
Using Paleoecological Data to Inform the Conservation Strategy for Floristic Diversity and Isoetes taiwanensis in Northern Taiwan


Abstract
:1. Introduction
2. Materials and Methods
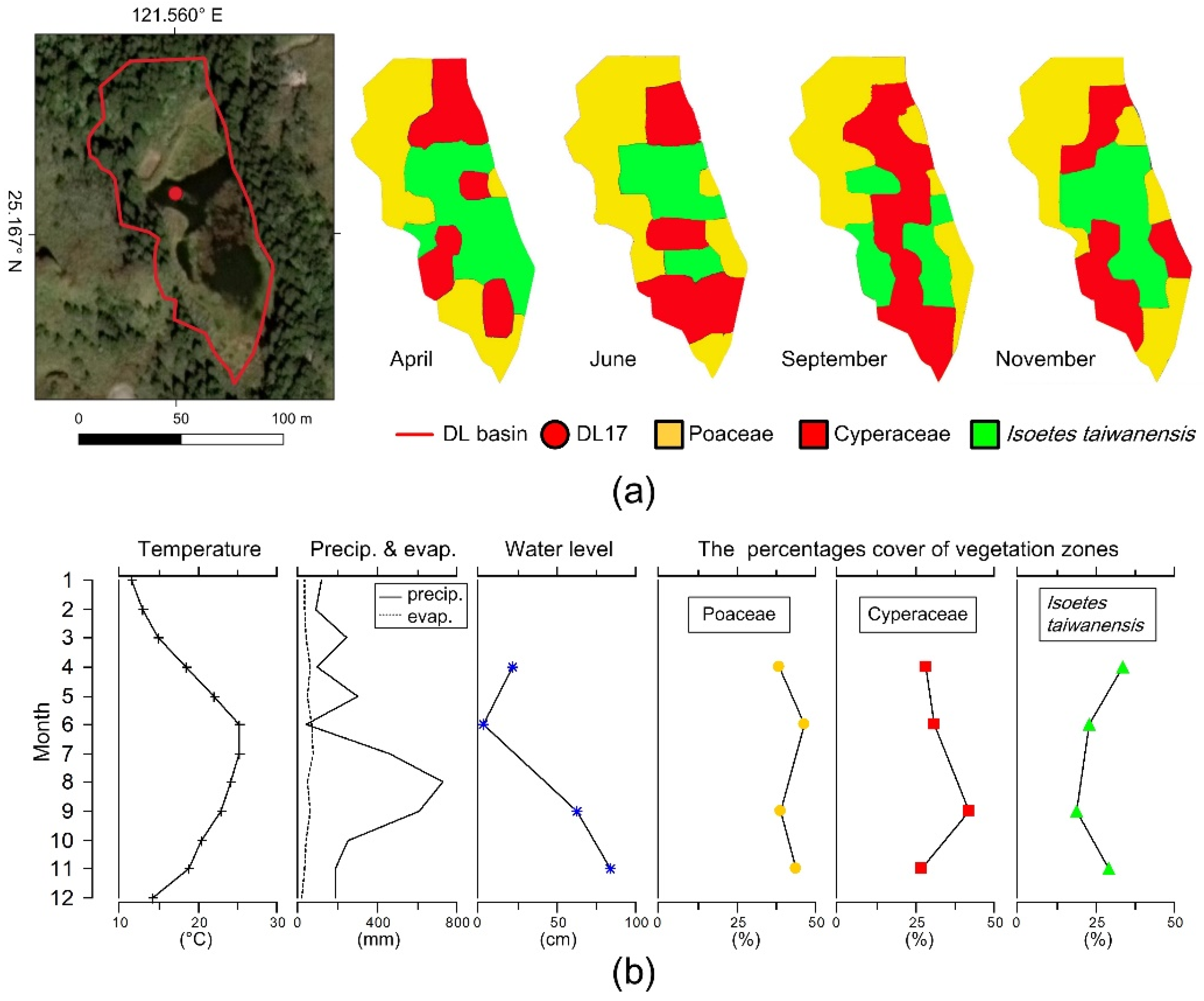
2.1. Study Area and Sampling
2.2. Multiproxy Analysis and Paleo-Floristic Diversity Assessment
3. Results
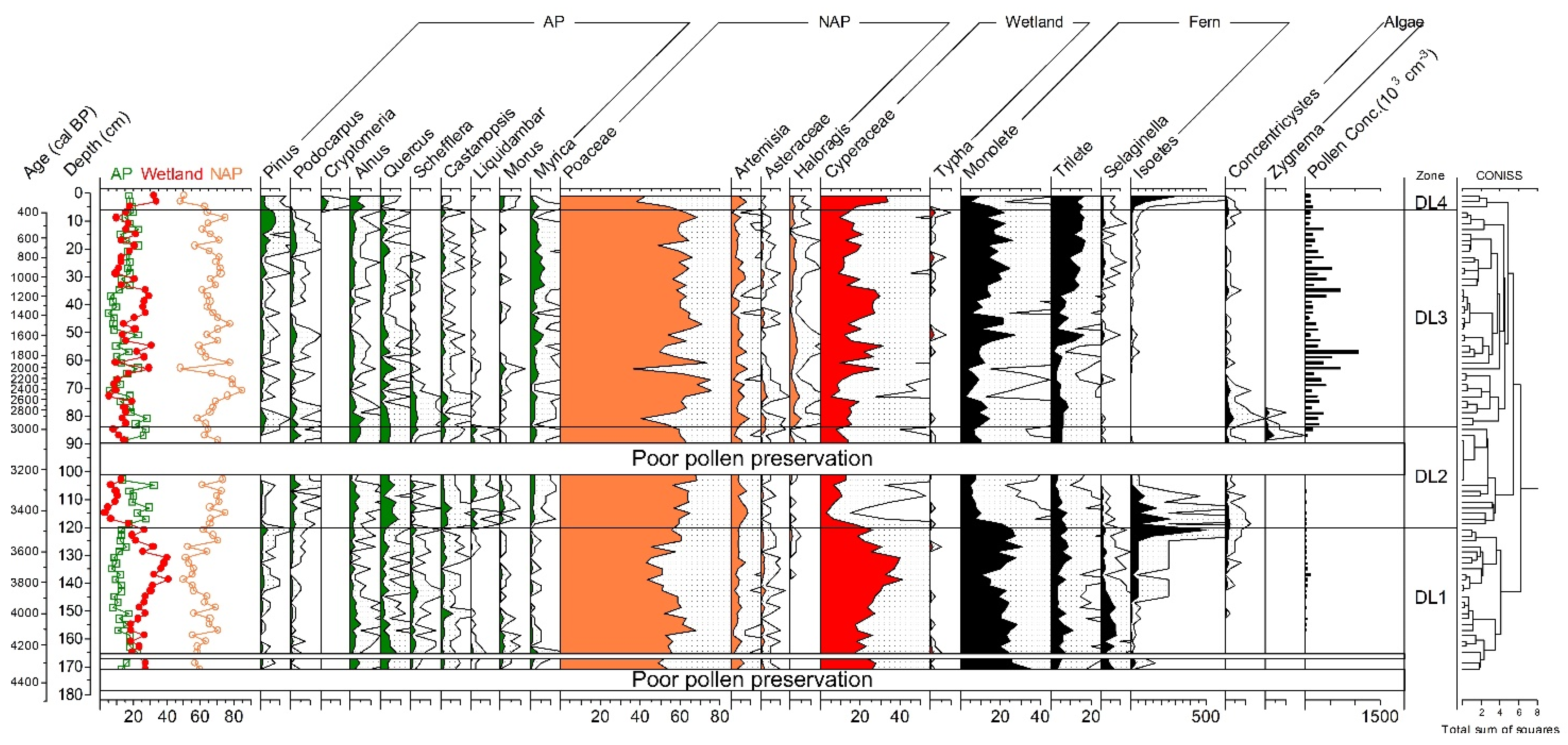
3.1. Core Description and Age–Depth Model Construction
3.2. Pollen, Charcoal, Biodiversity Estimation, and Ordination Results
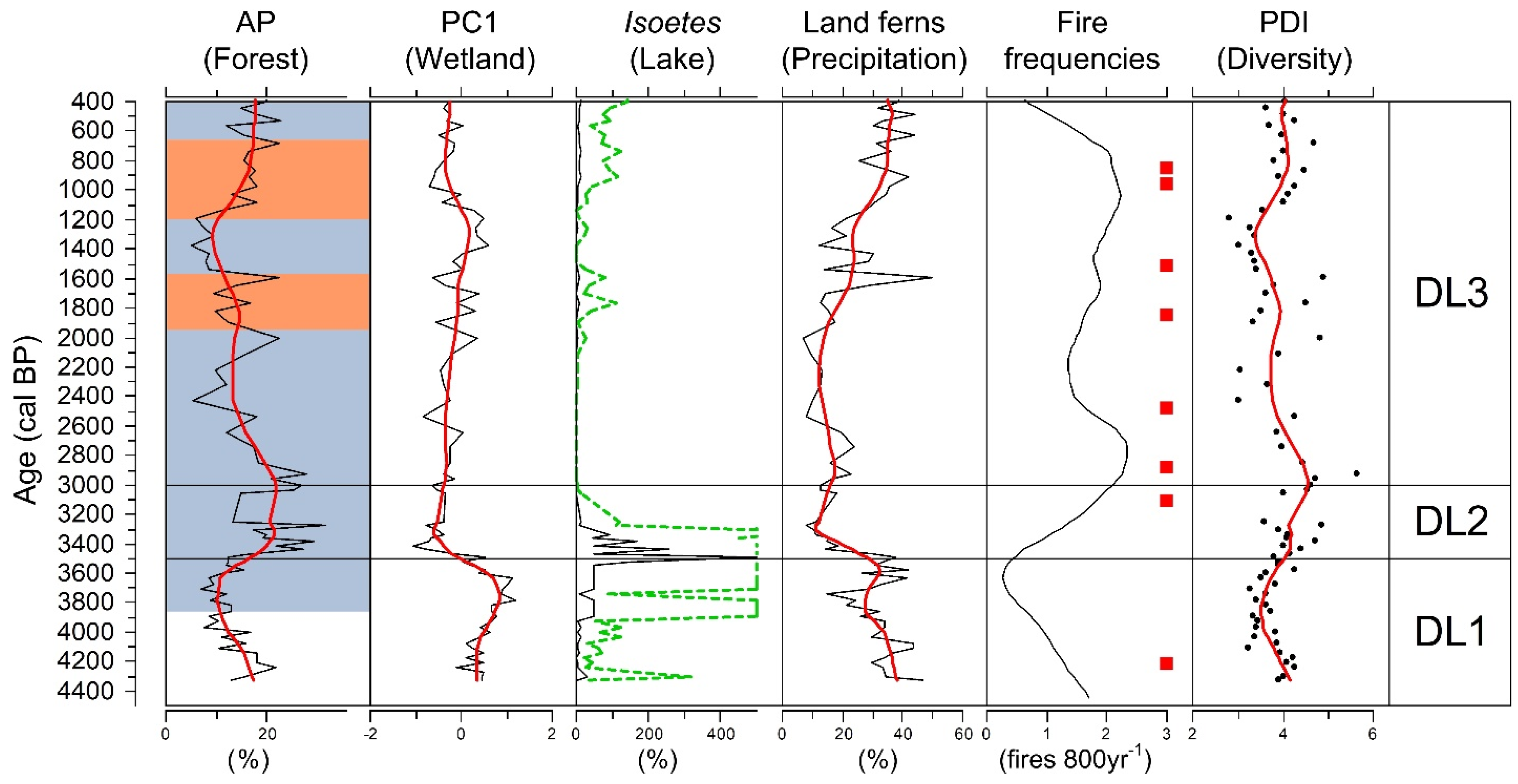
4. Discussion
4.1. Paleoenvironment Interpretation and Paleoclimate Reconstruction
4.2. Drivers of Local Vegetation Dynamics
4.3. Implications for Isoetes taiwanensis Conservation
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roleček, J.; Svitavská Svobodová, H.; Jamrichová, E.; Dudová, L.; Hájková, P.; Kletetschka, G.; Kuneš, P.; Abraham, V. Conservation Targets from the Perspective of a Palaeoecological Reconstruction: The Case Study of Dářko Peat Bog in the Czech Republic. Preslia 2020, 92, 87–114. [Google Scholar] [CrossRef]
- Lin, C.-H.; Shih, M.-H.; Lai, Y.-C. Mantle Wedge Diapirs Detected by a Dense Seismic Array in Northern Taiwan. Sci. Rep. 2021, 11, 1561. [Google Scholar] [CrossRef]
- Yu, Y.-T.; Lur, H.-S.; Chang, W.-L. Association of Water Depth and Aquatic-Plant Competition in Conservation of Isoetes Taiwanensis in the Menghuan Pond Wetland in Taiwan. Paddy Water Environ. 2013, 11, 513–519. [Google Scholar] [CrossRef]
- Chen, S.-H.; Wu, J.-T.; Yang, T.-N.; Chuang, P.-P.; Huang, S.-Y.; Wang, Y.-S. Late Holocene Paleoenvironmental Changes in Subtropical Taiwan Inferred from Pollen and Diatoms in Lake Sediments. J. Paleolimnol. 2009, 41, 315–327. [Google Scholar] [CrossRef]
- Huang, T.C.; Chen, H.J. The Pollen Analysis of the Dream Lake, Taipei, Yang Ming Shan National Park. J. Palynol. 1988, 23, 213–216. [Google Scholar]
- Huang, T.-C.; Chen, H.-J.; Li, L.-C. A Palynological Study of Isoëtes taiwanensis DeVol. Am. Fern J. 1992, 82, 142–150. [Google Scholar] [CrossRef]
