
Effect of Habitat Disturbance on the Composition of Soil Nematode Functional Groups Associated with a Tropical Herb: Heliconia collinsiana

 , and
, and 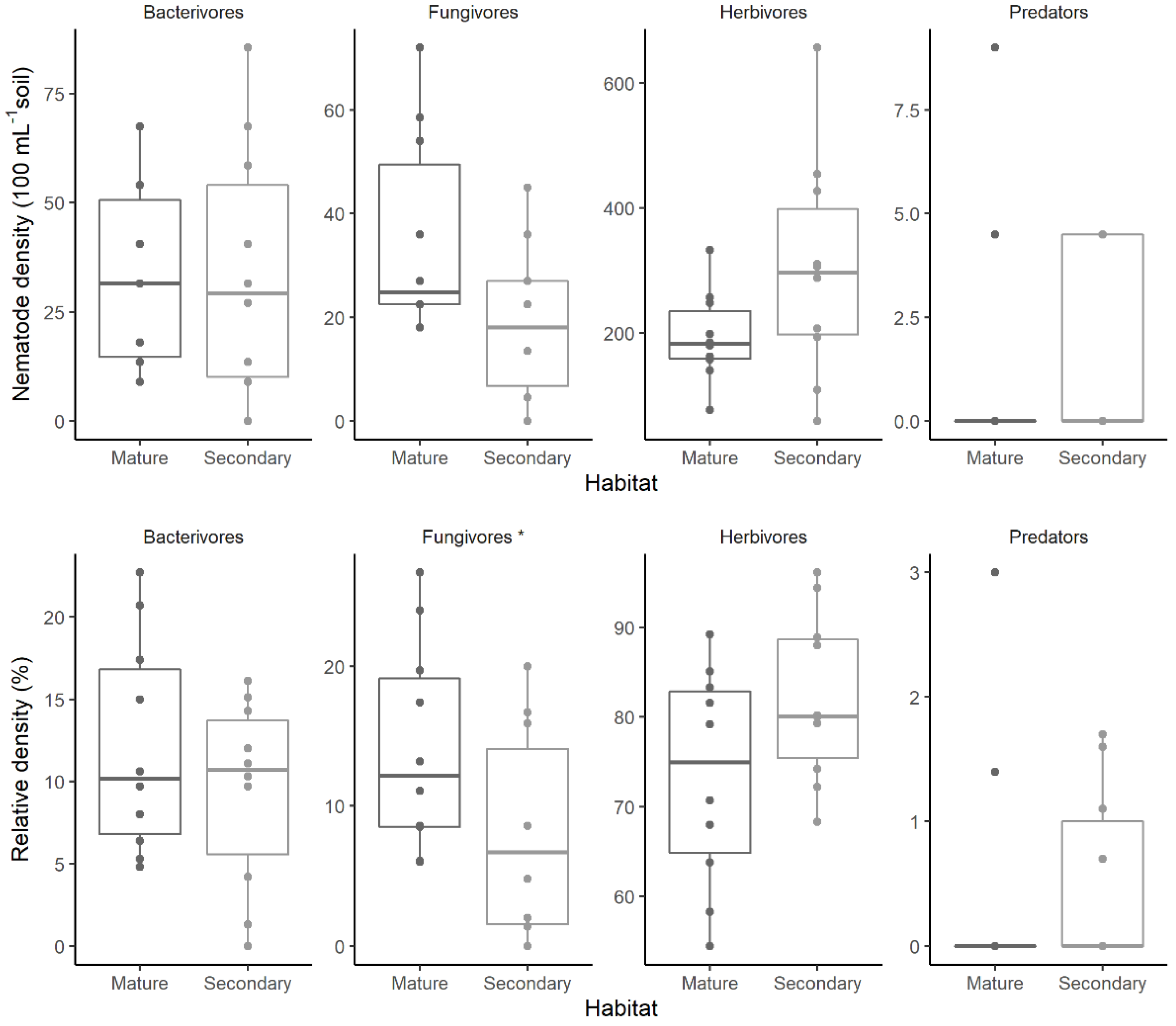{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Species
2.3. Nematodes Associated with the Genus Heliconia
2.4. Nematode Sampling and Processing
2.5. Nematode Extractions and Counts
2.6. Soil pH, Electrical Conductivity (EC), and Organic Matter Content
2.7. Statistical Analysis
3. Results
3.1. Nematode Density
3.2. Nematode Guild Composition
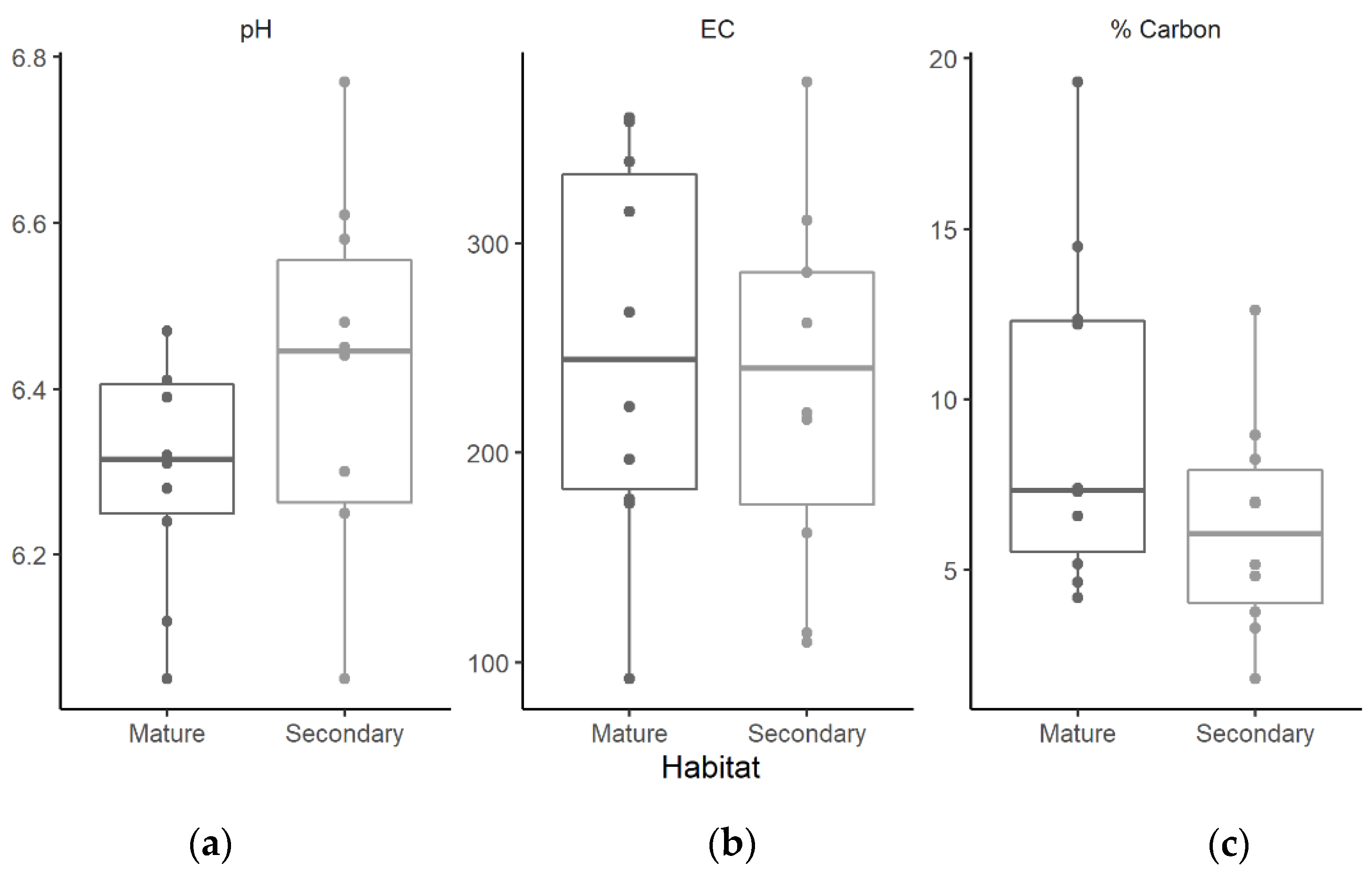
3.3. Soil pH, Electrical Conductivity, and Organic Matter Content
4. Discussion
4.1. Habitat Disturbance and Nematode Populations
4.2. Habitat Disturbance and Nematode Infection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayala-Orozco, B.; Gavito, M.E.; Mora, F.; Siddique, I.; Balvanera, P.; Jaramillo, V.J.; Cotler, H.; Romero-Duque, L.P.; Martínez-Meyer, E. Resilience of soil properties to land-use change in a tropical dry forest ecosystem. Land Degrad. Dev. 2018, 29, 315–325. [Google Scholar] [CrossRef]
- Carrillo-Saucedo, S.M.; Gavito, M.E. Resilience of soil aggregation and exocellular enzymatic functions associated with arbuscular mycorrhizal fungal communities along a successional gradient in a tropical dry forest. Mycorrhiza 2020, 30, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Allek, A.; Crouzeilles, R. Soil dynamics in forest restoration: A data set for temperate and tropical regions. Ecology 2021, 102, e03207. [Google Scholar] [CrossRef]
- Berkelmans, R.; Ferris, H.; Tenuta, M.; van Bruggen, A.H.C. Effects of long-term crop management on nematode trophic levels other than plant feeders disappear after 1 year of disruptive soil management. Appl. Soil Ecol. 2003, 23, 223–235. [Google Scholar] [CrossRef]
- Benítez-Malvido, J.; Giménez, A.; Graciá, E.; Rodríguez-Caro, R.C.; De Ybáñez, R.R.; Siliceo-Cantero, H.H.; Traveset, A. Impact of habitat loss on the diversity and structure of ecological networks between oxyurid nematodes and spur-thighed tortoises (Testudo graeca L.). PeerJ 2019, 7, e8076. