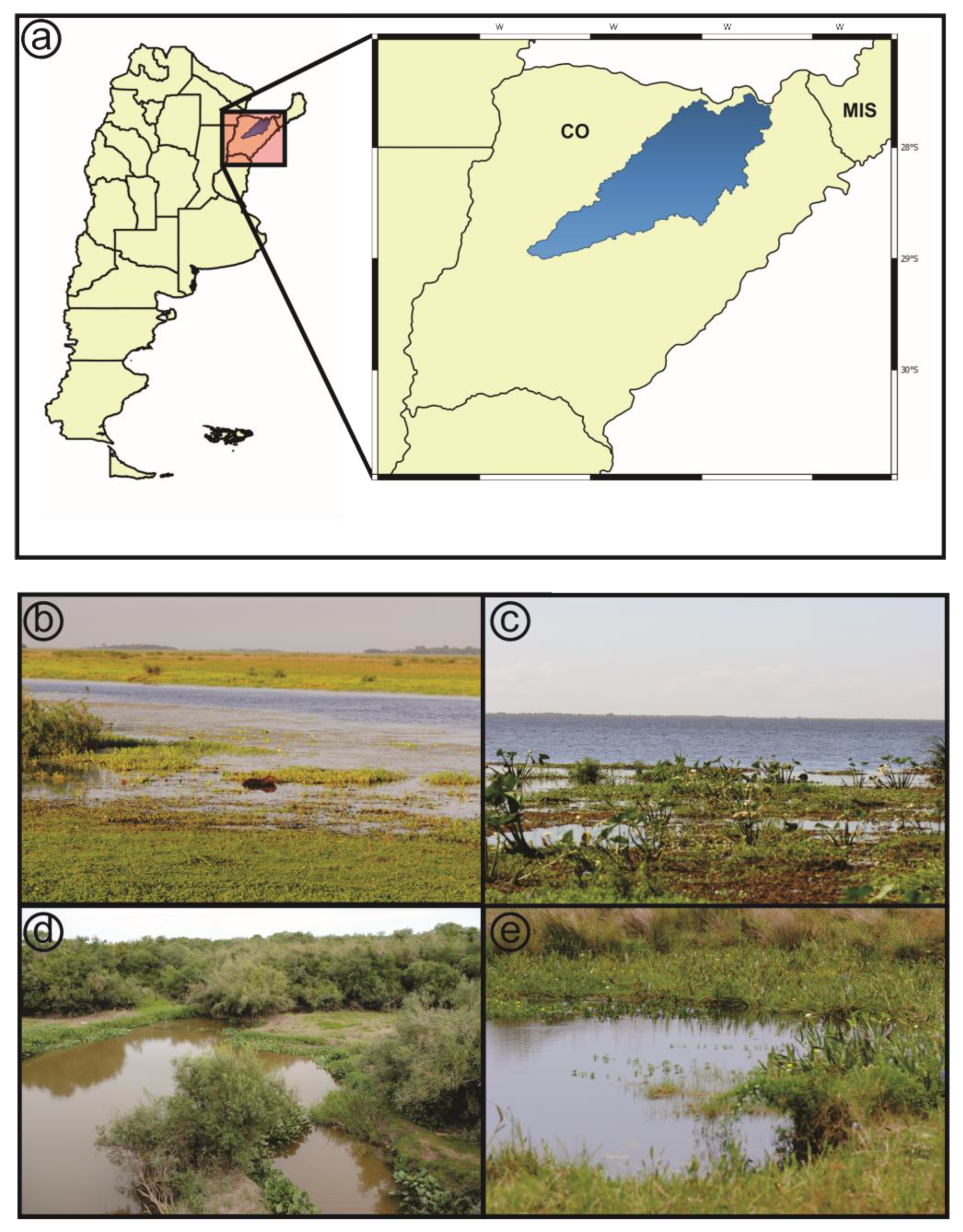
2.1. Study Area
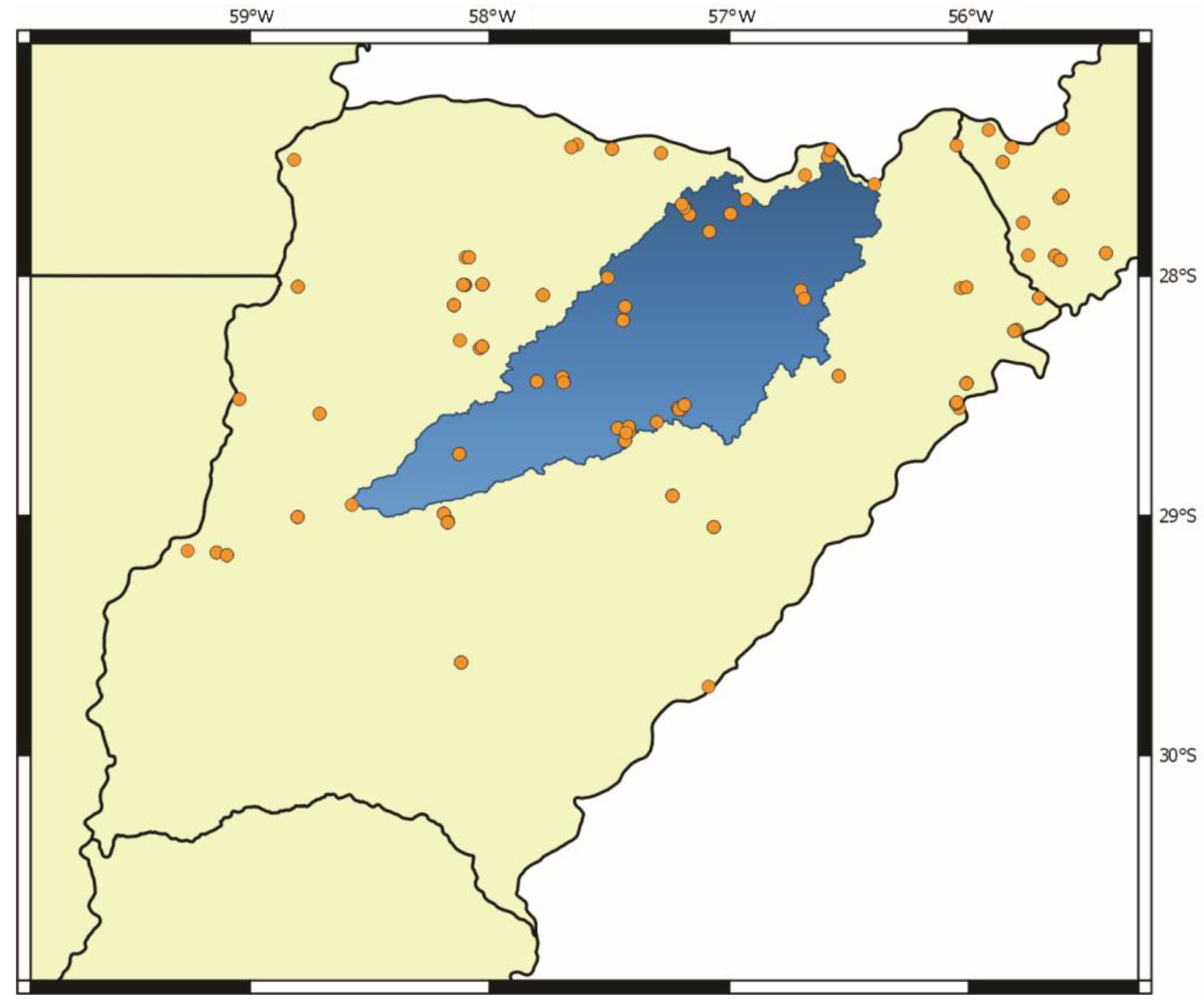
2.2. Sampling Procedures
2.3. Regionalization Schemes
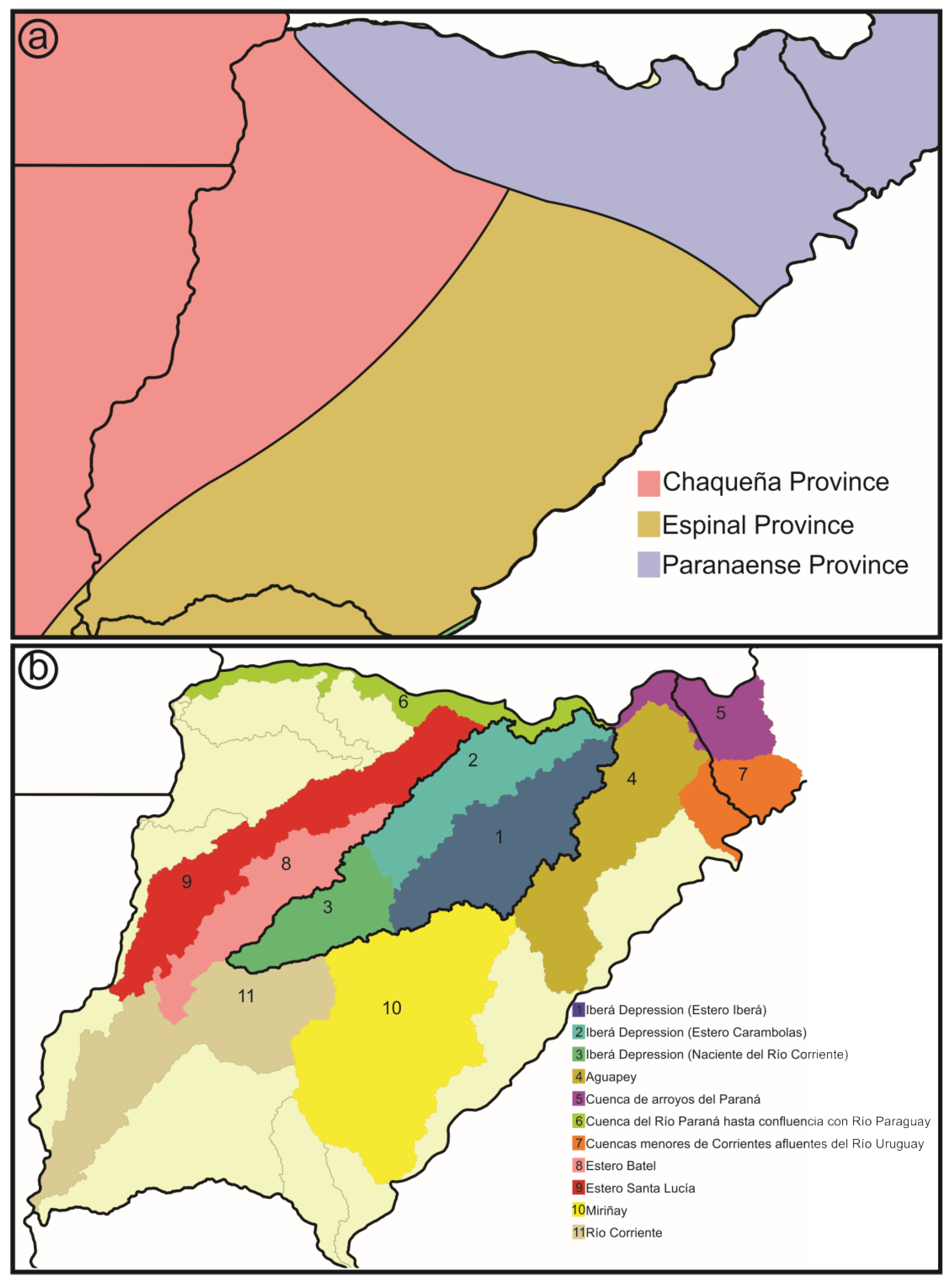
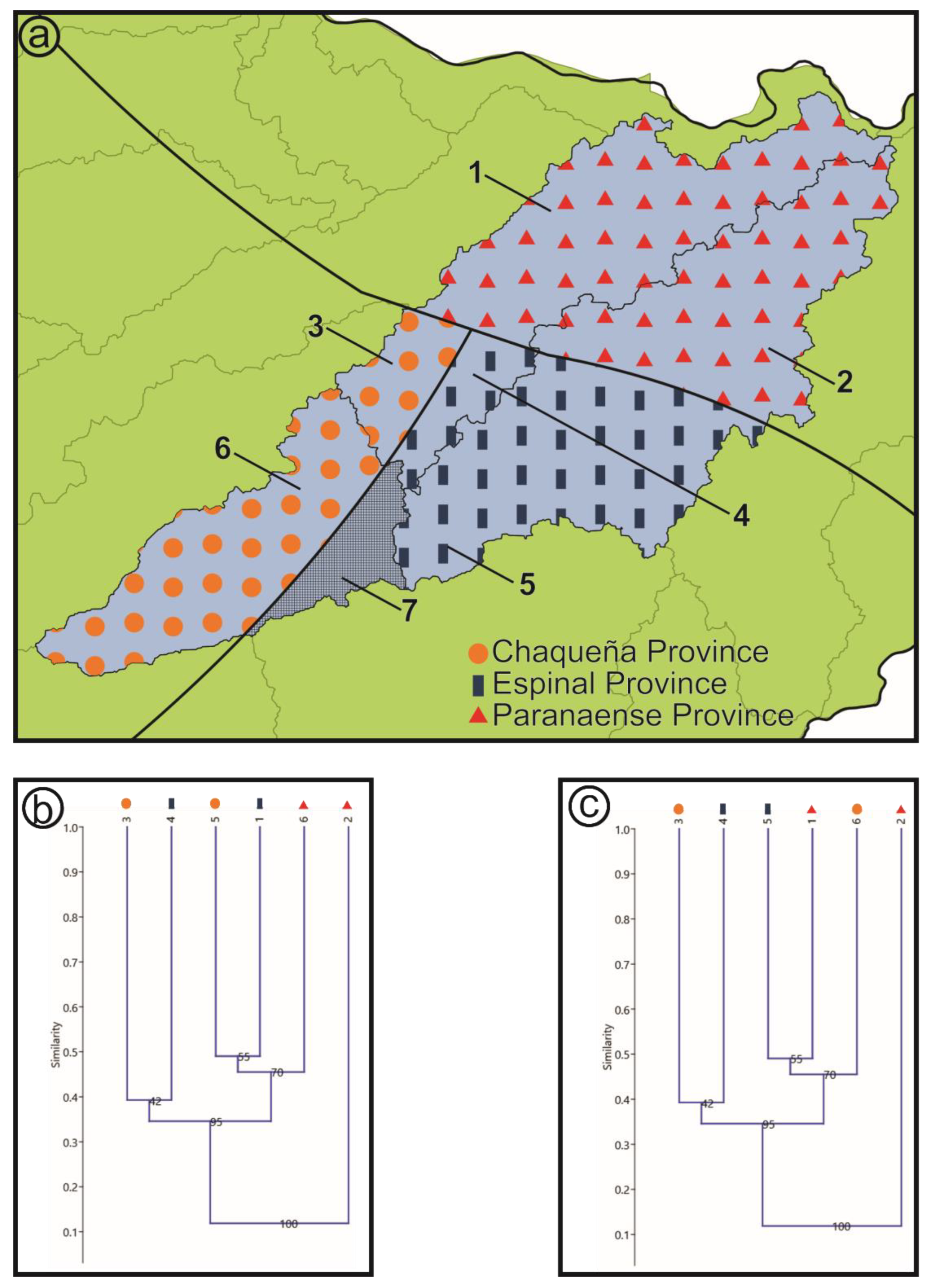
- Biogeographic provinces sensu Cabrera and Willink [17]: This scheme proposes a hierarchical partition, which is a divisive, non-agglomerative classification of the regions into domains, provinces, and districts. In this categorization, the successive levels mainly based on the phytogeographic hierarchy, from domain to district, are carried out by the presence of endemisms and predominance of families and species [18,24]. The vegetation of the Neotropical and Subantarctic sectors of Argentina is then classified into 12 provinces [24], three of them are represented in the area: Espinal, Chaqueña and Paranaense (Figure 4a).
