
Self-Compatibility and Reproductive Success of Oenothera drummondii subsp. drummondii: Is It Similar between Native and Non-Native Populations?



Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Studied
2.2. Seed Collection and Plant Material
2.3. Pollination Treatments and Fruit and Seed Data
2.4. Germination Experiment
2.5. Data Analysis
3. Results
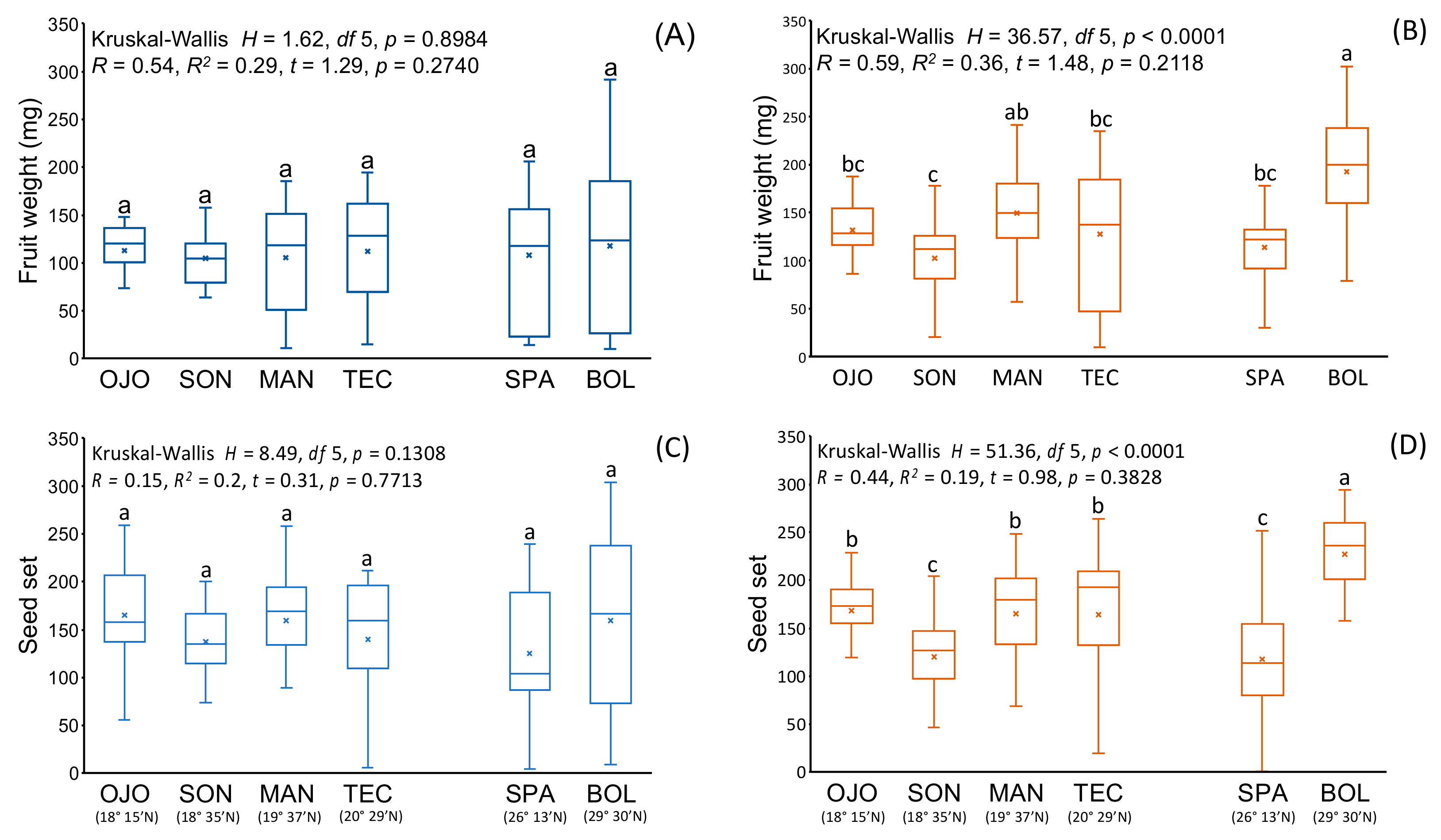
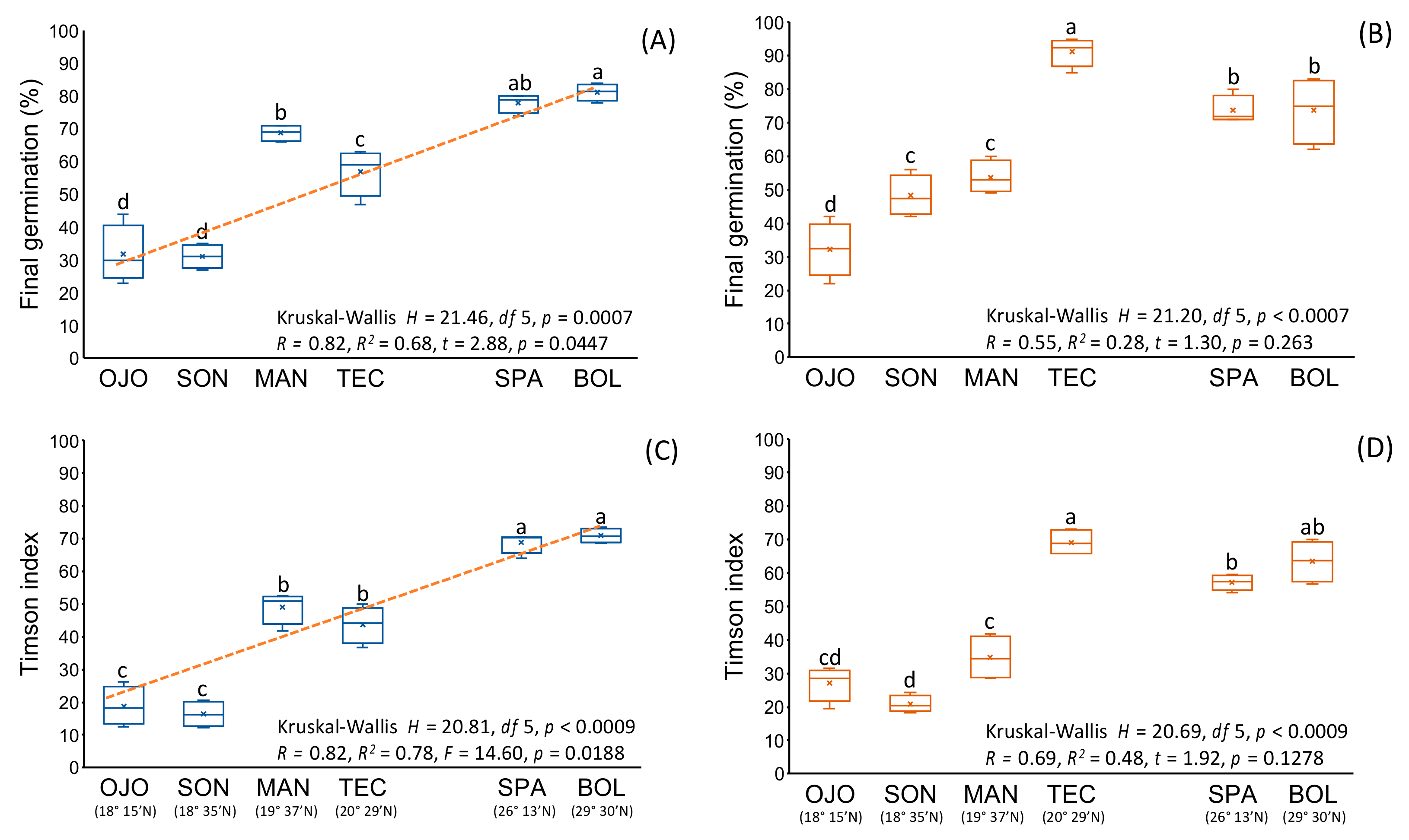
3.1. Compatibility, Fruit and Seed Characteristics among Native Populations
3.2. Fruit and Seed Characteristics of Non-Native Populations
3.3. Comparison of Reproductive Characteristics between Native and Non-Native Populations
3.4. Integral Reproductive Success Index
4. Discussion
4.1. Mating System and Fruit and Seed Characteristics in Native Populations
4.2. Fruit and Seed Characteristics of Native Populations
4.3. Comparison between Native and Non-Native Populations
4.4. Integral Reproductive Success Index (IRSI)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Country | Population | Acronym | Latitude | Longitude | |
|---|---|---|---|---|---|---|
| 1 | Native | Mexico | Ojoshal | OJO | 18°15′ N | 93°59′ W |
| 2 | Native | Mexico | Sontecomapan | SON | 18°33′ N | 94°59′ W |
| 3 | Native | Mexico | La Mancha | MAN | 19°37′ N | 96°22′ W |
| 4 | Native | Mexico | Tecolutla | TEC | 20°29′ N | 97°01′ W |
| 5 | Native | USA | South Padre | SPA | 26°13′ N | 97°11′ W |
| 6 | Native | USA | Bolivar | BOL | 29°30′ N | 94°30′ W |
| 7 | Non-native | Israel | Rishon-Lezion | LEZ | 31°59′ N | 34°43′ W |
| 8 | Non-native | Australia | Mandurah | MAH | 32°32′ S | 115°41′ E |
| 9 | Non-native | South Africa | Muizenberg | MUI | 34°06′ S | 18°28′ E |
| 10 | Non-native | Spain | Dique | DIQ | 37°09′ N | 06°54′ W |
| Chracteristics of Fruits and Seeds | Pollination Treatment | Native Populations | Non-Native Populations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OJO | SON | MAN | TEC | SPA | BOL | LEZ | MAH | MUI | DIQ | ||
| Spontaneous | 0 | 12 | 20 | 15 | 2 | 19 | 5 | 4 | 0 | 7 | |
| Fruit set (%) | Self | 96 | 100 | 96 | 96 | 100 | 92 | 96 | 97 | 100 | 100 |
| Cross | 92 | 100 | 96 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
