
Aquatic Macrophyte Vegetation Promotes Taxonomic and Functional Diversity of Odonata Assemblages in Intermittent Karst Rivers in the Mediterranean

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
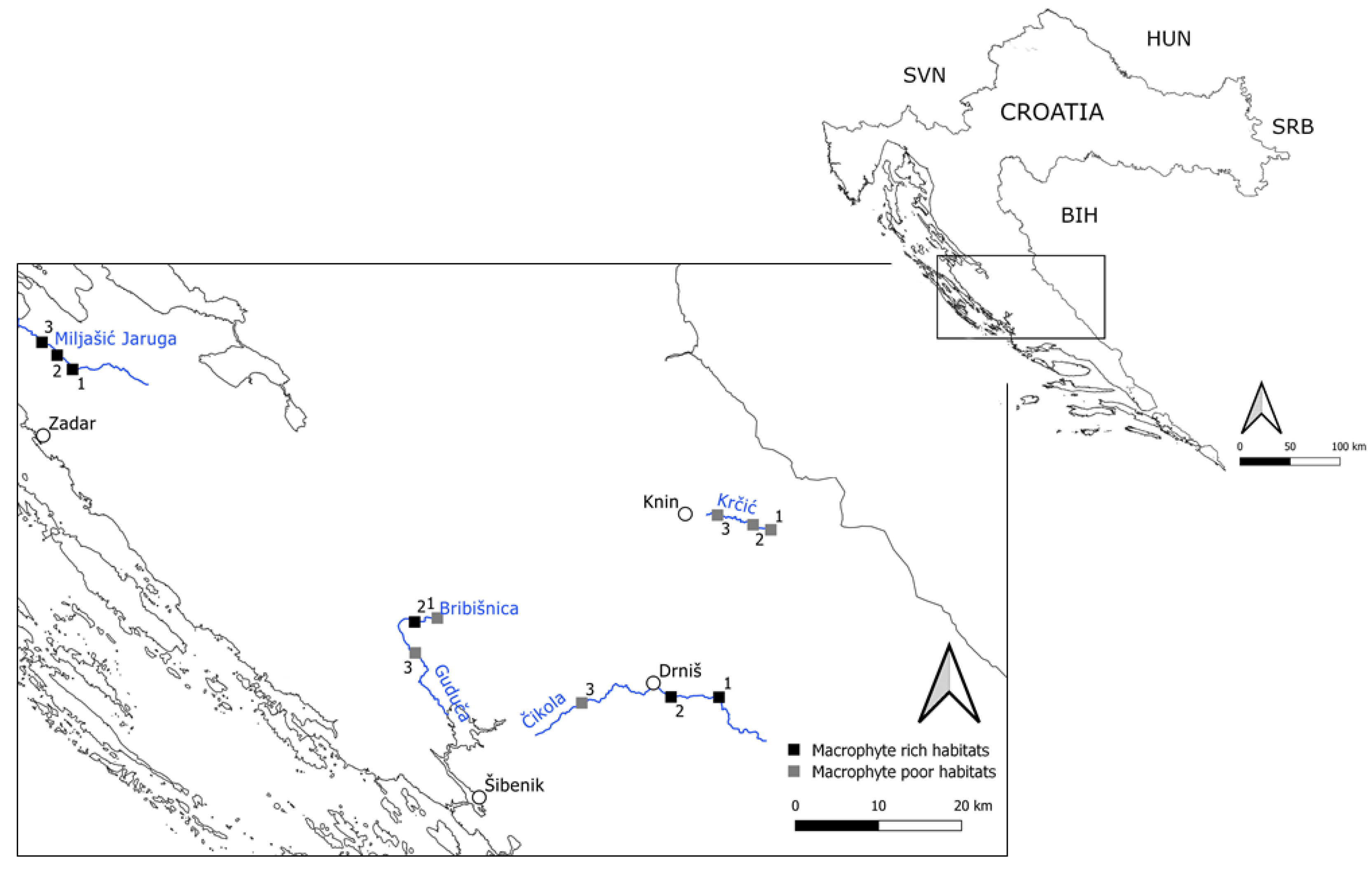
2.1. Study Area
2.2. Vegetation Analysis
2.3. Environmental Variables
2.4. Odonata Sampling
2.5. Data Analyses
3. Results
3.1. Vegetation Analysis
3.2. Environmental Variables
3.3. Odonata Species Occurrence
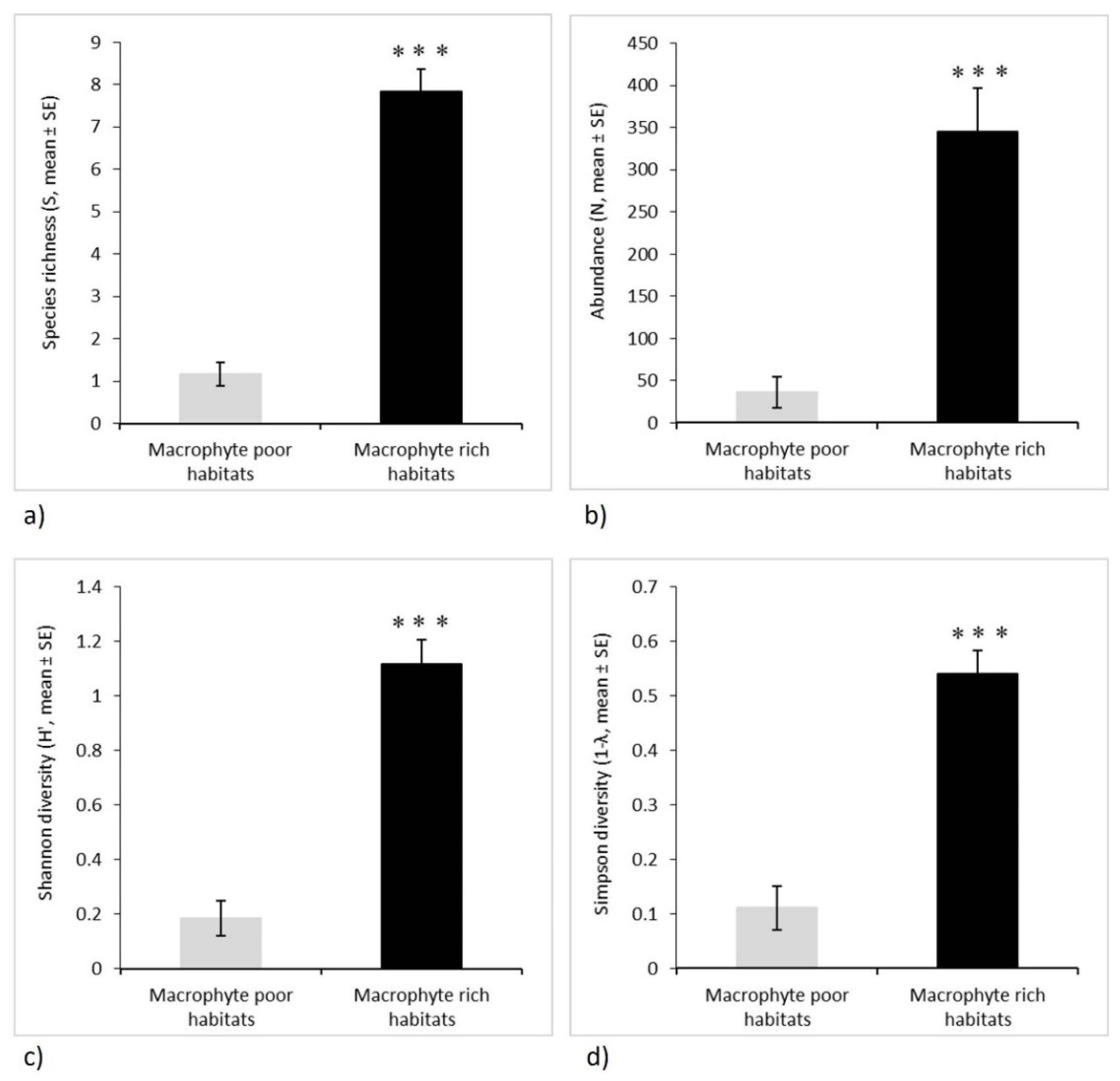
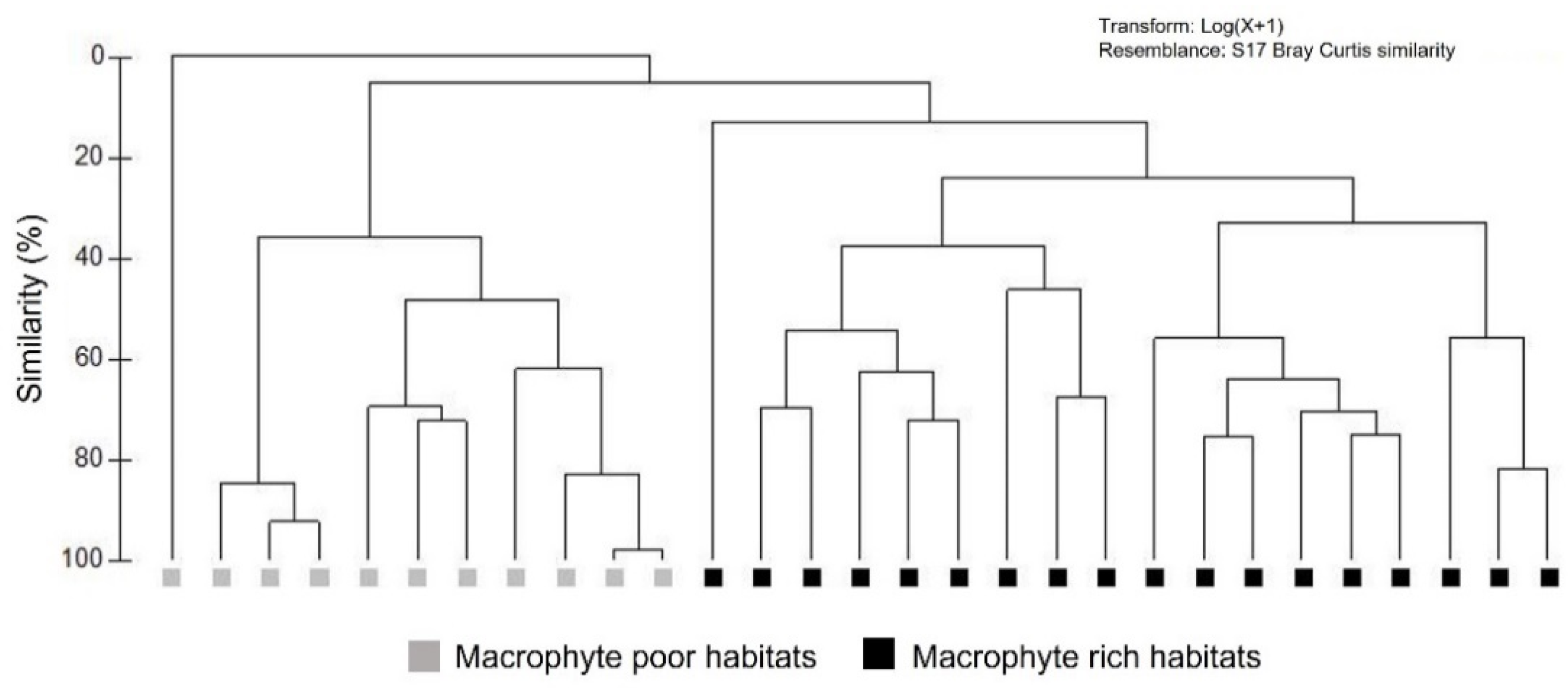
3.4. Odonata Assemblages and Their Functional Diversity
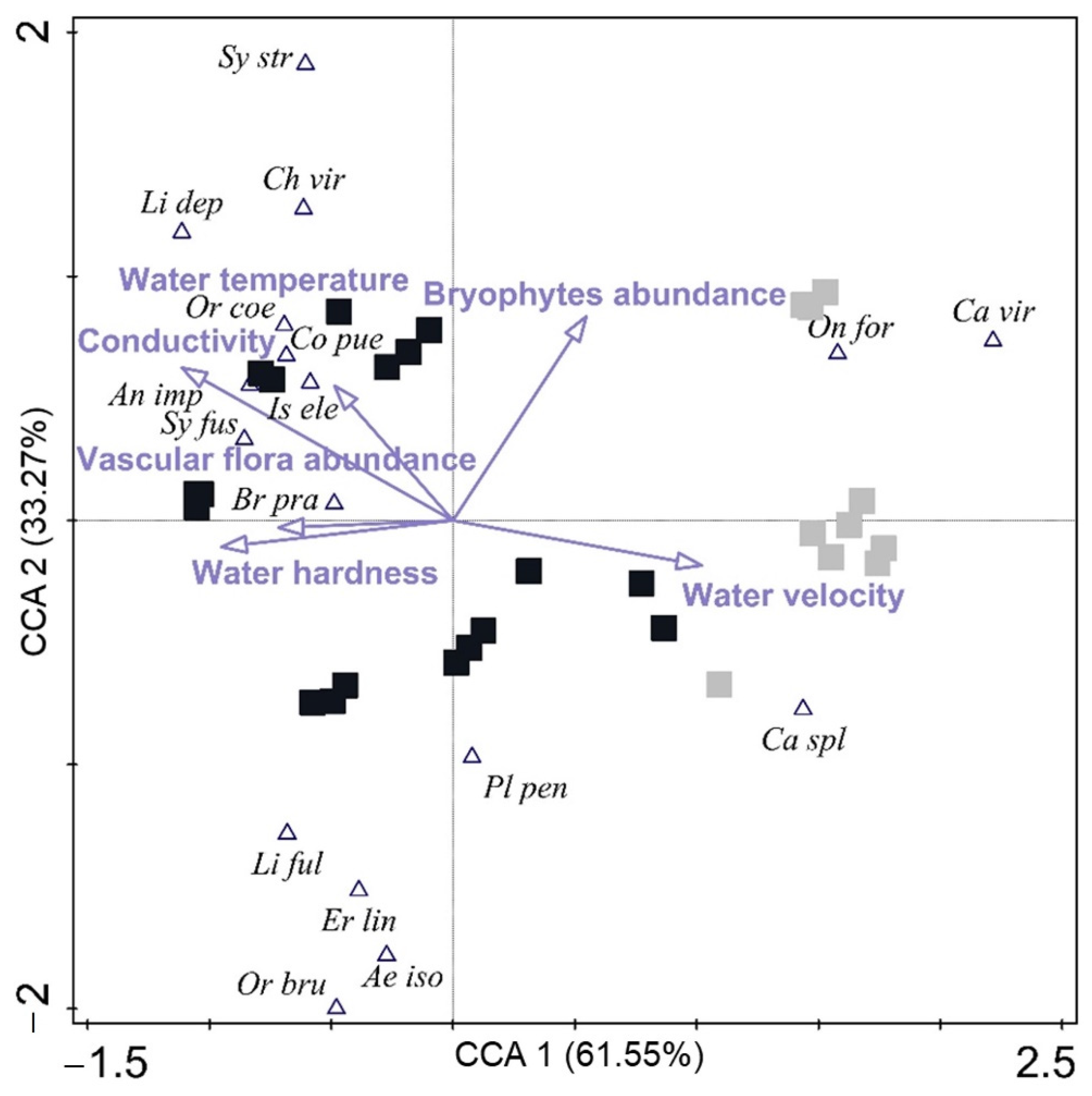
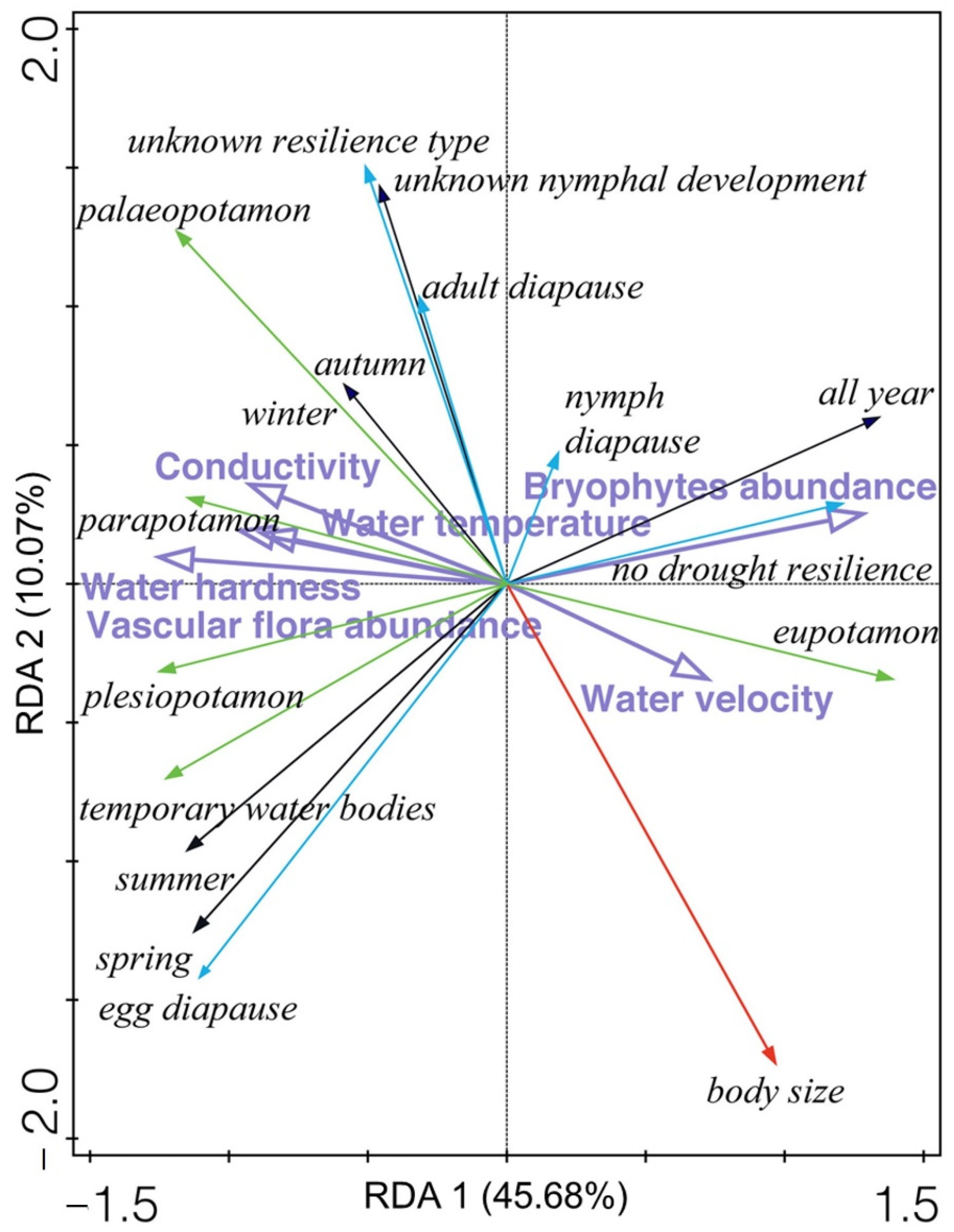
3.5. Odonata Species and Functional Traits Related to Environmental Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Functional Traits | |||
|---|---|---|---|---|
| Average Body Size (mm) | Hydrological Preference | Nymphal Development | Drought Resilience Form | |
| Calopteryx splendens (Harris, 1782) | 46.5 | Lotic (eupotamon, parapotamon) | All year | No drought resilience |
| Calopteryx virgo (Linnaeus, 1758) | 47.0 | Lotic (eupotamon) | All year | No drought resilience |
| Chalcolestes viridis (Vander Linden, 1825) | 43.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats. Recorded from temporary waterbodies. | Spring, summer | Egg diapause |
| Sympecma fusca (Vander Linden, 1820) | 36.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats. Recorded from temporary waterbodies. | Mainly in summer | Adult diapause |
| Ischnura elegans (Van der Linden, 1820) | 32.0 | Eurytopic. Recorded from temporary waterbodies | All year | No drought resilience |
| Coenagrion puella (Linnaeus, 1758) | 34.0 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats. Recorded from temporary waterbodies. | All year | No drought resilience |
| Erythromma lindenii (Selys, 1840) | 33.0 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats | Unknown | Unknown |