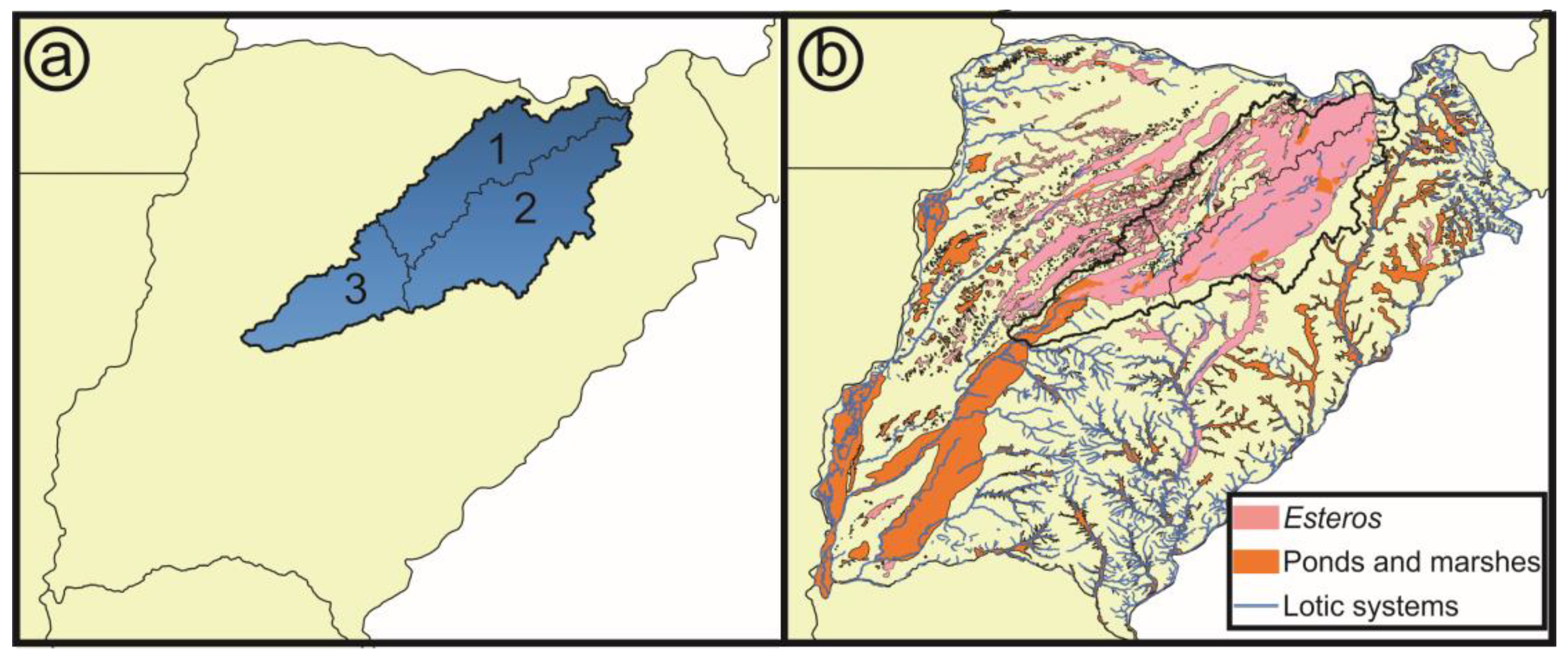
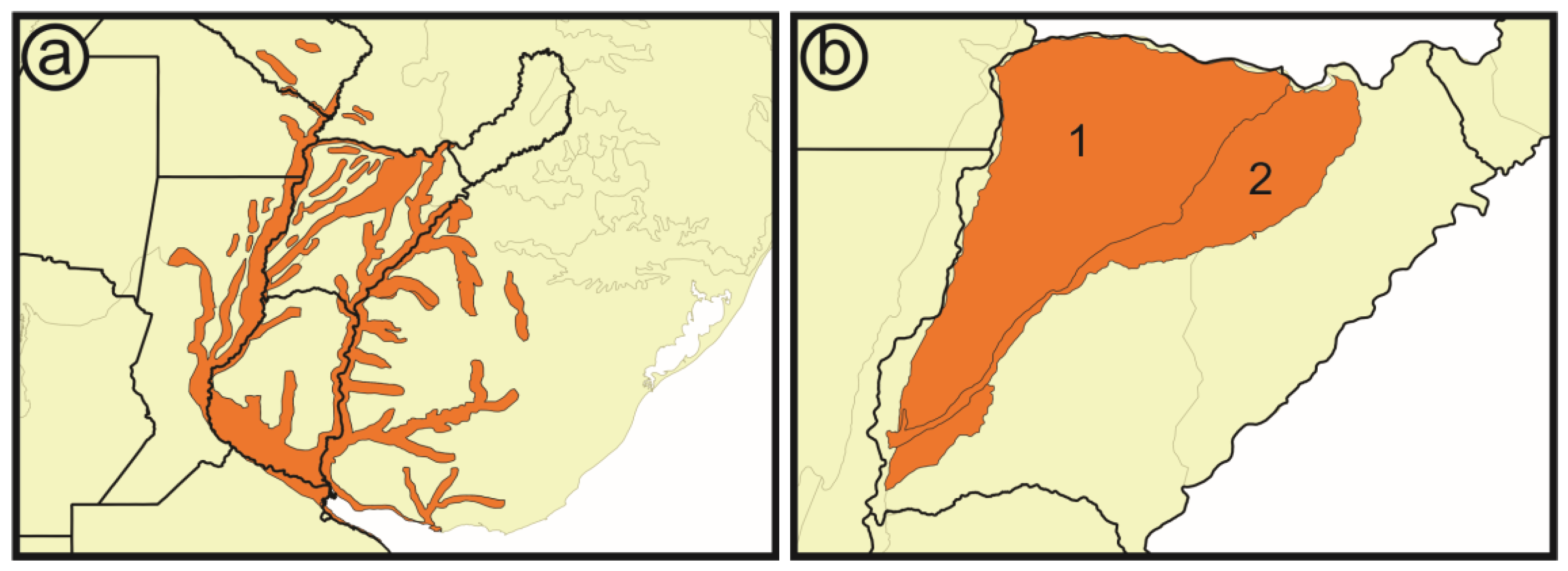
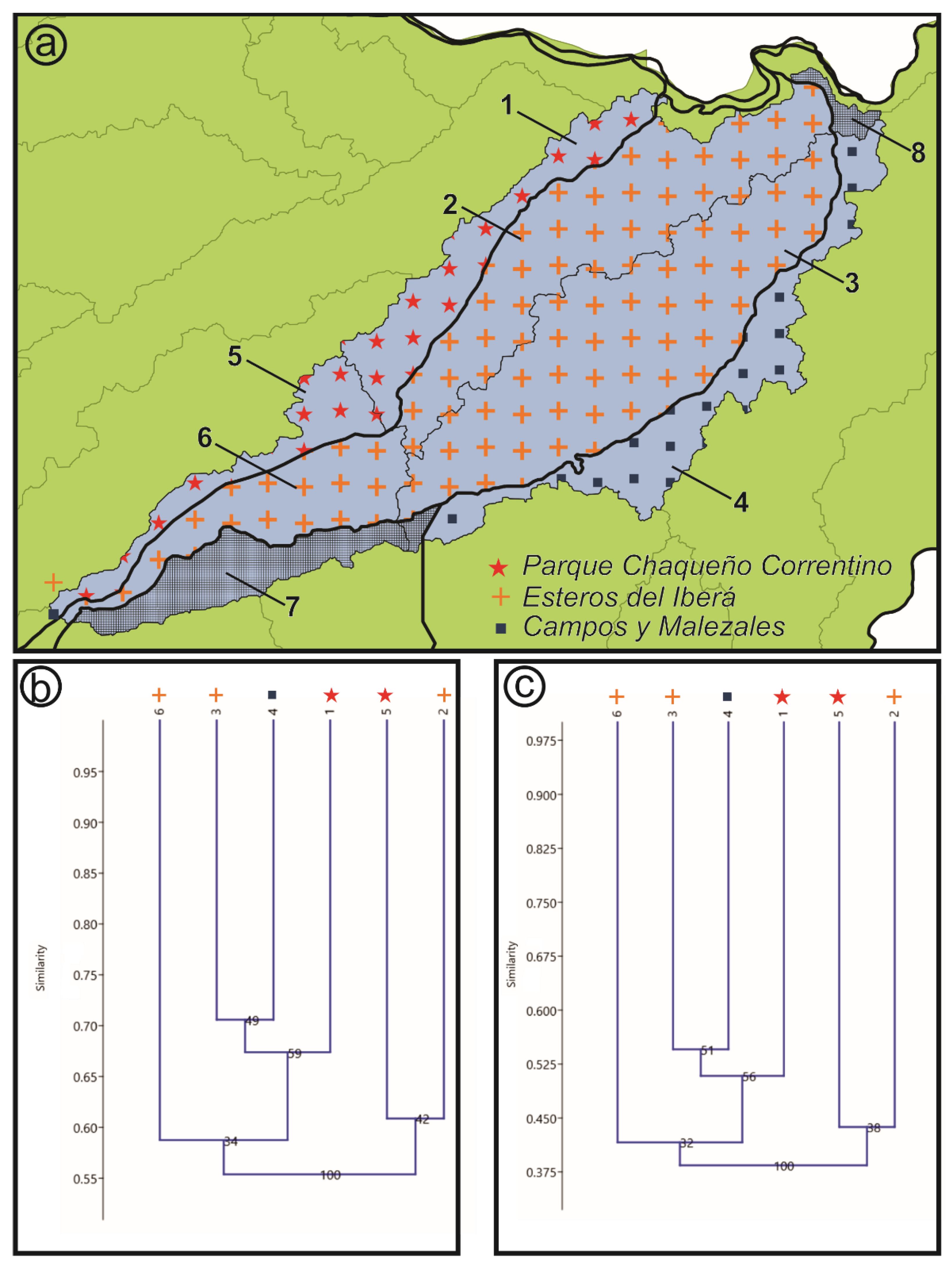
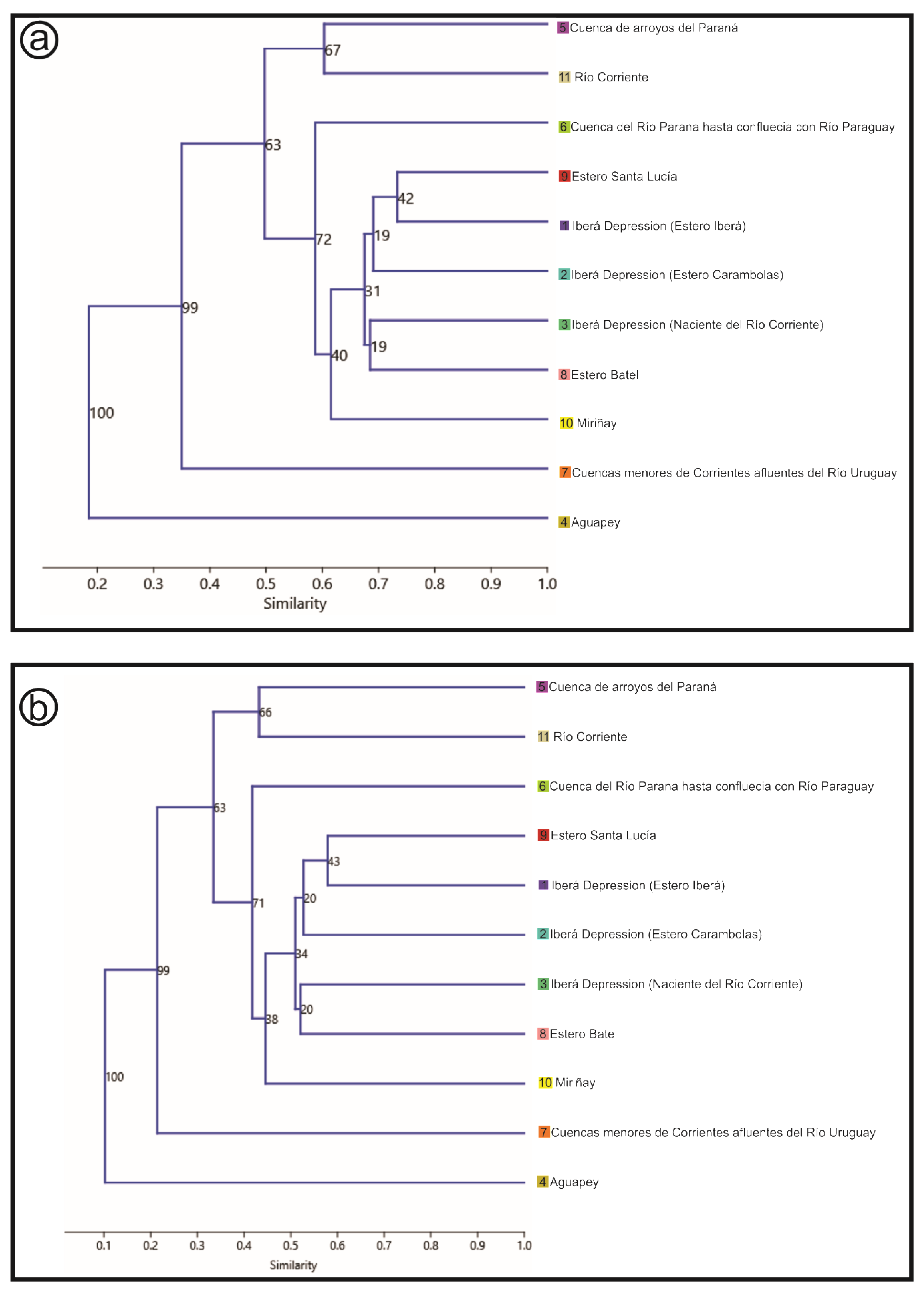
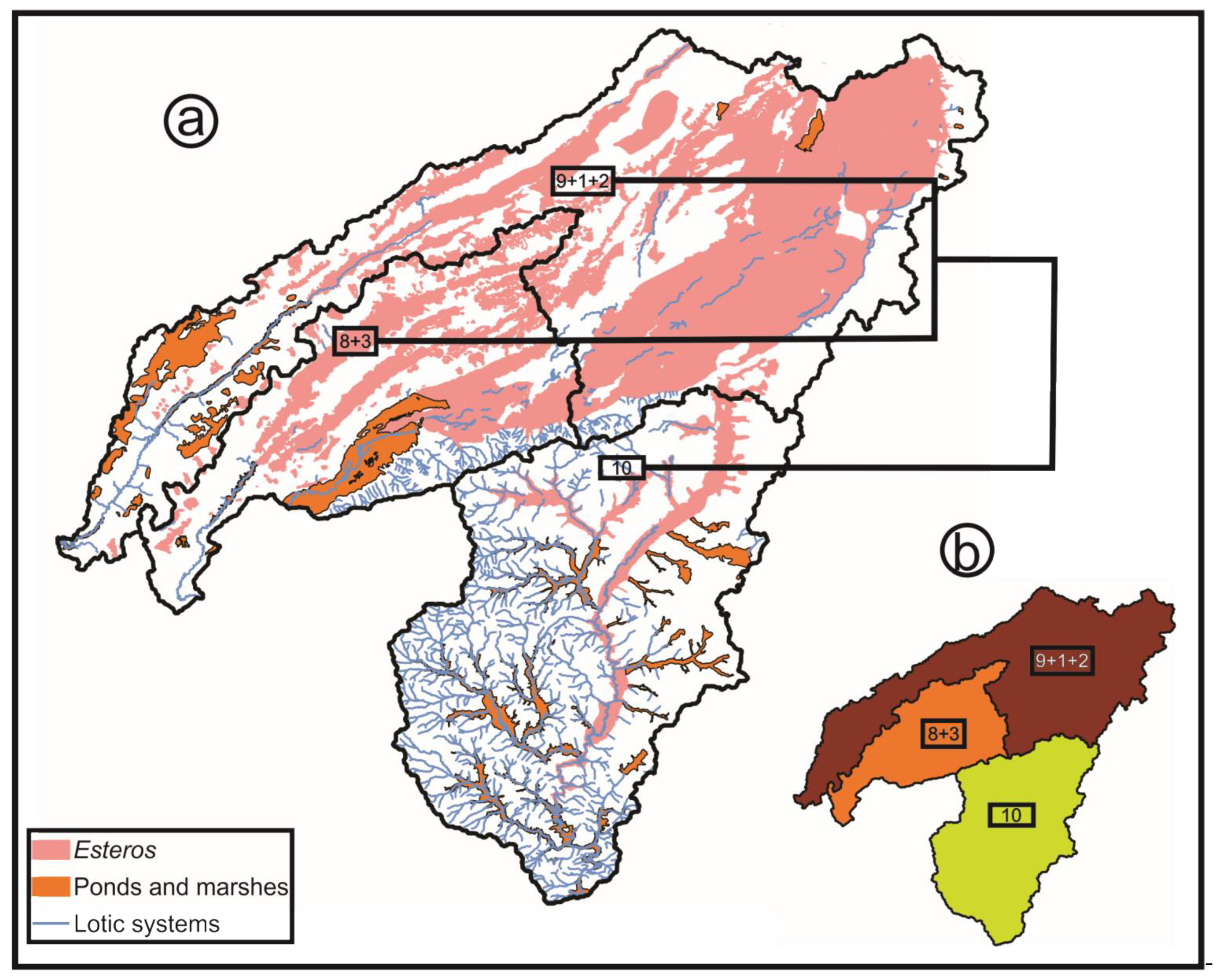
- Hydrological basins from Corrientes gathered from the Subsecretaría de Recursos Hídricos of Corrientes [21]: According to this scheme, the Iberá Depression is divided into three different sub-basins (Iberá, Carambolas and Naciente del Río Corriente). Surrounding these, there are eight basins that were used to test if the Iberá Depression functions as a unit (Aguapey, Cuenca de arroyos del Paraná, Cuenca del Río Paraná hasta confluencia con Río Paraguay, Cuencas menores de Corrientes afluentes del Río Uruguay, Estero Batel, Estero Santa Lucia, Miriñay, and Río Corriente) (Figure 4b).
2.4. Data Analysis
3. Results
3.1. Checklist
3.2. Biogeographical Provinces
3.3. Ecoregions
3.4. Basins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Noss, R.F. Indicators for Monitoring Biodiversity: A Hierarchical Approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Lehner, B.; Grill, G. Global River Hydrography and Network Routing: Baseline Data and New Approaches to Study the World’s Large River Systems. Hydrol. Process. 