
Environmental Factors Affecting the Spatiotemporal Distribution of Copepods in a Small Mesotidal Inlet and Estuary


 and
and
Abstract
:1. Introduction
2. Materials and Methods
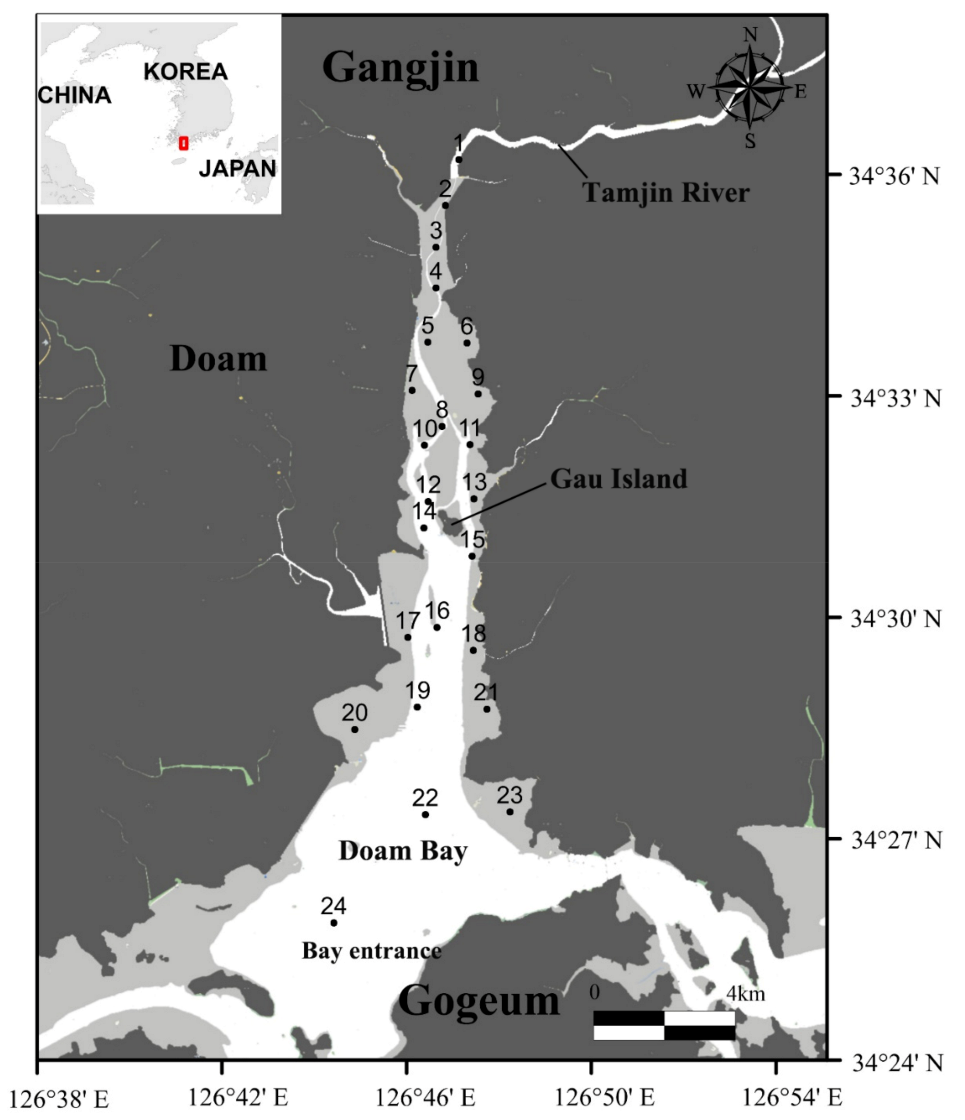
2.1. Study Area
2.2. Environmental Factors
2.3. Mesozooplankton Sampling and Identification
2.4. Data Analysis
3. Results
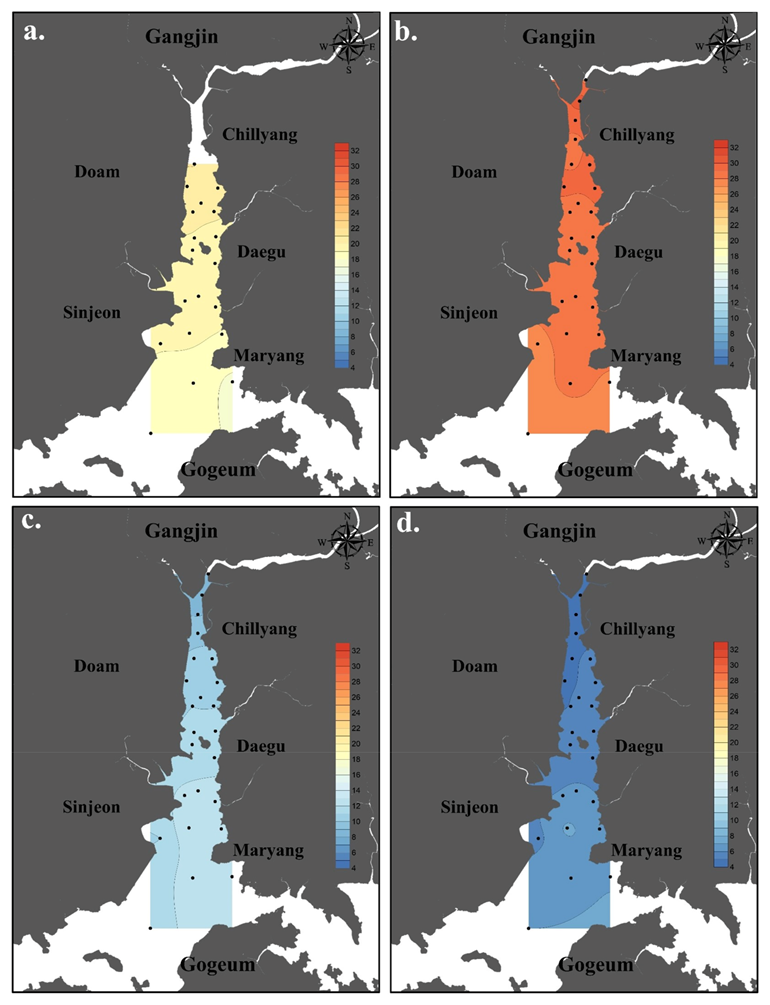
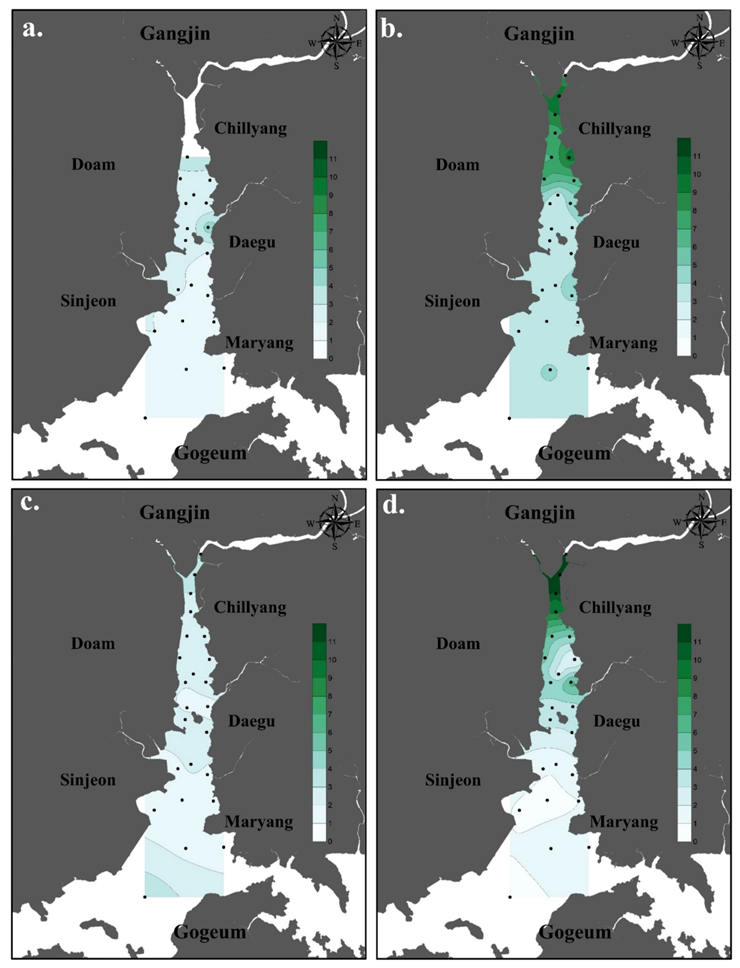
3.1. Environmental Factors
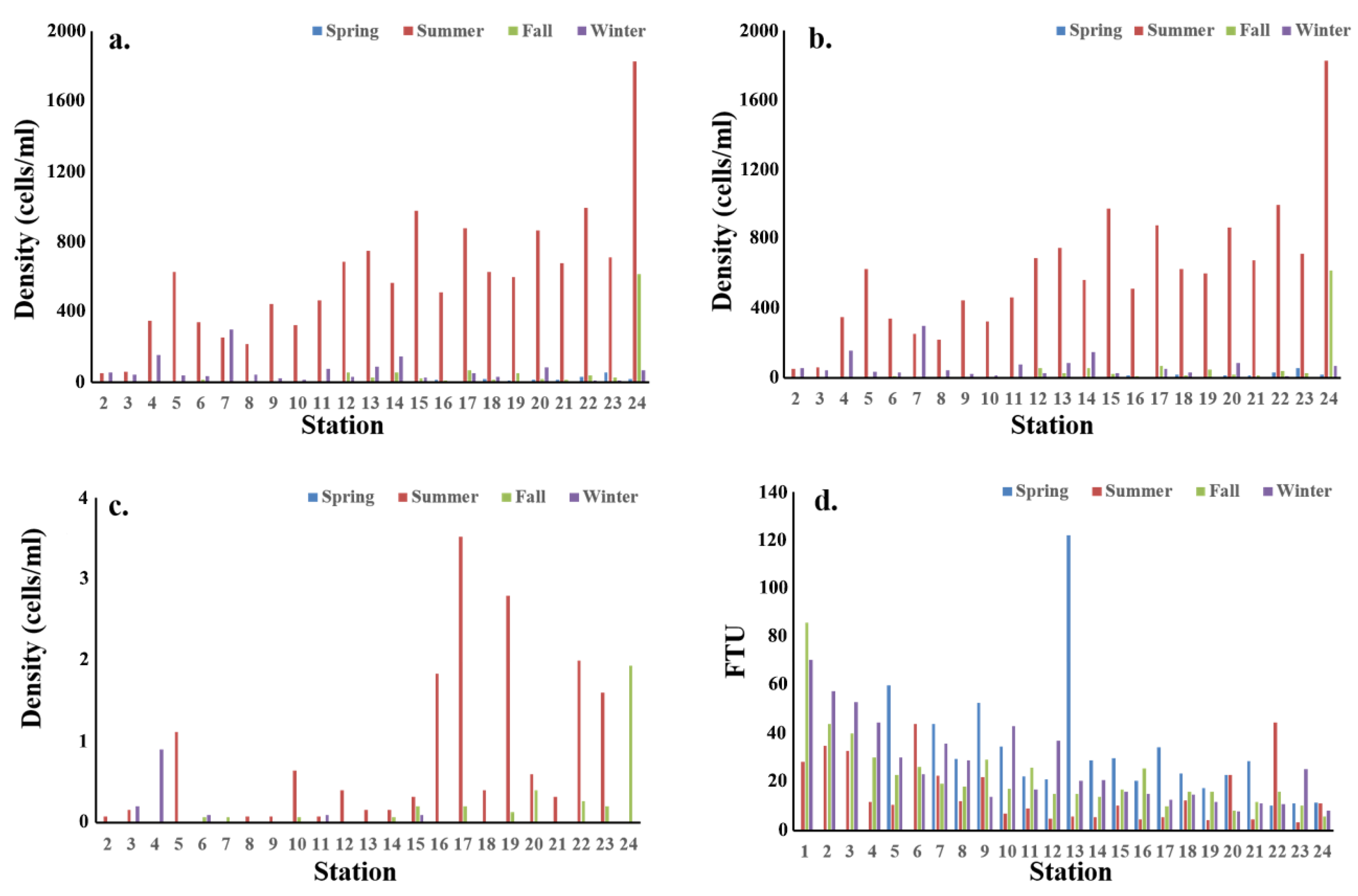