- Wang, L.-C.; Chang, Y.-P.; Li, H.-C.; Chen, S.-H.; Wu, J.-T.; Lee, T.-Q.; Shiau, L.-J. Revealing the Vegetation, Fire and Human Activities in the Lowland of Eastern Taiwan during Late Holocene. Quat. Int. 2020, 544, 32–40. [Google Scholar] [CrossRef]
- Belousov, A.; Belousova, M.; Chen, C.-H.; Zellmer, G.F. Deposits, Character and Timing of Recent Eruptions and Gravitational Collapses in Tatun Volcanic Group, Northern Taiwan: Hazard-Related Issues. J. Volcanol. Geotherm. Res. 2010, 191, 205–221. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.H.; Huang, S.Y. Aeropalynological Study of Yangminshan National Park, Taiwan. Taiwania 2000, 45, 281–295. [Google Scholar]
- Lin, H.-J. Investigation and Monitoring of Habitats in Mengxiang Lake Ecological Reserve in Yangmingshan National Park; Yangmingshan National Park Headquarters: Taipei, Taiwan, 2015. [Google Scholar]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Blaauw, M.; Christen, J.A. Flexible Paleoclimate Age-Depth Models Using an Autoregressive Gamma Process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Huang, T.-C. Pollen Flora of Taiwan; National Taiwan University Botany Department Press: Taipei, Taiwan, 1972; p. 297. [Google Scholar]
- Huang, T.-C. Spore Flora of Taiwan; National Taiwan University Botany Department Press: Taipei, Taiwan, 1981; p. 111. [Google Scholar]
- Grimm, E.C. CONISS: A FORTRAN 77 Program for Stratigraphically Constrained Cluster Analysis by the Method of Incremental Sum of Squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Felde, V.A.; Bjune, A.E.; Grytnes, J.-A.; Seppä, H.; Giesecke, T. Does Pollen-Assemblage Richness Reflect Floristic Richness? A Review of Recent Developments and Future Challenges. Rev. Palaeobot. Palynol. 2016, 228, 1–25. [Google Scholar] [CrossRef]
- Liang, C.; Zhao, Y.; Qin, F.; Cui, Q.-Y.; Li, Q.; Li, H.; Zhang, Z.-Y. Complex Responses of Vegetation Diversity to Holocene Climate Change in the Eastern Tibetan Plateau. Veg. Hist. Archaeobotany 2019, 28, 379–390. [Google Scholar] [CrossRef]
- Matthias, I.; Semmler, M.S.S.; Giesecke, T. Pollen Diversity Captures Landscape Structure and Diversity. J. Ecol. 2015, 103, 880–890. [Google Scholar] [CrossRef]