| Platycnemis pennipes (Pallas, 1771) | 36.0 | Predominantly lotic (eupotamon, parapotamon) but can occur in lentic habitats | All year | No drought resilience |
| Aeshna affinis Vander Linden, 1820 | 61.5 | Lentic (palaeopotamon). Recorded from temporary waterbodies | Spring, summer | Egg diapause |
| Aeshna isoceles (Müller, 1767) | 64.0 | Lentic (plesiopotamon, palaeopotamon) | All year | No drought resilience |
| Anax imperator (Selys, 1839) | 75.0 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats | All year | No drought resilience |
| Brachytron pratense (Müller, 1764) | 58.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats | All year | No drought resilience |
| Onychogomphus forcipatus (Linnaeus, 1758) | 48.0 | Lotic (eupotamon, parapotamon) | All year | No drought resilience |
| Cordulegaster heros Theischinger, 1979 | 80.5 | Lotic (eupotamon) | All year | No drought resilience |
| Somatochlora meridionalis Nielsen, 1935 | 52.5 | Lotic (eupotamon, parapotamon) | All year | No drought resilience |
| Libellula depressa Linnaeus, 1758 | 43.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats. Recorded from temporary waterbodies. | All year | Nymph diapause |
| Libellula fulva Müller, 1764 | 43.5 | Predominantly lotic (eupotamon, parapotamon) but can occur in lentic habitats | All year | Nymph diapause |
| Orthetrum cancellatum (Linnaeus, 1758) | 47.0 | Predominantly lotic (eupotamon, parapotamon) but can occur in lentic habitats | All year | No drought resilience |
| Orthetrum coerulescens (Fabricius, 1798) | 40.5 | Predominantly lotic (eupotamon, parapotamon) but can occur in lentic habitats. Recorded from temporary waterbodies | All year | Nymph diapause |
| Orthetrum brunneum (Fonscolombe, 1837) | 45.0 | Lotic (eupotamon, parapotamon) | All year | Unknown |
| Sympetrum sanguineum (Müller, 1764) | 36.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats. Recorded from temporary waterbodies. | Spring, summer | Egg diapause |
| Sympetrum fonscolombii (Selys, 1840) | 36.5 | Eurytopic. Recorded from temporary waterbodies | All year | Unknown |