2013, 27, 2171–2186. [Google Scholar] [CrossRef]
- He, F.; Bremerich, V.; Zarfl, C.; Geldmann, J.; Langhans, S.D.; David, J.N.W.; Darwall, W.; Tockner, K.; Jähnig, S.C. Freshwater Megafauna Diversity: Patterns, Status and Threats. Divers. Distrib. 2018, 24, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Bailey, R.G. Identifying Ecoregion Boundaries. Environ. Manage. 2004, 34, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Lozano, F.; del Palacio, A.; Ramos, L.; Muzón, J. The Odonata of Argentina: State of knowledge and updated checklist. Int. J. Odonatol. 2020, 23, 113–153. [Google Scholar] [CrossRef]
- Muzón, J.; Pessacq, P.; Lozano, F. The Odonata (Insecta) of Patagonia: A Synopsis of their current status with illustrated keys for their identification. Zootaxa 2014, 3784, 346–388. [Google Scholar] [CrossRef] [Green Version]
- Muzón, J.; Von Ellenrieder, N.; Pessacq, P.; Lozano, F.; Garré, A.; Lambruschini, J.; Ramos, L.; Weigel Muñoz, S. Odonata from Iberá Wetlands (Corrientes, Argentina): Preliminary Inventory and Biodiversity. Rev. Soc. Entomol. Argent. 2008, 67, 59–67. [Google Scholar]
- Rubio, G.D.; Moreno, C.E. Orb-Weaving Spider Diversity in the Iberá Marshlands, Argentina. Neotrop. Entomol. 2010, 39, 496–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casciotta, J.R.; Almirón, A.E.; Bechara, J. Peces Del Iberá; Grafikar: La Plata, Argentina, 2005. [Google Scholar]
- Calcaterra, L.A.; Cuezzo, F.; Cabrera, S.M.; Briano, J.A. Ground Ant Diversity (Hymenoptera: Formicidae) in the Ibera Nature Reserve, the Largest Wetland of Argentina. Ann. Entomol. Soc. Am. 2010, 103, 71–83. [Google Scholar] [CrossRef]
- Coscarón, M.C.; Melo, M.C.; Coddington, J.; Corronca, J. Estimating Biodiversity: A Case Study on True Bugs in Argentinian Wetlands. Biodivers. Conserv. 2009, 18, 1491–1507. [Google Scholar] [CrossRef]
- Ingaramo, M.D.R.; Etchepare, E.G.; Álvarez, B.B.; Porcel, E. Riqueza y Composición de la Fauna de Anuros en la Región Oriental de la Reserva Natural Provincial Esteros Del Iberá, Corrientes, Argentina. Rev. Biol. Trop. 2012, 60, 759–769. [Google Scholar] [CrossRef]
- Dufek, M.I.; Oscherov, E.B.; Damborsky, M.P. Actualización de La Distribución de Tábanos (Diptera: Tabanidae) En La Reserva Natural Esteros Del Iberá, Argentina. Entomotropica 2016, 31, 36–39. [Google Scholar]
- Rubio, G.D.; Nadal, M.F.; Munévar, A.C.; Avalos, G.; Perger, R. Iberá Wetlands: Diversity Hotspot, Valid Ecoregion or Transitional Area? Perspective from a Faunistic Jumping Spiders Revision (Araneae: Salticidae). Species 2018, 19, 117–131. [Google Scholar]
- Lozano, F.; del Palacio, A.; Ramos, L.; Granato, L.; Drozd, A.; Muzón, J. Recovery of local dragonfly diversity following restoration of an artificial lake in an urban area near Buenos Aires. Basic Appl. Ecol. 2022, 58, 88–97. [Google Scholar] [CrossRef]
- Comunicación del CCT CONICET Incendios En Corrientes: Evaluarán Los Efectos a Mediano Plazo Sobre La Biodiversidad. Available online: https://nordeste.conicet.gov.ar/incendios-efectos-sobre-la-biodiversidad/ (accessed on 20 August 2022).