- Gosling, W.D.; Julier, A.C.M.; Adu-Bredu, S.; Djagbletey, G.D.; Fraser, W.T.; Jardine, P.E.; Lomax, B.H.; Malhi, Y.; Manu, E.A.; Mayle, F.E.; et al. Pollen-Vegetation Richness and Diversity Relationships in the Tropics. Veg. Hist. Archaeobotany 2018, 27, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Schwörer, C.; Colombaroli, D.; Kaltenrieder, P.; Rey, F.; Tinner, W. Early Human Impact (5000–3000 BC) Affects Mountain Forest Dynamics in the Alps. J. Ecol. 2015, 103, 281–295. [Google Scholar] [CrossRef]
- Jantz, N.; Behling, H. A Holocene Environmental Record Reflecting Vegetation, Climate, and Fire Variability at the Páramo of Quimsacocha, Southwestern Ecuadorian Andes. Veg. Hist. Archaeobotany 2012, 21, 169–185. [Google Scholar] [CrossRef] [Green Version]
- McGlinn, D.J.; Xiao, X.; May, F.; Gotelli, N.J.; Engel, T.; Blowes, S.A.; Knight, T.M.; Purschke, O.; Chase, J.M.; McGill, B.J. Measurement of Biodiversity (MoB): A Method to Separate the Scale-Dependent Effects of Species Abundance Distribution, Density, and Aggregation on Diversity Change. Methods Ecol. Evol. 2019, 10, 258–269. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a Fire Proxy. In Tracking Environmental Change Using Lake Sediments; Smol, J.P., Birks, H.J.B., Last, W.M., Bradley, R.S., Alverson, K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; Volume 3, pp. 75–97. ISBN 1-4020-0681-0. [Google Scholar]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next Generation of Scientific Image Data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Higuera, P.E.; Brubaker, L.B.; Anderson, P.M.; Hu, F.S.; Brown, T.A. Vegetation Mediated the Impacts of Postglacial Climate Change on Fire Regimes in the South-Central Brooks Range, Alaska. Ecol. Monogr. 2009, 79, 201–219. [Google Scholar] [CrossRef]
- Hsu, L.-T. The Study of the Vegetation Changes in Yangmingshan National Park; Yangmingshan National Park Headquarters: Taipei, Taiwan, 2008; p. 121. [Google Scholar]
- Woodward, C.; Haines, H.A. Unprecedented Long-Distance Transport of Macroscopic Charcoal from a Large, Intense Forest Fire in Eastern Australia: Implications for Fire History Reconstruction. Holocene 2020, 30, 947–952. [Google Scholar] [CrossRef]