| Sympetrum striolatum (Charpentier, 1840) | 39.5 | Eurytopic. Recorded from temporary waterbodies | All year | Egg diapause |
| Sympetrum meridionale (Selys, 1841) | 37.5 | Lentic (palaeopotamon). Recorded from temporary waterbodies | Spring, summer | Egg diapause |
| Crocothemis erythraea (Brullé, 1832) | 40.5 | Predominantly lentic (plesiopotamon, palaeopotamon), but can occur in lotic habitats | All year | No drought resilience |
References
- Lytle, D.A.; Poff, N.L. Adaptation to natural flow regimes. Trends Ecol. Evol. 2004, 19, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Datry, T.; Bonada, N.; Boulton, A. Intermittent Rivers and Ephemeral Streams: Ecology and Management, 1st ed.; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- McDonοugh, O.T.; Hosen, J.D.; Palmer, M.A. Temporary streams: The hydrology, geography and ecology of non-perennially flowing waters. In River Ecosystems: Dynamics, Management and Conservation, 1st ed.; Elliot, H.S., Martin, L.E., Eds.; Nova Science Publ. Inc.: New York, NY, USA, 2011; pp. 259–289. [Google Scholar]
- Arthington, A.H.; Bernardo, J.M.; Ilhèu, M. Temporary rivers: Linking ecohydrology, ecological quality and reconciliation ecology. River Res. Appl. 2014, 30, 1209–1215. [Google Scholar] [CrossRef]
- Tockner, K.; Robinson, C.T.; Uehlinger, U. Rivers of Europe, 1st ed.; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Larned, S.T.; Datry, T.; Arscott, D.B.; Tockner, K. Emerging concepts in temporary-river ecology. Freshw. Biol. 2010, 55, 717–738. [Google Scholar] [CrossRef]
- Vörösmarty, C.; McIntyre, P.; Gessner, M.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Reidy Liermann, A.; et al. Global threats to human water security and river biodiversity. Nature 2010, 468, 334. [Google Scholar] [CrossRef]