| Self-Compatibility Index | 1.04 | 1.00 | 1.00 | 0.96 | 1.00 | 0.92 | 0.96 | 0.97 | 1.00 | 1.00 | |
| Spontaneous | 11.4 ± 2.1 | 18.1 ± 14.6 | 34.2 ± 29.4 | 24.3 ± 24.1 | 22.9 ± 17.2 | 35.8 ± 48.6 | 18.0 ± 12.2 | 22.9 ± 5.3 | 11.9 ± 12.9 | 17.8 ± 24.9 | |
| Fruit weight (mg) | Self | 112.7 ± 28.5 | 104.6 ± 37.6 | 105.6 ± 56.8 | 112.4 ± 56.9 | 107.8 ± 68.5 | 117.5 ± 85.0 | 112.6 ± 64.0 | 139.0 ± 81.7 | 97.7 ± 35.5 | 109.0 ± 48.7 |
| Cross | 131.3 + 51.4 | 102.8 + 41.5 | 149.2 + 50.7 | 127.9 + 70.9 | 113.8 + 44.0 | 192.8 ± 66.5 | 96.3 ± 61.6 | 176.1 ± 67.5 | 116.6 ± 37.3 | 84.4 ± 56.6 | |
| ns | ns | *** | ns | ns | *** | ns | ns | * | ns | ||
| Spontaneous | 0.0 ± 0.0 | 46.7 ± 38.8 | 42.0 ± 36.6 | 54.8 ± 83.2 | 62.0 ± 35.4 | 89.8 ± 109.9 | 58.0 ± 0.0 | 10.0 ± 0.0 | 0.0 ± 0.0 | 72 ± 100.4 | |
| Seed set | Self | 165.5 ± 47.1 | 137.7 ± 34.7 | 159.6 ± 58.7 | 139.7 ± 61.8 | 137.8 ± 66.1 | 159.8 ± 95.2 | 215.7 ± 59.0 | 165.6 ± 96.8 | 161.5 ± 35.8 | 190.5 ± 60.3 |
| Cross | 168.4 ± 67.5 | 120.2 ± 44.7 | 164.9 ± 65.4 | 164.6 ± 75.4 | 117.9 ± 62.8 | 227.1 ± 45.3 | 176.8 ± 94.8 | 175.2 ± 87.8 | 173.2 ± 49.7 | 161.7 ± 83.3 | |
| ns | ns | ns | ns | ns | ** | ns | ns | ns | ns | ||
| Pollination Treatment | Native Populations | Non-Native Populations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OJO | SON | MAN | TEC | SPA | BOL | LEZ | MAH | MUI | DIQ | ||
| First, germination (d) | Self | 4.0 ± 0.0 | 4.5 ± 1.0 | 5.0 ± 0.8 | 5.5 ± 1.0 | 5.0 ± 1.2 | 4.0 ± 0.0 | 4.5 ± 1.0 | 4.0 ± 0.0 | 4.0 ± 0.0 | 4.0 ± 0.0 |
| Cross | 4.0 ± 0.0 | 7.3 ± 1.5 | 5.0 ± 0.8 | 4.0 ± 0.0 | 5.75 ± 0.50 | 4.0 ± 0.0 | 4.0 ± 0.0 | 5.0 ± 1.2 | 4.0 ± 0.0 | 5.3 ± 1.5 | |
| ns | * | ns | * | ns | ns | ns | ns | ns | ns | ||
| Timson Index | Self | 18.8 ± 5.9 | 16.3 ± 3.9 | 49.0 ± 4.9 | 43.7 ± 5.6 | 68.8 ± 3.2 | 70.9 ± 2.2 | 61.6 ± 3.6 | 65.2 ± 1.8 | 62.9 ± 3.7 | 54.5 ± 15.4 |
| Cross | 27.1 ± 5.3 | 20.9 ± 2.5 | 43.8 ± 6.7 | 69.2 ± 3.8 | 57.2 ± 2.3 | 63.6 ± 6.2 | 72.0 ± 3.8 | 87.6 ± 1.6 | 76.7 + 4.5 | 67.6 ± 12.9 | |
| ns | ns | * | * | * | ns | * | * | * | ns | ||
| Final germination (%) | Self | 31.8 ± 8.9 | 31.0 ± 3.7 | 68.8 ± 2.6 | 57.0 ± 7.1 | 78.0 ± 2.8 | 81.3 ± 2.5 | 73.0 ± 4.3 | 72.8 ± 2.1 | 77.3 ± 4.4 | 62.8 ± 15.5 |