- Conedera, M.; Tinner, W.; Neff, C.; Meurer, M.; Dickens, A.F.; Krebs, P. Reconstructing Past Fire Regimes: Methods, Applications, and Relevance to Fire Management and Conservation. Quat. Sci. Rev. 2009, 28, 555–576. [Google Scholar] [CrossRef]
- Bunting, M.J.; Gaillard, M.-J.; Sugita, S.; Middleton, R.; Broström, A. Vegetation Structure and Pollen Source Area. The Holocene 2004, 14, 651–660. [Google Scholar] [CrossRef]
- Conedera, M.; Tinner, W.; Crameri, S.; Torriani, D.; Herold, A. Taxon-Related Pollen Source Areas for Lake Basins in the Southern Alps: An Empirical Approach. Veg. Hist. Archaeobotany 2006, 15, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Liew, P.-M.; Wu, M.-H.; Lee, C.-Y.; Chang, C.-L.; Lee, T.-Q. Recent 4000 Years of Climatic Trends Based on Pollen Records from Lakes and a Bog in Taiwan. Quat. Int. 2014, 349, 105–112. [Google Scholar] [CrossRef]
- Luo, C.; Chen, M.; Xiang, R.; Liu, J.; Zhang, L.; Lu, J.; Yang, M. Modern Pollen Distribution in Marine Sediments from the Northern Part of the South China Sea. Mar. Micropaleontol. 2014, 108, 41–56. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Liew, P.-M.; Lee, T.-Q. Pollen Records from Southern Taiwan: Implications for East Asian Summer Monsoon Variation during the Holocene. Holocene 2010, 20, 81–89. [Google Scholar] [CrossRef]
- Wang, L.-C.; Wu, J.-T.; Lee, T.-Q.; Lee, P.-F.; Chen, S.-H. Climate Changes Inferred from Integrated Multi-Site Pollen Data in Northern Taiwan. J. Asian Earth Sci. 2011, 40, 1164–1170. [Google Scholar] [CrossRef]
- Wang, L.-C.; Behling, H.; Kao, S.-J.; Li, H.-C.; Selvaraj, K.; Hsieh, M.-L.; Chang, Y.-P. Late Holocene Environment of Subalpine Northeastern Taiwan from Pollen and Diatom Analysis of Lake Sediments. J. Asian Earth Sci. 2015, 114, 447–456. [Google Scholar] [CrossRef]
- Wang, L.-C.; Tang, Z.-W.; Chen, H.-F.; Li, H.-C.; Shiau, L.-J.; Huang, J.-J.S.; Wei, K.-Y.; Chuang, C.-K.; Chou, Y.-M. Late Holocene Vegetation, Climate, and Natural Disturbance Records from an Alpine Pond in Central Taiwan. Quat. Int. 2019, 528, 63–72. [Google Scholar] [CrossRef]