- Stubbington, R.; Chadd, R.; Cid, N.; Csabai, Z.; Miliša, M.; Morais, M.; Munné, A.; Pařil, P.; Pešić, V.; Tziortzis, I.; et al. Biomonitoring of intermittent rivers and ephemeral streams in Europe: Current practice and priorities to enhance ecological status assessments. Sci. Total Environ. 2018, 618, 1096–1113. [Google Scholar] [CrossRef] [PubMed]
- Boix, D.; García-Berthou, E.; Gascón, S.; Benejam, L.L.; Tornés, E.; Sala, J.; Benito, J.; Munné, A.; Solà, C.; Sabater, S. Response of community structure to sustained drought in Mediterranean rivers. J. Hydrol. 2010, 383, 135–146. [Google Scholar] [CrossRef]
- Gleick, P.H. Global freshwater resources: Soft-path solutions for the 21st century. Science 2003, 302, 1524–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanafield, M.; Bourke, S.A.; Zimmer, M.A.; Costigan, K.H. An overview of the hydrology of non-perennial rivers and streams. WIREs Water 2021, 8, 1–25. [Google Scholar] [CrossRef]
- Döll, P.; Schmied, H.M. How is the impact of climate change on river flow regimes related to the impact on mean annual runoff? A global-scale analysis. Environ. Res. Lett. 2012, 7, 14–37. [Google Scholar] [CrossRef] [Green Version]
- Datry, T.; Larned, S.T.; Tockner, T. Intermittent rivers: A challenge for freshwater ecology. BioScience 2014, 64, 229–235. [Google Scholar] [CrossRef] [Green Version]
- del Rosario, R.B.; Resh, V.H. Invertebrates in intermittent and perennial streams: Is the hyporheic zone a refuge from drying? J. N. Am. Benthol. Soc. 2000, 19, 680–696. [Google Scholar] [CrossRef]
- Bonada, N.; Zamora-Munoz, C.; Rieradevall, M.; Prat, N. Ecological profiles of caddisfly larvae in Mediterranean streams: Implications for bioassessment methods. Environ. Pollut. 2004, 132, 509–521. [Google Scholar] [CrossRef]
- Argyroudi, A.; Chatzinikolaou, Y.; Poirazidis, K.; Lazaridou, M. Do intermittent and ephemeral Mediterranean rivers belong to the same river type? Aquat. Ecol. 2009, 43, 465–476. [Google Scholar]
- Government of the Republic of Croatia (GRC). Directive on Water Classification. Available online: https://www.irb.hr/ (accessed on 28 November 2021).