- Cabrera, A.L.; Willink, A. Biogeografía de América Latina; Programa Regional de Desarrollo Científico y Tecnológico, Organización de los Estados Americanos: Washington, DC, USA, 1980. [Google Scholar]
- Brown, A.; Pacheco, S. Propuesta de Actualización del Mapa Ecorregional de la Argentina; La situación: Ambient, Argentina, 2005; pp. 28–31. [Google Scholar]
- Herbst, R.; Jorge, N.; Cruz, S. Mapa Litoestratigráfico de la Provincia de Corrientes. Inst. Geol. Paleontol. 1985, 2, 1–50. [Google Scholar]
- Orfeo, O.; Neiff, J.J. Esteros del Iberá. Un Enorme Laboratorio a Cielo Abierto. Sitios Interés Geológico La República Argent: Argentina. 2008. Available online: http://repositorio.segemar.gov.ar/308849217/1343 (accessed on 1 October 2022).
- Secretaria de Recursos Hídricos. Atlas de Cuencas y Regiones Hídricas Superficiales de la República Argentina; 2010. Available online: https://www.argentina.gob.ar/obras-publicas/hidricas/cartografia-hidrica-provincial (accessed on 1 October 2022).
- Morrone, J.J.; Escalante, T.; Rodríguez-Tapia, G.; Carmona, A.; Arana, M.; Mercado-Gómez, J.D. Biogeographic regionalization of theNeotropical region: New map and shapefile. Ann. Acad. Bras. Cienc. 2022, 94, e20211167. [Google Scholar] [CrossRef]
- Apodaca, M.J.; Katinas, L.; Guerrero, E.L. Hidden areas of endemism: Small units in the South-eastern Neotropics. Syst. Biodivers. 2019, 17, 425–438. [Google Scholar] [CrossRef]
- Ribichich, A.M. El modelo clásico de la Fitogeografía de Argentina: UN Análisis Crítico. Interciencia 2002, 27, 669–675. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Cardoso, P.; Borges, P.A.; Veech, J.A. Testing the performance of beta diversity measures based on incidence data: The robustness to undersampling. Divers. Distrib. 2009, 15, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Singh, W. Robustness of three hierarchical agglomerative clustering techniques for Ecological Data. Master’s Thesis, University of Iceland, Reykjavík, Iceland, 2008; p. 100. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 40, ISBN 0444892508. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospat. Found. Proj. 2022. Available online: http://qgis.osgeo.org (accessed on 1 August 2022).
- López, H.L.; Menni, R.C.; Donato, M.; Miquelarena, A.M. Biogeographical Revision of Argentina (Andean and Neotropical Regions): An analysis using freshwater fishes. J. Biogeogr. 2008, 35, 1564–1579. [Google Scholar] [CrossRef]
- Ringuelet, R.A. Zoogeografía y Ecología de los Peces de Aguas Continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 1975, 2, 1–122. [Google Scholar]
- José de Paggi, S. Ecological and biogeographical remarks on the Rotifer Fauna of Argentina. Inst. Nac. Limnol. 1990, 23, 297–311. [Google Scholar]
- Martínez, S.J. Cuenca del Río Miriñay. Available online: https://www.argentina.gob.ar/sites/default/files/43.pdf (accessed on 15 August 2022).









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Palacio, A.; Lozano, F.; Ramos, L.S.; Navarro, M.d.l.M.; Muzón, J. Odonata from Iberá Wetland System (Corrientes, Argentina) Are Regional Biogeographic Schemes Useful to Assess Odonata Biodiversity and Its Conservation? Diversity 2022, 14, 842. https://doi.org/10.3390/d14100842
del Palacio A, Lozano F, Ramos LS, Navarro MdlM, Muzón J. Odonata from Iberá Wetland System (Corrientes, Argentina) Are Regional Biogeographic Schemes Useful to Assess Odonata Biodiversity and Its Conservation? Diversity. 2022; 14(10):842. https://doi.org/10.3390/d14100842
Chicago/Turabian Styledel Palacio, Alejandro, Federico Lozano, Lia S. Ramos, María de las Mercedes Navarro, and Javier Muzón. 2022. "Odonata from Iberá Wetland System (Corrientes, Argentina) Are Regional Biogeographic Schemes Useful to Assess Odonata Biodiversity and Its Conservation?" Diversity 14, no. 10: 842. https://doi.org/10.3390/d14100842