| Cross | 32.3 ± 8.2 | 48.3 ± 6.1 | 53.8 ± 4.9 | 91.25 ± 4.4 | 73.8 ± 9.9 | 73.8 ± 9.9 | 79.8 ± 4.9 | 95.3 ± 1.9 | 85.3 ± 3.9 | 78.3 ± 15.8 | |
| ns | * | * | * | ns | ns | ns | * | * | ns | ||
| Integral Reproductive Success | Self | 0.23 | 0.20 | 0.49 | 0.35 | 0.50 | 0.50 | 0.70 | 0.54 | 0.58 | 0.55 |
| Index (IRSI) | Cross | 0.22 | 0.26 | 0.38 | 0.66 | 0.38 | 0.74 | 0.62 | 0.74 | 0.65 | 0.56 |
References
- Pierre-Olivier, C. The Evolution of Plant Mating System: Is It Time for a Synthesis? In Studies in Population Genetics; Fusté, M.C., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 17–38. [Google Scholar]
- Whitehead, M.R.; Lanfear, R.; Mitchell, R.J.; Karron, J.D. Plant mating systems often vary widely among populations. Front. Ecol. Evol. 2018, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Barrett, S.C.H. Mating strategies in flowering plants: The outcrossing–selfing paradigm and beyond. Philos. Transect. R. Soc. B 2003, 358, 991–1004. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Harder, L.D. The ecology of mating and its evolutionary consequences in seed plants. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 135–157. [Google Scholar] [CrossRef] [Green Version]
- Goodwillie, C.; Kalisz, S.; Eckert, C.G. The evolutionary enigma of mixed mating systems in plants: Occurrence, theoretical explanations, and empirical evidence. Ann. Rev. Eco. Evol. Syst. 2005, 36, 47–79. [Google Scholar] [CrossRef] [Green Version]
- Gamble, D.E.; Bontrager, M.; Angert, A.L. Floral trait variation and links to climate in the mixed-mating annual Clarkia pulchella. Botany 2018, 96, 425–435. [Google Scholar] [CrossRef]
- Antoń, S.; Denisow, B. Pollination biology and breeding system in five nocturnal species of Oenothera (Onagraceae): Reproductive assurance and opportunities for outcrossing. Plant Syst. Evol. 2018, 304, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- Randle, A.M.; Slyder, J.B.; Kalisz, S. Can differences in autonomous selfing ability explain differences in range size among sister-taxa pairs of Collinsia (Plantaginaceae)? An extension of Baker’s Law. New Phytol. 2009, 183, 618–629. [Google Scholar] [CrossRef]