- Huang, Z.; Ma, C.; Chyi, S.-J.; Tang, L.; Zhao, L. Paleofire, Vegetation, and Climate Reconstructions of the Middle to Late Holocene From Lacustrine Sediments of the Toushe Basin, Taiwan. Geophys. Res. Lett. 2020, 47, e2020GL090401. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, Y.; Edwards, R.L.; Cheng, H.; Liu, D.; Kong, X. A High-Resolved Record of the Asian Summer Monsoon from Dongge Cave, China for the Past 1200 Years. Quat. Sci. Rev. 2015, 122, 250–257. [Google Scholar] [CrossRef]
- Zhang, Z.; Yao, Q.; Liu, K.; Li, L.; Yin, R.; Wang, G.; Sun, J. Historical Flooding Regime along the Amur River and Its Links to East Asia Summer Monsoon Circulation. Geomorphology 2021, 388, 107782. [Google Scholar] [CrossRef]
- Manville, V.; Hodgson, K.A.; Nairn, I.A. A Review of Break-out Floods from Volcanogenic Lakes in New Zealand. N. Z. J. Geol. Geophys. 2007, 50, 131–150. [Google Scholar] [CrossRef]
- Wang, L.-C.; Behling, H.; Chen, Y.-M.; Huang, M.-S.; Chen, C.-T.A.; Lou, J.-Y.; Chang, Y.-P.; Li, H.-C. Holocene Monsoonal Climate Changes Tracked by Multiproxy Approach from a Lacustrine Sediment Core of the Subalpine Retreat Lake in Taiwan. Quat. Int. 2014, 333, 69–76. [Google Scholar] [CrossRef]
- Wang, L.-C.; Behling, H.; Lee, T.-Q.; Li, H.-C.; Huh, C.-A.; Shiau, L.-J.; Chang, Y.-P. Late Holocene Environmental Reconstructions and Their Implications on Flood Events, Typhoon, and Agricultural Activities in NE Taiwan. Clim. Past 2014, 10, 1857–1869. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Mao, L.; Lü, X.; Ma, Q.; Zhou, Z.; Yang, C.; Kong, Z.; Batten, D.J. Palaeoecological and Palaeoenvironmental Significance of Some Important Spores and Micro-Algae in Quaternary Deposits. Chin. Sci. Bull. 2013, 58, 3125–3139. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-C.; Chou, Y.-M.; Chen, H.-F.; Chang, Y.-P.; Chiang, H.-W.; Yang, T.-N.; Shiau, L.-J.; Chen, Y.-G. Paleolimnological Evidence for Lacustrine Environmental Evolution and Paleo-Typhoon Records during the Late Holocene in Eastern Taiwan. J. Paleolimnol. 2021. [Google Scholar] [CrossRef]