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Malvido, J.; Martínez-Falcón, A.P.; Dáttilo, W.; del Val, E. Diversity and network structure of invertebrate communities associated to Heliconia species in natural and human disturbed tropical rain forests. Glob. Ecol. Conserv. 2014, 2, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Malvido, J.; Dáttilo, W.; Martínez-Falcón, A.P.; Durán-Barrón, C.; Valenzuela, J.; López, S.; Lombera, R. The multiple impacts of tropical forest fragmentation on arthropod biodiversity and on their patterns of interactions with host plants. PLoS ONE 2016, 11, e0146461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Malvido, J.; Dáttilo, W. Interaction intimacy of pathogens and herbivores with their host plants influences the topological structure of ecological networks in different ways. Am. J. Bot. 2015, 102, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Malvido, J. Fungal diseases in Neotropical forests disturbed by humans. In Conservation Medicine: Applied Cases of Ecological Health; Aguirre, A., Daszak, P., Ostfeld, R.S., Eds.; Oxford University Press: London, UK, 2012; pp. 302–311. [Google Scholar]
- Santos, B.A.; Benítez-Malvido, J. Insect herbivory and leaf disease in natural and human disturbed habitats: Lessons from early-successional Heliconia herbs. Biotropica 2012, 44, 53–62. [Google Scholar] [CrossRef]
- Benítez-Malvido, J.; Lázaro, A.; Ferraz, I.D. Effect of distance to edge and edge interaction on seedling regeneration and biotic damage in tropical rainforest fragments: A long-term experiment. J. Ecol. 2018, 106, 2204–2217. [Google Scholar] [CrossRef]
- Krishnadas, M.; Kumar, A.N.; Comita, L.S. Edge effects reduce a-diversity but not b-diversity during community assembly in a human-modified tropical forest. Ecol. Appl. 2019, 29, e01996. [Google Scholar] [CrossRef]
- Boag, B.; Yeates, G.W. Soil nematode biodiversity in terrestrial ecosystems. Biodivers. Conserv. 1998, 7, 617–630. [Google Scholar] [CrossRef]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161. [Google Scholar] [PubMed]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Hoogen, J.; Geisen, S.; Wall, D.H.; Wardle, D.A.; Traunspurger, W.; de Goede, R.G.; Crowther, T.W.; Adams, B.J.; Ahmad, W.; Ferris, H.; et al. A global database of soil nematode abundance and functional group composition. Sci. Data 2020, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ingham, R.E.; Trofymow, J.A.; Ingham, E.R.; Coleman, D.C. Interactions of bacteria, fungi, and their nematode grazers: Effects on nutrient cycling and plant growth. Ecol. Monogr. 1985, 55, 119–140. [Google Scholar] [CrossRef]