- Golfieri, B.; Hardersen, S.; Maiolini, B.; Surian, N. Odonates as indicators of the ecological integrity of the river corridor: Development and application of the Odonate River Indec (ORI) in northern Italy. Ecol. Indic. 2016, 61, 234–247. [Google Scholar] [CrossRef]
- Martín, R.; Maynou, X. Dragonflies (Insecta: Odonata) as indicators of habitat quality in Mediterranean streams and rivers in the province of Barcelona (Catalonia, Iberian Peninsula). Int. J. Odonatol. 2016, 19, 107–124. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Shimano, Y.; Gardner, T.A.; Hughes, R.M.; De Marco, P.; Juen, L. Neotropical dragonflies (Insecta: Odonata) as indicators of ecological condition of small streams in the eastern Amazon. Austral Ecol. 2015, 40, 733–744. [Google Scholar] [CrossRef]
- O’Malley, Z.G.; Compson, Z.G.; Orlofske, J.M.; Baird, D.J.; Curry, R.A.; Monk, W.A. Riparian and in-channel habitat properties linked to dragonfly emergence. Sci. Rep. 2020, 10, 17665. [Google Scholar] [CrossRef]
- Vilenica, M. Ecological traits of dragonfly (Odonata) assemblages along an oligotrophic Dinaric karst hydrosystem. Ann. Limnol. -Int. J. Lim. 2017, 53, 377–389. [Google Scholar] [CrossRef]
- Vilenica, M.; Kerovec, M.; Pozojević, I.; Mihaljević, Z. Odonata assemblages in anthropogenically impacted lotic habitats. J. Limnol. 2020, 80, 1–9. [Google Scholar] [CrossRef]
- Brito, J.; Calvão, L.; Cunha, E.; Maioli, L.; Barbirato, M.; Rolim, S.; Juen, L. Environmental variables affect the diversity of adult damselflies (Odonata: Zygoptera) in western Amazonia. Int. J. Odonatol. 2021, 24, 108–121. [Google Scholar] [CrossRef]
- Adu, B.W.; Amusan, B.O.; Oke, T.O. Assessment of the water quality and Odonata assemblages in three waterbodies in Ilara-Mokin, south-western Nigeria. Int. J. Odonatol. 2019, 22, 101–114. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Juen, L. Structuring of dragonfly communities (insecta: Odonata) in Eastern Amazon: Effects of environmental and spatial factors in preserved and altered streams. Insects 2019, 10, 322. [Google Scholar] [CrossRef] [Green Version]
- Brito, J.S.; Michelan, T.S.; Juen, L. Aquatic macrophytes are important substrates for Libellulidae (Odonata) larvae and adults. Limnology 2021, 22, 139–149. [Google Scholar] [CrossRef]
- McAbendroth, L.; Ramsay, P.M.; Foggo, A.; Rundle, S.D.; Bilton, D.T. Does macrophyte fractal complexity drive invertebrate diversity, biomass and body size distributions? Oikos 2005, 111, 279–290. [Google Scholar] [CrossRef]
- Silva, C.V.; Henry, R. Aquatic invertebrates assemblages associated with two floating macrophytes species of contrasting root systems in a tropical wetland. Limnology 2020, 21, 107–118. [Google Scholar] [CrossRef]
- Vilenica, M.; Alegro, A.; Koletić, N.; Mihaljević, Z. New evidence of Lindenia tetraphylla (Vander Linden, 1825) (Odonata, Gomphidae) reproduction at the north-western border of its distribution. Nat. Croat. 2016, 25, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Calvão, L.B.; Juen, L.; Oliveira-Junior, J.M.B.; Batista, J.D.; De, M.P.J. Land use modifies Odonata diversity in streams of the Brazilian Cerrado. J. Insect Conserv. 2018, 22, 675–685. [Google Scholar] [CrossRef]
- Huikkonen, I.; Helle, I.; Elo, M. Heterogenic aquatic vegetation promotes abundance and species richness of Odonata (Insecta) in constructed agricultural wetlands. Insect Conserv. Divers. 2019, 13, 374–383. [Google Scholar] [CrossRef]
- Vilenica, M.; Pozojević, I.; Vučković, N.; Mihaljević, Z. How suitable are man-made water bodies as habitats for Odonata? Knowl. Manag. Aquat. Ecosyst. 2020, 421, 1–10. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behavior and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- Stoks, R.; McPeek, M.A. Predators and life histories shape Lestes damselfly assemblages along a freshwater habitat gradient. Ecology 2003, 84, 1576–1587. [Google Scholar] [CrossRef]
- Magnusson, A.K.; Williams, D.D. The roles of natural temporal and spatial variation versus biotic influences in shaping the physicochemical environment of intermittent ponds: A case study. Arch. Hydrobiol. 2006, 165, 537–556. [Google Scholar] [CrossRef]
- Chester, E.T.; Robson, B.J. Drought refuges, spatial scale and recolonisation by invertebrates in non-perennial streams. Freshw. Biol. 2011, 56, 2094–2104. [Google Scholar] [CrossRef]
- Deacon, C.; Samways, M.J.; Pryke, J.S. Aquatic insects decline in abundance and occupy low-quality artificial habitats to survive hydrological droughts. Freshw. Biol. 2019, 64, 1643–1654. [Google Scholar] [CrossRef]
- Illies, J. Limnofauna Europaea, 2nd ed.; Gustav Fischer Verlag: Stuttgart, Germany, 1978. [Google Scholar]
- Mihevc, A.; Zupan-Hajna, N.; Prelovšek, M. Case study from the Dinaric karst of Slovenia. In Introduction to the Dinaric Karst, 1st ed.; Mihevc, A., Prelovšek, M., Zupan Hajna, N., Eds.; Karst Research Institute: Postojna, Slovenia, 2010; pp. 49–66. [Google Scholar]
- Bonacci, O.; Željković, I.; Galić, A. Karst rivers’ particularity: An example from Dinaric karst (Croatia/Bosnia and Herzegovina). Environ. Earth Sci. 2013, 70, 963–974. [Google Scholar] [CrossRef]
- Previšić, A.; Walton, C.; Kučinić, M.; Mitrikeski, P.T.; Kerovec, M. Pleistocene divergence of Dinaric Drusus endemics (Trichoptera, Limnephilidae) in multiple microrefugia within the Balkan Peninsula. Mol. Ecol. 2009, 18, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Suhling, F.; Sahlén, G.; Gorb, S.; Kalkman, V.J.; Dijkstra, K.D.B.; van Tol, J. Order odonata. In Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Thorp, J., Rogers, D.C., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 893–932. [Google Scholar] [CrossRef]