- Park, J.; Han, J.; Jin, Q.; Bahk, J.; Yi, S. The Link between ENSO-like Forcing and Hydroclimate Variability of Coastal East Asia during the Last Millennium. Sci. Rep. 2017, 7, 8166. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.M.; Conroy, J.L.; Collins, A.; Hlohowskyj, S.R.; Overpeck, J.T.; Riedinger-Whitmore, M.; Cole, J.E.; Bush, M.B.; Whitney, H.; Corley, T.L.; et al. Tropical Pacific Climate Variability over the Last 6000 Years as Recorded in Bainbridge Crater Lake, Galápagos. Paleoceanography 2017, 32, 903–922. [Google Scholar] [CrossRef]
- Cheng, H.; Edwards, R.L.; Sinha, A.; Spötl, C.; Yi, L.; Chen, S.; Kelly, M.; Kathayat, G.; Wang, X.; Li, X.; et al. The Asian Monsoon over the Past 640,000 Years and Ice Age Terminations. Nature 2016, 534, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-L.; Wang, W.-H.; Wen, S. Dense Seismic Arrays Deny a Massive Magma Chamber beneath the Taipei Metropolis, Taiwan. Sci. Rep. 2021, 11, 1083. [Google Scholar] [CrossRef] [PubMed]
- de Porras, M.E.; Maldonado, A.; Abarzúa, A.M.; Cárdenas, M.L.; Francois, J.P.; Martel-Cea, A.; Stern, C.R.; Méndez, C.; Reyes, O. Postglacial Vegetation, Fire and Climate Dynamics at Central Chilean Patagonia (Lake Shaman, 44° S). Quat. Sci. Rev. 2012, 50, 71–85. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Chang, C.-L.; Liew, P.-M.; Lee, T.-Q.; Song, S.-R. Climate Change, Vegetation History, and Agricultural Activity of Lake Li-Yu Tan, Central Taiwan, during the Last 2.6 Ka BP. Quat. Int. 2014, 325, 105–110. [Google Scholar] [CrossRef]
- Wenske, D.; Böse, M.; Frechen, M.; Lüthgens, C. Late Holocene Mobilisation of Loess-like Sediments in Hohuan Shan, High Mountains of Taiwan. Quat. Int. 2011, 234, 174–181. [Google Scholar] [CrossRef]
- Fang, K.; Yao, Q.; Guo, Z.; Zheng, B.; Du, J.; Qi, F.; Yan, P.; Li, J.; Ou, T.; Liu, J.; et al. ENSO Modulates Wildfire Activity in China. Nat. Commun. 2021, 12, 1764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cui, B.; Lan, Y.; Han, Z.; Wang, T.; Guo, A. Four Terrestrialization Characteristics of Baiyangdian Lake, China. Procedia Environ. Sci. 2012, 13, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Wang, L.; Zhou, W.; Chen, W.; Wu, R.; Liu, L.; Nath, D.; Leung, M.Y.-T. Revisiting the Northern Mode of East Asian Winter Monsoon Variation and Its Response to Global Warming. J. Clim. 2018, 31, 9001–9014. [Google Scholar] [CrossRef]