- Munoz, F.; Violle, C.; Cheptou, P.-O. CSR ecological strategies and plant mating systems: Outcrossing increases with competitiveness but stress-tolerance is related to mixed mating. Oikos 2016, 125, 1296–1303. [Google Scholar] [CrossRef]
- Grossenbacher, D.L.; Runquist, R.; Goldberg, E.; Brandvain, Y. Geographic range size is predicted by plant mating system. Ecol. Lett. 2015, 18, 706–713. [Google Scholar] [CrossRef]
- Razanajatovo, M.; Maurel, N.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Pyšek, P.; Weigelt, P.; Winter, M.; van Kleunen, M. Plants capable of selfing are more likely to become naturalized. Nat. Commun. 2016, 7, 13313. [Google Scholar] [CrossRef] [Green Version]
- Eckert, C.; Kalisz, S.; Geber, M.A.; Sargent, R.; Elle, E.; Cheptou, P.O.; Goodwillie, C.; Johnston, M.O.; Kelly, J.K.; Moeller, D.A.; et al. Plant mating systems in a changing world. Trends Ecol. Evol. 2009, 25, 35–43. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Eckert, C.G. Evolution of dispersal and mating systems along geographic gradients: Implications for shifting ranges. Funct. Ecol. 2014, 28, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions, Section II; Nentwig, W., Ed.; Springer: Berlin, Germany, 2007; pp. 97–125. [Google Scholar]
- Van Kleunen, M.; Bossdorf, O.; Dawson, W. The ecology and evolution of alien plants. Ann. Rev. Ecol. Evol. Syst. 2018, 49, 25–47. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Pišek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Baker, H.G. Self-compatibility and establishment after ‘long-distance’ dispersal. Evolution 1955, 9, 347–348. [Google Scholar] [CrossRef]
- Cheptou, P.O. Does the evolution of self-fertilization rescue populations or increase the risk of extinction? Ann. Bot. 2019, 123, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Van Boheemen, L.A.; Atwater, D.Z.; Hodgins, K.A. Rapid and repeated local adaptation to climate in an invasive plant. New Phytol. 2019, 222, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Stout, J.C. Pollination of invasive Rhododendron ponticum (Ericaceae) in Ireland. Apidologie 2007, 38, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Watts, S.; Connerty, S.; Lock, J.; Parker, L.; Wilson, I.; Schueller, S.; Nattero, J.; Cocucci, A.A.; Izhaki, I.; et al. Pollination ecology of the invasive tree tobacco Nicotiana glauca: Comparisons across native and non-native ranges. J. Pollinat. Ecol. 2012, 9, 85–95. [Google Scholar] [CrossRef]