- Procter, D.L. Global overview of the functional roles of soil-living nematodes in terrestrial communities and ecosystems. J. Nematol. 1990, 22, 1–7. [Google Scholar]
- Crowther, T.W.; Boddy, L.; Jones, T.H. Species-specific effects of soil fauna on fungal foraging and decomposition. Oecologia 2011, 167, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreno, S.; Talavera, M. Nematodes as environmental indicators in agroecosystems. Ecosistemas 2013, 22, 50–55. [Google Scholar]
- Sewake, K.T.; Uchida, J.Y. Diseases of Heliconia in Hawaii; Research Extension 159; Agriculture and Human Resources: Honolulu, HI, USA, 1995. [Google Scholar]
- López-Cardona, N.; Castaño-Zapata, J. Characterization of Phytopathogenic Fungi, Bacteria, Nematodes and Viruses in Four Commercial Varieties of Heliconia (Heliconia sp.). Rev. Fac. Nac. Agron. Medellín 2012, 65, 6697–6710. [Google Scholar]
- Mattos-Sobrinho, C.C.; Silveira, A.J.; César, F.B.C.; Oliveira, C.M.G.; Bittencourt, M.A.L. Phytonematodes associated with Heliconia spp. in commercial crops in the South coast of Bahia, Brazil. Nematropica 2012, 42, 351–355. [Google Scholar]
- Aristizábal, L.F.; Ospina, K.A.; Vallejo, U.A.; Henao, E.R.; Salgado, M.; Arthurs, S.P. Entomofauna Associated with Heliconia spp. (Zingiberales: Heliconiaceae) Grown in the Central Area of Colombia. Fla. Entomol. 2013, 96, 112–119. [Google Scholar] [CrossRef]
- Hadley, A.S.; Frey, S.J.K.; Robinson, W.D.; Betts, M.G. Forest fragmentation and loss reduce richness, availability, and specialization in tropical hummingbird communities. Biotropica 2018, 50, 74–83. [Google Scholar] [CrossRef]
- Berry, F.; Kress, W.J. Heliconia: An Identification Guide (No. 635.93421); Smithsonian Institution Press: Washington, DC, USA, 1991. [Google Scholar]
- Assis, S.M.P.; Mariano, R.L.R.; Gondim, M.G.C., Jr.; Menezes, M.; Rosa, R.C.T. Disease and Pests of Heliconias/Doenças e Pragas das Helicônias; Editora da UFRPE: Recife, Brazil, 2002. [Google Scholar]