- Taylor, W.C.; Hickey, R.J. Habitat, Evolution, and Speciation in Isoetes. Ann. MO Bot. Gard. 1992, 79, 613–622. [Google Scholar] [CrossRef]
- Kim, C.; Shin, H.; Chang, Y.-T.; Choi, H.-K. Speciation Pathway of Isoëtes (Isoëtaceae) in East Asia Inferred from Molecular Phylogenetic Relationships. Am. J. Bot. 2010, 97, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.-Y.; Wang, Q.-F. Current Status and Conservation Strategies for Isoetes in China: A Case Study for the Conservation of Threatened Aquatic Plants. Oryx 2005, 39, 335–338. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab Code | Sample ID | Depth (cm) | Type of Sample | pMC (%) | 14C Age (BP) | Calibrated 14C Age (cal BP) |
|---|---|---|---|---|---|---|
| NTUAMS-4088 | DL 18 | 18 | peat | 91.41 ± 0.90 | 722 ± 7 | 669 ± 6 |
| NTUAMS-4472-1 | DL 40 | 40 | peat | 84.13 ± 1.13 | 1389 ± 19 | 1299 ± 14 |
| NTUAMS-4473-2 | DL 60 | 60 | peat | 78.56 ± 1.09 | 1938 ± 27 | 1857 ± 78 |
| NTUAMS-4089 | DL 82 | 82 | peat | 68.53 ± 0.88 | 3035 ± 39 | 3252 ± 107 |
| NTUAMS-4090 | DL 130 | 130 | sediment | 65.82 ± 0.70 | 3359 ± 36 | 3565 ± 82 |
| NTUAMS-4474-1 | DL 160 | 160 | sediment | 62.17 ± 0.87 | 3818 ± 53 | 4247 ± 162 |
| NTUAMS-4091 | DL 176 | 176 | sediment | 61.17 ± 0.78 | 3948 ± 50 | 4382 ± 141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-C. Using Paleoecological Data to Inform the Conservation Strategy for Floristic Diversity and Isoetes taiwanensis in Northern Taiwan. Diversity 2021, 13, 395. https://doi.org/10.3390/d13080395
Wang L-C. Using Paleoecological Data to Inform the Conservation Strategy for Floristic Diversity and Isoetes taiwanensis in Northern Taiwan. Diversity. 2021; 13(8):395. https://doi.org/10.3390/d13080395
Chicago/Turabian StyleWang, Liang-Chi. 2021. "Using Paleoecological Data to Inform the Conservation Strategy for Floristic Diversity and Isoetes taiwanensis in Northern Taiwan" Diversity 13, no. 8: 395. https://doi.org/10.3390/d13080395