- Petanidou, T.; Godfree, R.C.; Song, D.S.; Kantsa, A.; Dupont, Y.L.; Waser, N. Self-compatibility and plant invasiveness: Comparing species in native and invasive ranges. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 3–12. [Google Scholar] [CrossRef]
- Issaly, E.A.; Sérsic, A.N.; Pauw, A.; Cocucci, A.A.; Travest, A.; Benítez-Vieyra, S.M.; Paiaro, V. Reproductive ecology of the bird-pollinated Nicotiana glauca across native and introduced ranges with contrasting pollination environments. Biol. Invasions 2020, 22, 485–498. [Google Scholar] [CrossRef]
- Dietrich, W.; Wagner, W.L. Systematics of Oenothera Section Oenothera Subsection Raimannia and Subsection Nutantigemma (Onagraceae). Syst. Bot. Monogr. 1988, 24, 1–91. [Google Scholar] [CrossRef]
- Frean, M.; Balkwill, K.; Gold, C.; Burt, S. The expanding distributions and invasiveness of Oenothera in southern Africa. S. Afr. J. Bot. 1997, 63, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Heenan, P.B.; de Lange, P.J.; Cameron, E.K.; Champion, P.D. Checklist of dicotyledons, gymnosperms, and pteridophytes naturalised or casual in New Zealand: Additional records 1999–2000. N. Z. J. Bot. 2002, 40, 155–174. [Google Scholar] [CrossRef]
- Heyligers, P.C. Flora of the Stockton and Port Hunter sandy foreshores with comments on fifteen notable introduced species. Cunninghamia 2008, 10, 493–511. [Google Scholar]
- Gregory, D.P. Hawkmoth pollination in the Genus Oenothera. Aliso 1964, 5, 385–419. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Fernández, J.B.; Morales-Sánchez, J.A.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. Recovery of beach-foredune vegetation after disturbance by storms. J. Coast. Res. 2020, 95 (Suppl. S1), 34–38. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; (University of Sevilla, Sevilla, Andalucia, Spain); García-Franco, J.G.; Instituto de Ecología, A.C., Xalapa, Veracruz, Mexico). Personal communication, 2019.