3.2. The Abundance of Mesozooplankton and Copepods
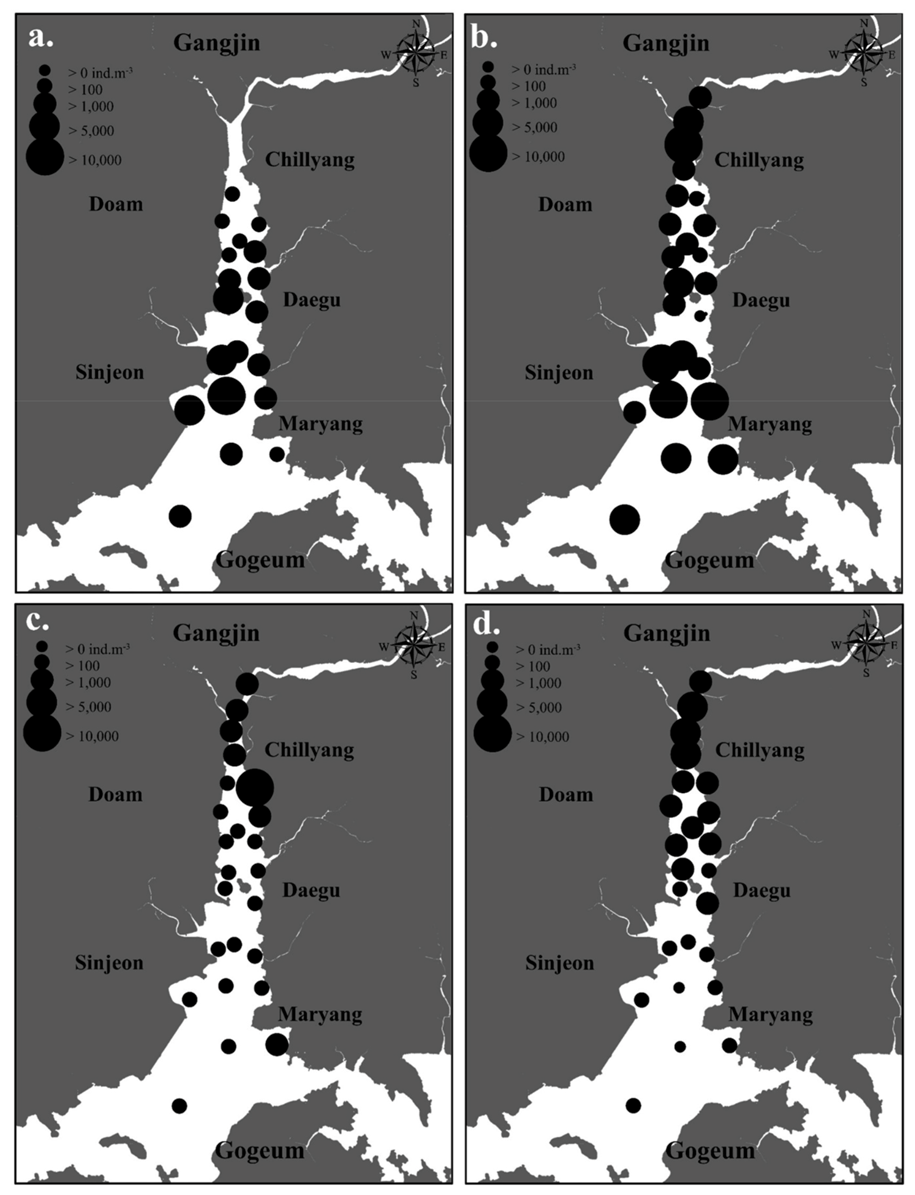
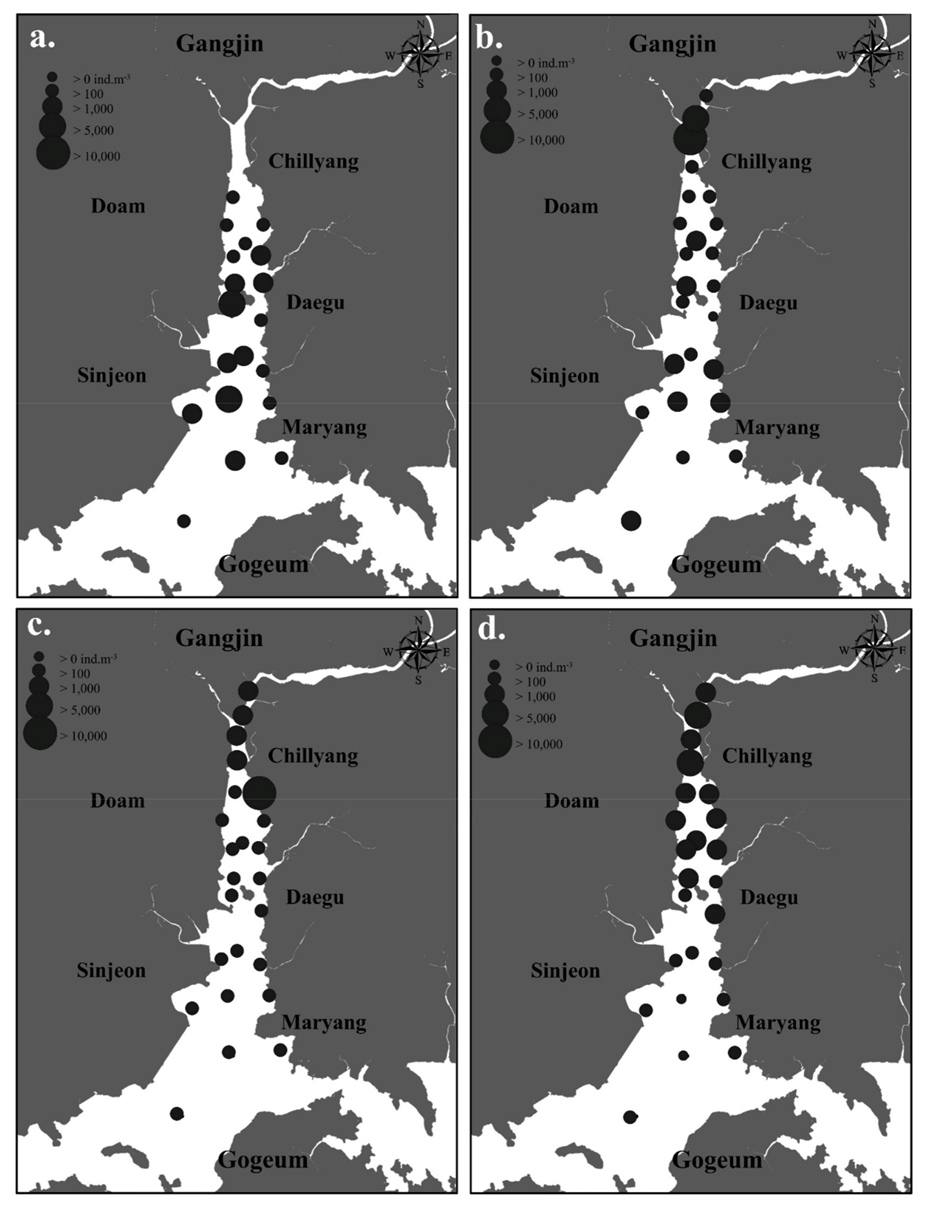
3.3. The Spatiotemporal Distribution of Copepods
3.4. Seasonal Copepod Communities by Cluster Analysis
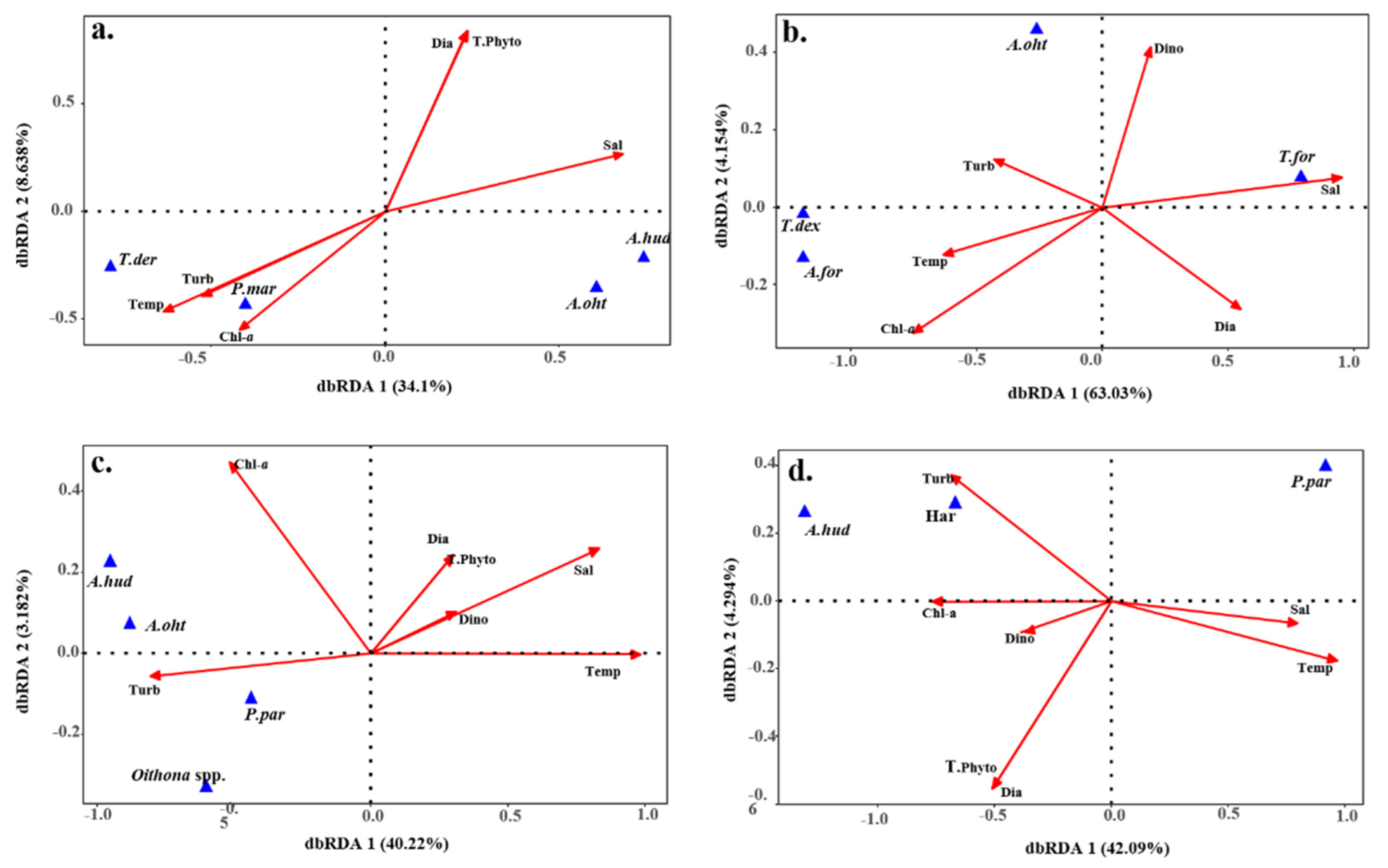
3.5. Seasonal Correlation between Environmental Factors and Major Copepods
4. Discussion
4.1. Environmental Characteristics of Doam Bay
4.2. Geographical Distribution Characteristics of Brackish Copepods in Doam Bay