- Pešić, V.; Gligorović, B.; Savić, A.; Buczyński, P. Ecological patterns of Odonata assemblages in karst springs in central Montenegro. Knowl. Manag. Aquat. 2017, 418, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, O. Hydrological investigations of Dinaric Karst at the Krčić catchment and the River Krka springs (Yugoslavia). J. Hydrol. 1985, 82, 317–326. [Google Scholar] [CrossRef]
- Bonacci, O.; Jukić, D.; Ljubenkov, I. Definition of catchment area in karst: Case of the rivers Krčić and Krka, Croatia. Hydrol. Sci. J. 2006, 51, 682–699. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, O.; Terzić, J.; Roje-Bonacci, T. Hidrološka analiza krške rijeke Čikole. Hrvat. Vode 2018, 26, 281–292. [Google Scholar]
- Bonacci, O.; Terzić, J.; Roje-Bonacci, T.; Frangen, T. An intermittent karst river: The case of the Čikola River (Dinaric Karst, Croatia). Water 2019, 11, 2415. [Google Scholar] [CrossRef] [Green Version]
- Brkić, Ž.; Biondić, R.; Pavičić, A.; Slišković, I.; Marković, T.; Terzić, J.; Dukarić, F.; Dolić, M. Određivanje Cjelina Podzemnih Voda na Jadranskom Slivu prema Kriterijima Okvirne Direktive o Vodama EU; Hrvatski Geološki Institute: Zagreb, Croatia, 2006. [Google Scholar]
- Magaš, D. Geographical factors of formation and development of the neolithic settlement Crno Vrilo. In Crno Vrilo, 1st ed.; Marijanović, B., Ed.; University of Zadar: Zadar, Croatia, 2009; pp. 7–23. [Google Scholar]
- Lukač Reberski, J.; Rubinić, J.; Terzić, J.; Radišić, M. Climate change impacts on groundwater resources in the coastal karstic Adriatic area: A case study from the Dinaric karst. Nat. Resour. Res. 2020, 29, 1975–1988. [Google Scholar] [CrossRef]
- Cukrov, N.; Frančišković-Bilinski, S.; Mikac, N.; Roje, V. Natural and anthropogenic influences recorded in sediments from the Krka river estuary (Eastern Adriatic coast), evaluated by statistical methods. Fresnius Environ. Bull. 2008, 17, 855–863. [Google Scholar]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische bemerkungen und vorschläge zur quantitativen vegetationsanalyse. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1964. [Google Scholar]
- Dierschke, H. Pflanzensoziologie: Grundlagen und Methoden, 1st ed.; UTB: Stuttgart, Germany, 1994. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Dijkstra, K.D.B.; Lewington, R. Field Guide to the Dragonflies of Britain and Europe, 1st ed.; British Wildlife Publishing: Gillingham, UK, 2006. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows, Version 27.0; IBM: Armonk, NY, USA, 2020. [Google Scholar]
- Field, A. Discovering Statistics Using SPSS, 3rd ed.; SAGE Publications: London, UK, 2009. [Google Scholar]
- Thiele, J.; Markussen, B. Potential of GLMM in modelling invasive spread. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2012, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual/Tutorial; Plymouth: Auckland, New Zealand, 2006. [Google Scholar]
- Freshwaterecology. Available online: https://www.freshwaterecology.info/fwe_search.php?og=mzb (accessed on 3 November 2021).
- Schmidt-Kloiber, A.; Hering, D. www.freshwaterecology.info—An online tool that unifies, standardises and codifies more than 20,000 European freshwater organisms and their ecological preferences. Ecol. Indic. 2015, 53, 271–282. [Google Scholar] [CrossRef]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Kotarac, M.; Šalamun, A.; Vilenica, M. EU Natura 2000 Integration Project: Field Research and Laboratory Processing for Collecting New Inventory Data for Taxonomic Groups: Actinopterygii and Cephalaspidomorphi, Amphibia and Reptilia, Aves, Chiroptera, Decapoda, Lepidoptera, Odonata, Plecoptera–Final Report for the Taxonomic Group Odonata; Ministry of Environmental and Nature Protection: Zagreb, Croatia, 2016.
- Boudot, J.P.; Kalkman, V.J. Atlas of the European Dragonflies and Damselflies; KNNV Uitgeverij: Zeist, The Netherlands, 2015. [Google Scholar]
- Hardersen, S. Dragonfly (Odonata) communities at three lotic sites with different hydrological characteristics. Ital. J. Zool. 2008, 75, 271–283. [Google Scholar] [CrossRef]
- Belmar, O.; Velasco, J.; Gutiérrez-Cánovas, C.; Mellado-Díaz, A.; Millán, A.; Wood, P.J. The influence of natural flow regimes on macroinvertebrate assemblages in a semiarid Mediterranean basin. Ecohydrology 2012, 6, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Yalles Satha, A.; Samraoui, B. Environmental factors influencing Odonata communities of three Mediterranean rivers: Kebir-East, Seybouse, and Rhummel wadis, northeastern Algeria. Rev. Ecol. 2017, 72, 314–329. [Google Scholar]
- Bonada, N.; Rieradevall, M.; Prat, N.; Resh, V.H. Benthic macro invertebrate assemblages and macrohabitat connectivity in Mediterranean-climate streams of northern California. J. N. Am. Benthol. Soc. 2006, 25, 32–43. [Google Scholar] [CrossRef]
- Bonada, N.; Rieradevall, M.; Prat, N. Macroinvertebrate community structure and biological traits related to flow permanence in a Mediterranean river network. Hydrobiologia 2007, 589, 91–106. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Dias-Silva, K.; Teodósio, M.A.; Juen, L. The response of neotropical dragonflies (insecta: Odonata) to local and regional abiotic factors in small streams of the amazon. Insects 2019, 10, 446. [Google Scholar] [CrossRef] [Green Version]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; De Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg City, Luxembourg, 2010. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://doi.org/10.2305/IUCN.UK.2010-1.RLTS.T158700A5263990.en (accessed on 16 November 2021).
- Biudes, J.F.V.; Camargo, A.F.M. Studying the limitant factors to primary production of aquatic macrophytes in Brazil. (Estudos dos Fatores Limitantes à Produção Primária por Macrófitas Aquáticas no Brasil). Oecol. Aust. 2008, 12, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Buffagni, A.; Erba, S.; Armanini, D.G. The lentic–lotic character of Mediterranean rivers and its importance to aquatic invertebrate communities. Aquat. Sci. 2010, 72, 45–60. [Google Scholar] [CrossRef]