- Theiss, K.E.; Holsinger, K.E.; Evans, M.E.K. Breeding system variation in 10 evening primroses (Oenothera sections Anogra and Kleinia; Onagraceae). Am. J. Bot. 2010, 97, 1031–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, P.H. A survey of reproductive biology in Onagraceae. N. Z. J. Bot. 1979, 17, 575–593. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, W.; Wagner, W.L.; Raven, P.H. Systematics of Oenothera Section Oenothera Subsection Oenothera (Onagraceae). Syst. Bot. Monogr. 1997, 50, 1–234. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; García-Franco, J.G. Floral trait variation in Oenothera drummondii subsp. drummondii across a wide latitudinal range of native and non-native populations. Flora 2021, 280, 151851. [Google Scholar] [CrossRef]
- Khan, M.A.; Ungar, I.A. The effect of salinity and temperature on the germination of polymorphic seeds and growth of Atriplex triangularis Willd. Am. J. Bot. 1984, 71, 481–489. [Google Scholar] [CrossRef]
- Sawma, J.T.; Mohler, C.L. Evaluating seed viability by an unimbued seed crush test in comparison with the tetrazolium test. Weed Technol. 2002, 16, 781–786. [Google Scholar] [CrossRef]
- Lloyd, D.G.; Schoen, D.J. Self-fertilization and cross-fertilization in plants. 1. Functional dimensions. Int. J. Plant Sci. 1992, 153, 358–369. [Google Scholar] [CrossRef]
- Vogler, D.W.; Kalisz, S. Sex among the flowers: The distribution of plant mating systems. Evolution 2001, 55, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Fornoni, J.; Ordano, M.; Pérez-Ishiwara, R.; Boege, K.; Domínguez, C.A. A comparison of floral integration between selfing and outcrossing species: A meta-analysis. Ann. Bot. 2016, 117, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Raguso, R.A.; Kelber, A.; Pfaff, M.; Levin, R.A.; McDade, L.A. Floral biology of North American Oenothera sect. Lavauxia (Onagraceae): Advertisements, rewards, and extreme variation in floral depth. Ann. Mo. Bot. Gard. 2007, 94, 236–257. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. Multiple seed dispersal modes of an invasive plant species on coastal dunes. Biol. Invasions 2021, 23, 111–127. [Google Scholar] [CrossRef]
- Ashman, T.L. Flower Longevity. In Plant Cell Death Processes; Nooden, L.D., Ed.; Elsevier Aademic Press: San Diego, CA, USA, 2004; pp. 349–362. [Google Scholar]
- Hernández-Espinosa, R.; González-Astorga, J.; Espinosa de los Monteros, A.; Cabrera-Toledo, D.; Gallego-Fernández, J.B. Transferability of microsatellite markers developed in Oenothera spp. to the invasive ipecies Oenothera drummondii Hook. (Onagraceae). Diversity 2020, 12, 387. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. The impact on plant communities of an invasive alien herb, Oenothera drummondii, varies along the beach-coastal dunes gradient. Flora 2019, 260, 151466. [Google Scholar] [CrossRef]
- Obeso, J.R.; Aedo, C. Plant-species richness and extinction on isolated dunes along the rocky coast of Northwestern Spain. J. Veg. Sci. 1992, 3, 129–132. [Google Scholar] [CrossRef]
- Hirayama, K.; Ishida, K.; Tomaru, N. Effects of pollen shortage and self-pollination on seed production of an endangered tree, Magnolia stellata. Ann. Bot. 2005, 95, 1009–1015. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Burd, M. The effect of pollen limitation on the evolution of mating system and seed size in hermaphroditic plants. Am. Nat. 2019, 193, 447–457. [Google Scholar] [CrossRef]