- Lugo-Cruz, E.; del Rivero-Bautista, N.; Sánchez-Soto, S.; Osorio-Osorio, R.; Romero-Nápoles, J. Insectos fitófagos asociados a cultivos de heliconias (Heliconia spp.) en Tabasco, México. Agroproductividad 2020, 13, 31–36. [Google Scholar]
- Alarcón-Restrepo, J.J. Enfermedades en la producción de heliconias en los departamentos de Caldas, Risaralda y Quindío. Agronomia 2007, 15, 45–61. [Google Scholar]
- Assis, T.C.; de Andrade, D.E.G.T. Fitonematoses em Zingiberales ornamentais no Estado de Pernambuco. An. Acad. Pernambucana Ciênc. Agron. 2007, 4, 185–198. [Google Scholar]
- Benítez-Malvido, J.; Lemus-Albor, A. The Seedling Community of Tropical Rain Forest Edges and Its Interaction with Herbivores and Pathogens. Biotropica 2005, 37, 301–313. [Google Scholar] [CrossRef]
- Medellín, R.A. Mammal diversity and conservation in the Selva Lacandona, Chiapas, Mexico. Conserv. Biol. 1994, 8, 780–799. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- López-Pérez, S.; Benítez-Malvido, J.; Lobato-García, J.M.; Siliceo-Cantero, H.H.; Santillán-Mendoza, R. A New State Record for Chelobasis bicolor Gray (Coleoptera: Chrysomelidae: Cassidinae: Arescini) and New Host Association with Heliconia bourgaeana Peterson (Heliconiaceae) in Mexico. Coleopt. Bull. 2020, 74, 572–575. [Google Scholar] [CrossRef]
- Lins, S.R.O.; Coelho, R.S.B. Ocorrência de Doenças em Plantas Ornamentais Tropicais no Estado de Pernambuco. Fitopatol. Bras. 2004, 29, 332–335. [Google Scholar] [CrossRef] [Green Version]
- Sardinha, D.H.S.; Rodrigues, A.A.C.; Diniz, N.B.; De Lemos, R.N.S.; Da Silva, G.S. Fungos e nematóides fitopatogênicos associados ao cultivo de flores tropicais em São Luís-MA. Summa Phytopathol. 2012, 38, 159–162. [Google Scholar] [CrossRef] [Green Version]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Walkley, A. A critical examination of a rapid method for determining organic carbon in soils-effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 1947, 63, 251–264. [Google Scholar] [CrossRef]
- Harrison, X. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2014, 2, e616. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.V. emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version, 1.8.1-1. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 26 September 2022).