- Kemp, J.L.; Harper, D.M.; Crosa, G.A. Use of ‘functional habitats’ to link ecology with morphology and hydrology in river rehabilitation. Aquat. Conserv. Mar. Freshw. Ecosyst. 1999, 9, 159–178. [Google Scholar] [CrossRef]
- Pitlo, R.H.; Dawson, F.H. Flow-resistance of aquatic weeds. In Aquatic Weeds, The Ecology and Management of Nuisance Aquatic Vegetation, 1st ed.; Pieterse, A.H., Murphy, K.J., Eds.; Oxford University Press: New York, NY, USA, 1990; pp. 74–84. [Google Scholar]
- Lemly, A.D.; Hilderbrand, R.H. Influence of large woody debris on stream insect communities and benthic detritus. Hydrobiologia 2000, 421, 179–185. [Google Scholar] [CrossRef]
- Johnson, W.C. Tree recruitment and survival in rivers: Influence of hydrological processes. Hydrol. Process 2000, 14, 3051–3074. [Google Scholar] [CrossRef]
- Corbet, P.; Brooks, S. Dragonflies, Collins New Naturalist Library No 106; Collins: London, UK, 2008. [Google Scholar]
- Perron, M.A.C.; Pick, F.R. Stormwater ponds as habitat for Odonata in urban areas: The importance of obligate wetland plant species. Biodivers. Conserv. 2020, 29, 913–931. [Google Scholar] [CrossRef]
- Perron, M.A.C.; Richmond, I.C.; Pick, F.R. Plants, water quality and land cover as drivers of Odonata assemblages in urban ponds. Sci. Total Environ. 2021, 773, 145467. [Google Scholar] [CrossRef]
- Daso, J.M.; Arquisal, I.B.; Yuto, C.M.M.; Mondejar, E.P. Species diversity of Odonata in Bolyok Falls, Naawan, Misamis Oriental, Philippines. Aquac. Aquar. Conserv. Legis. 2021, 14, 664–671. [Google Scholar]
- Holtmann, L.; Brüggeshemke, J.; Juchem, M.; Fartmann, T. Odonate assemblages of urban stormwater ponds: The conservation value depends on pond type. J. Insect Conserv. 2019, 23, 123–132. [Google Scholar] [CrossRef]
- Johansson, F.; Brodin, T. Effects of fish predators and abiotic factors on dragonfly community structure. J. Freshw. Ecol. 2003, 18, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Kriska, G. Dragonflies and damselflies—Odonata. In Freshwater Invertebrates in Central Europe, 1st ed.; Springer: Vienna, Austria, 2013; pp. 194–209. [Google Scholar] [CrossRef]
- Boda, R.; Bereczki, C.; Ortmann-Ajkai, A.; Mauchart, P.; Pernecker, B.; Csabai, Z. Emergence behaviour of the red listed Balkan Goldenring (Cordulegaster heros Theischinger, 1979) in Hungarian upstreams: Vegetation structure affects the last steps of the larvae. J. Insect Conserv. 2015, 19, 547–557. [Google Scholar] [CrossRef]
- Wildermuth, H. Habitat selection and oviposition site recognition by the dragonfly Aeshna juncea (L.): An experimental approach in natural habitats (Anisoptera: Aeshnidae). Odonatologica 1993, 22, 27–44. [Google Scholar]
- Purse, B.V.; Thompson, D.J. Oviposition site selection by Coenagrion mercuriale (Odonata: Coenagrionidae). Int. J. Odonatol. 2009, 12, 257–273. [Google Scholar] [CrossRef]
- Wildermuth, H. Dragonflies recognize the water of rendezvous and oviposition sites by horizontally polarized light: A behavioural field test. Sci. Nat. 1998, 85, 297–302. [Google Scholar] [CrossRef]
- Ward, L.; Mill, P.J. Habitat factors influencing the presence of adult Calopteryx splendens (Odonata: Zygoptera). Eur. J. Entomol. 2005, 102, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Steytler, N.S.; Samways, M.J. Biotope selection by adult male dragonflies (Odonata) at an artifical lake created for insect conservation in South Africa. Biol. Conserv. 1995, 72, 381–386. [Google Scholar] [CrossRef]
- McPeek, M.A. Ecological factors limiting the distributions and abundances of Odonata. In Dragonflies: Model Organisms for Ecological and Evolutionary Research, 1st ed.; Córdoba-Aguilar, A., Ed.; Oxford University Press: Oxford, UK, 2008; pp. 51–62. [Google Scholar]
- Sanchez-Montoya, M.D.; Punti, T.; Suarez, M.L.; Vidal-Abarca, M.D.; Rieradevall, M.; Poquet, J.M.; Zamora-Munoz, C.; Robles, S.; Alvarez, M.; Alba-Tercedor, J.; et al. Concordance between ecotypes and macroinvertebrate assemblages in Mediterranean streams. Freshw. Biol. 2007, 52, 2240–2255. [Google Scholar] [CrossRef]
- Pernecker, B.; Mauchart, P.; Csabai, Z. What to do if streams go dry? Behaviour of Balkan Goldenring (Cordulegaster heros, Odonata) larvae in a simulated drought experiment in SW Hungary. Ecol. Entomol. 2020, 45, 1457–1465. [Google Scholar] [CrossRef]
- Petrovičová, K.; Langraf, V.; David, S.; Krumpálová, Z.; Schlarmannová, J. Distinct Odonata assemblage variations in lentic reservoirs in Slovakia (Central Europe). Biologia 2021. [Google Scholar] [CrossRef]
- Veech, J.A.; Summerville, K.S.; Crist, T.O.; Gering, J.C. The additive partitioning of species diversity: Recent revival of an old idea. Oikos 2002, 99, 3–9. [Google Scholar] [CrossRef] [Green Version]







| Habitat Type | ||
|---|---|---|
| Macrophyte Poor | Macrophyte Rich | |
| Vascular Plants | ||
| Dominant vascular plants | Mentha longifolia (L.) L., Oenanthe fistulosa L. | Phragmites australis (Cav.) Steud., Scirpus lacustris L., Cyperus longus L. |
| Species richness (total, mean ± SE) | 1.00 ± 0.45 b | 10.50 ± 1.61 a |
| Abundance (total, mean ± SE) | 3.33 ± 1.69 b | 39.00 ± 5.72 a |
| Species richness of LM (mean ± SE) | 1.00 ± 0.45 b | 7.50 ± 1.09 a |
| Abundance of LM (mean ± SE) | 3.33 ± 1.69 b | 25.00 ± 3.52 a |
| Species richness of HM (mean ± SE) | 0.00 b | 2.83 ± 0.70 a |
| Abundance of HM (mean ± SE) | 0.00 b | 14.00 ± 2.68 a |
| Bryophytes | ||
| Dominant bryophytes | Cinclidotus aquaticus (Hedw.) Bruch et Schimp., C. fontinaloides (Hedw.) P. Beauv., Rhynchostegium riparioides (Hedw.) Cardot, Cratoneuron filicinum (Hedw.) Spruce | Cinclidotus fontinaloides (Hedw.) P. Beauv., Calliergonella cuspidata (Hedw.) Loseke, Fontinalis antipyretica Hedw. |