- Weller, S.G. The relationship of rarity to plant reproductive biology. In Restoration of Endangered Species; Bowles, M.L., Whelan, C.J., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 90–117. [Google Scholar]
- Weekley, C.W.; Race, T. The breeding system of Ziziphus celata Judd and D. W. Hall (Rhamnaceae), a rare endemic plant of the Lake Wales Ridge, Florida, USA: Implications for recovery. Biol. Conserv. 2001, 100, 207–213. [Google Scholar] [CrossRef]
- Eisikowitch, D.; Lazar, Z. Flower change in Oenothera drummondii Jooker as a response to pollinators’ visits. Bot. J. Linn. Soc. 1987, 95, 101–111. [Google Scholar] [CrossRef]
- Mihulka, A.; PysÏek, P. Invasion history of Oenothera congeners in Europe: A comparative study of spreading rates in the last 200 years. J. Biogeogr. 2001, 28, 597–609. [Google Scholar] [CrossRef]
- Ye, Z.-M.; Jin, X.-F.; Wang, Q.-F.; Yang, C.-F.; Inouye, D.W. Pollinators shift to nectar robbers when florivory occurs, with effects on reproductive success in Iris bulleyana (Iridaceae). Plant Biol. 2017, 19, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Herbertsson, L.; Jönsson, A.M.; Anderson, G.K.S.; Seibel, K.; Rubdiöf, M.; Ekroos, J.; Stjernman, M.; Olsson, O.; Smith, H.G. The impact of sown flower strips on plant reproductive success in Southern Sweden varies with landscape context. Agric. Ecosyst. Environ. 2018, 259, 127–134. [Google Scholar] [CrossRef]
- Sáyago, R.; Quesada, M.; Aguilar, R.; Ashworth, L.; Lopezaraiza-Mikel, M.; Martén-Rodríguez, S. Consequences of habitat fragmentation on the reproductive success of two Tillandsia species with contrasting life history strategies. AoB Plants 2018, 10, ply038. [Google Scholar] [CrossRef] [Green Version]
- Delnevo, N.; van Etten, E.J.; Byrne, M.; Stock, W.D. Floral display and habitat fragmentation: Effects on the reproductive success of the threatened mass-flowering Conospermum undulatum (Proteaceae). Ecol. Evol. 2019, 9, 11494–11503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickert, K.L.; O’Neal, E.S.; Davis, D.D.; Kasson, M.T. Seed production, viability, and reproductive limits of the invasive Ailanthus altissima (Tree-of-Heaven) within invaded environments. Forests 2017, 8, 226. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego-Fernández, J.B.; García-Franco, J.G. Self-Compatibility and Reproductive Success of Oenothera drummondii subsp. drummondii: Is It Similar between Native and Non-Native Populations? Diversity 2021, 13, 431. https://doi.org/10.3390/d13090431
Gallego-Fernández JB, García-Franco JG. Self-Compatibility and Reproductive Success of Oenothera drummondii subsp. drummondii: Is It Similar between Native and Non-Native Populations? Diversity. 2021; 13(9):431. https://doi.org/10.3390/d13090431
Chicago/Turabian StyleGallego-Fernández, Juan B., and José G. García-Franco. 2021. "Self-Compatibility and Reproductive Success of Oenothera drummondii subsp. drummondii: Is It Similar between Native and Non-Native Populations?" Diversity 13, no. 9: 431. https://doi.org/10.3390/d13090431