- Villegas-Urbano, N.P.; Restrepo-Alarcón, J.J.; Galindo, J.R. Enfermedades Limitantes de la Producción de Heliconias en los Departamentos de Caldas, Risaralda y Quindío; Instituto Colombiano Agropecuario: Bogotá, Colombia, 2006.
- Burdon, J.J. Fungal pathogens as selective forces in plant populations and communities. Austral. J. Ecol. 1991, 16, 423–432. [Google Scholar] [CrossRef]
- Yeates, G.W.; Coleman, D.C. Nematodes in decomposition. In Nematodes in Soil Ecosystems; Freckman, D.W., Ed.; University of Texas: Austin, TX, USA, 1982. [Google Scholar]
- Brussaard, L.; Behan-Pelletier, V.M.; Bignell, D.E.; Brown, V.K.; Didden, W.; Folgarait, P.; Fragoso, C.; Freckman, D.W.; Gupta, V.V.S.R.; Hattori, T.; et al. Biodiversity and ecosystem functioning in soil. Ambio 1997, 26, 563–570. [Google Scholar]
- Gruden, K.; Lidoy, J.; Petek, M.; Podpečan, V.; Flors, V.; Papadopoulou, K.K.; Pappas, M.L.; Martinez-Medina, A.; Bejarano, E.; Biere, A.; et al. Ménage à trois: Unraveling the mechanisms regulating plant–microbe–arthropod interactions. Trends Plant Sci. 2020, 25, 1215–1226. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Escuer, M.; Lara, M.; Bello, A. Nematodos de la subfamilia Criconematinae (Nematoda: Criconematidae) en la España peninsular. Orsis 1997, 12, 39–63. Available online: www.raco.cat/index.php/Orsis/article/view/24394 (accessed on 10 September 2022).
- Jarosz, A.M.; Davelos, A.L. Effects of disease in wild plant populations and the evolution of pathogen aggressiveness. New Phytol. 1995, 129, 371–387. [Google Scholar] [CrossRef]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Frecklenton, R.P.; Lewis, O.T. Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. Dyn. Popul. 1971, 298, 312. [Google Scholar]
- van Bruggen, A.H.C.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benítez-Malvido, J.; Jaramillo-López, P.F.; Lobato-García, J.M.; Siliceo-Cantero, H.H.; Mora-Ardila, F.; Lombera, R. Effect of Habitat Disturbance on the Composition of Soil Nematode Functional Groups Associated with a Tropical Herb: Heliconia collinsiana. Diversity 2022, 14, 836. https://doi.org/10.3390/d14100836
Benítez-Malvido J, Jaramillo-López PF, Lobato-García JM, Siliceo-Cantero HH, Mora-Ardila F, Lombera R. Effect of Habitat Disturbance on the Composition of Soil Nematode Functional Groups Associated with a Tropical Herb: Heliconia collinsiana. Diversity. 2022; 14(10):836. https://doi.org/10.3390/d14100836
Chicago/Turabian StyleBenítez-Malvido, Julieta, Pablo Fabián Jaramillo-López, Juan Manuel Lobato-García, Héctor Hugo Siliceo-Cantero, Francisco Mora-Ardila, and Rafael Lombera. 2022. "Effect of Habitat Disturbance on the Composition of Soil Nematode Functional Groups Associated with a Tropical Herb: Heliconia collinsiana" Diversity 14, no. 10: 836. https://doi.org/10.3390/d14100836