| Species richness (total, mean ± SE) | 3.33 ± 0.33 a | 1.50 ± 0.81 a |
| Abundance (mean ± SE) | 14.17 ± 1.25 a | 3.83 ± 2.17 b |
| Environmental Variables | MPH | MRH | F | p | d.f. 1 | d.f. 2 |
|---|---|---|---|---|---|---|
| (Mean ± SE) | (Mean ± SE) | |||||
| Alkalinity (mg CaCO3/L) | 126.94 ± 3.75 | 157.50 ± 2.63 | 23.589 | 0.000 | 1 | 34 |
| Water hardness (mg CaCO3/L) | 224.48 ± 14.18 | 277.88 ± 10.78 | 10.201 | 0.003 | 1 | 34 |
| Conductivity (μS/cm) | 414.61 ± 20.04 | 547.72 ± 16.80 | 10.224 | 0.003 | 1 | 34 |
| Water temperature (°C) | 10.84 ± 0.62 | 14.09 ± 0.38 | 6.706 | 0.014 | 1 | 34 |
| Water velocity (m/s) | 0.58 ± 0.03 | 0.31 ± 0.06 | 7.667 | 0.009 | 1 | 34 |
| Oxygen saturation (%) | 98.89 ± 1.22 | 107.81 ± 2.53 | 2.790 | 0.104 | 1 | 34 |
| Nitrates (mg N/L) | 0.25 ± 0.04 | 0.18 ± 0.02 | 1.085 | 0.305 | 1 | 34 |
| Water depth (cm) | 27.22 ± 4.15 | 33.61 ± 4.42 | 0.985 | 0.328 | 1 | 34 |
| Nitrites (mg N/L) | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.481 | 0.495 | 1 | 34 |
| Chemical oxygen demand (mg/L) | 3.61 ± 0.23 | 3.76 ± 0.37 | 0.315 | 0.578 | 1 | 34 |
| o-phosphates (mg N/L) | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.208 | 0.652 | 1 | 34 |
| Dissolved oxygen (mg/L) | 10.80 ± 0.19 | 11.04 ± 0.25 | 0.176 | 0.678 | 1 | 34 |
| pH | 7.98 ± 0.08 | 7.96 ± 0.03 | 0 | 0.988 | 1 | 34 |
| Habitat Type | Total | ||||||
|---|---|---|---|---|---|---|---|
| Macrophyte Poor | Macrophyte Rich | ||||||
| Species Name | Species Code | N | % | N | % | N | % |
| Calopteryx splendens (Harris, 1782) | Ca spl | 25 | 3.85 | 233 | 3.75 | 258 | 3.76 |
| Calopteryx virgo (Linnaeus, 1758) | Ca vir | 564 | 86.80 | 70 | 1.13 | 634 | 9.24 |
| Chalcolestes viridis (Vander Linden, 1825) | Ch vir | 675 | 10.86 | 675 | 9.84 | ||
| Sympecma fusca (Vander Linden, 1820) | Sy fus | 222 | 3.58 | 222 | 3.23 | ||
| Ischnura elegans (Van der Linden, 1820) | Is ele | 370 | 5.96 | 370 | 5.38 | ||
| Coenagrion puella (Linnaeus, 1758) | Co pue | 392 | 6.31 | 392 | 5.71 | ||
| Erythromma lindenii (Selys, 1840) | Er lin | 218 | 3.51 | 218 | 3.18 | ||
| Platycnemis pennipes (Pallas, 1771) | Pl pen | 20 | 3.08 | 3500 | 56.33 | 3520 | 51.28 |
| Aeshna affinis Vander Linden, 1820 | 1 | 0.15 | 4 | 0.07 | 5 | 0.07 | |
| Aeshna isoceles (Müller, 1767) | Ae iso | 31 | 0.50 | 31 | 0.45 | ||
| Anax imperator (Selys, 1839) | An imp | 30 | 0.48 | 30 | 0.44 | ||
| Brachytron pratense (Müller, 1764) | Br pra | 34 | 0.55 | 34 | 0.50 | ||
| Onychogomphus forcipatus (Linnaeus, 1758) | On for | 36 | 5.54 | 51 | 0.82 | 87 | 1.28 |
| Cordulegaster heros Theischinger, 1979 | 3 | 0.46 | 3 | 0.04 | |||
| Somatochlora meridionalis Nielsen, 1935 | 1 | 0.15 | 18 | 0.29 | 19 | 0.28 | |
| Libellula depressa Linnaeus, 1758 | Li dep | 25 | 0.40 | 25 | 0.36 | ||
| Libellula fulva Müller, 1764 | Li ful | 45 | 0.72 | 45 | 0.66 | ||
| Orthetrum cancellatum (Linnaeus, 1758) | 17 | 0.27 | 17 | 0.25 | |||
| Orthetrum coerulescens (Fabricius, 1798) | Or coe | 102 | 1.64 | 102 | 1.49 | ||
| Orthetrum brunneum (Fonscolombe, 1837) | Or bru | 48 | 0.77 | 48 | 0.70 | ||
| Sympetrum sanguineum (Müller, 1764) | 4 | 0.06 | 4 | 0.06 | |||
| Sympetrum fonscolombii (Selys, 1840) | 4 | 0.06 | 4 | 0.06 | |||
| Sympetrum striolatum (Charpentier, 1840) | Sy str | 105 | 1.69 | 105 | 1.53 | ||
| Sympetrum meridionale (Selys, 1841) | 1 | 0.02 | 1 | 0.01 | |||
| Crocothemis erythraea (Brullé, 1832) | 14 | 0.23 | 14 | 0.20 | |||
| Species richness (S) | 7 | 24 | 25 | ||||
| Abundance (N) | 650 | 6213 | 6863 | ||||
| Assemblage Parameter | F | p | d.f. 1 | d.f. 2 | |
|---|---|---|---|---|---|
| Species richness (S) | 45.756 | 0.000 | 1 | 34 | |
| Abundance (N) | 58.940 | 0.000 | 1 | 34 | |
| Shannon diversity (H′) | 29.200 | 0.000 | 1 | 23 | |
| Simpson diversity (1 − λ) | 19.700 | 0.000 | 1 | 23 | |
| Functional Parameter | |||||
| Functional diversity (RaoQ) | 28.563 | 0.000 | 1 | 27 | |
| CWM body size | 8.149 | 0.008 | 1 | 27 | |
| CWM hydrological preferences | eupotamon | 182.582 | 0.000 | 1 | 27 |
| parapotamon | 13.839 | 0.001 | 1 | 21 | |
| plesiopotamon | 91.216 | 0.000 | 1 | 22 | |
| palaeopotamon | 19.457 | 0.000 | 1 | 22 | |
| temporary water bodies | 7.247 | 0.014 | 1 | 19 | |
| CWM nymphal development | spring | 20.020 | 0.000 | 1 | 17 |
| summer | 128.804 | 0.000 | 1 | 17 | |
| autumn | - | - | - | - | |
| winter | - | - | - | - | |
| all year | 188.086 | 0.000 | 1 | 27 | |
| CWM drought resilience form | no drought resilience | 138.067 | 0.000 | 1 | 26 |
| egg diapause | 11.703 | 0.003 | 1 | 17 | |
| nymph diapause | 1.994 | 0.178 | 1 | 15 | |
| adult diapause | - | - | - | - | |
| unknown resilience type | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilenica, M.; Rebrina, F.; Matoničkin Kepčija, R.; Šegota, V.; Rumišek, M.; Ružanović, L.; Brigić, A. Aquatic Macrophyte Vegetation Promotes Taxonomic and Functional Diversity of Odonata Assemblages in Intermittent Karst Rivers in the Mediterranean. Diversity 2022, 14, 31. https://doi.org/10.3390/d14010031
Vilenica M, Rebrina F, Matoničkin Kepčija R, Šegota V, Rumišek M, Ružanović L, Brigić A. Aquatic Macrophyte Vegetation Promotes Taxonomic and Functional Diversity of Odonata Assemblages in Intermittent Karst Rivers in the Mediterranean. Diversity. 2022; 14(1):31. https://doi.org/10.3390/d14010031
Chicago/Turabian StyleVilenica, Marina, Fran Rebrina, Renata Matoničkin Kepčija, Vedran Šegota, Mario Rumišek, Lea Ružanović, and Andreja Brigić. 2022. "Aquatic Macrophyte Vegetation Promotes Taxonomic and Functional Diversity of Odonata Assemblages in Intermittent Karst Rivers in the Mediterranean" Diversity 14, no. 1: 31. https://doi.org/10.3390/d14010031