4.3. Environmental Characteristics and Spatiotemporal Distribution of Major Copepods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pritchard, D.W. What is an estuary: Physical view point. AAAS 1967, 83, 3–5. [Google Scholar]
- Day, J.W.; Kemp, W.M.; Yáñez-Arancibia, A.; Crump, B.C. Estuarine Ecology, 2nd ed.; Wiley-Blackwell: New York, NY, USA, 2012; pp. 1–558. [Google Scholar]
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem: Ecology, Threats and Management; Oxford University Press: Oxford, UK, 2004; pp. 1–209. [Google Scholar]
- Rodríguez, G. Some Aspects of the Ecology of Tropical Estuaries. In Tropical Ecological Systems; Golley, F.B., Medina, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 313–332. [Google Scholar]
- Grindley, J.R. Estuarine plankton. In Estuarine Ecology with Particular Reference to Southern Africa; Day, J.H., Ed.; Balkema: Cape Town, South Africa, 1981; pp. 117–146. [Google Scholar]
- Grindley, J.R. The role of zooplankton in the St. Lucia estuary system. In St. Lucia Research Review; Taylor, R.H., Ed.; Natal Parks Board Symposium: Pietermaritzburg, South Africa, 1982; pp. 88–107. [Google Scholar]
- Lam-Hoai, T.; Guiral, D.; Rougier, C. Seasonal change of community structure and size spectra of zooplankton in the Kaw River estuary (French Guiana). Estuar. Coast. Shelf Sci. 2006, 68, 47–61. [Google Scholar] [CrossRef]
- Schmitt, F.G.; Devreker, D.; Dur, G.; Souissi, S. Direct evidence of tidally oriented behavior of the copepod Eurytemora affinis in the Seine estuary. Ecol. Res. 2011, 26, 773–780. [Google Scholar] [CrossRef]
- Davies, J.L. A morphogenic approach to world shorelines. Z. Geomorphol. 1964, 8, 127–142. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Wooldridge, T.H. Ecological responses to reductions in freshwater supply and quality in South Africa’s estuaries: Lessons for management and conservation. J. Coast. Conserv. 1996, 2, 115–130. [Google Scholar] [CrossRef]
- Domingues, R.B.; Galvao, H. Phytoplankton and environmental variability in a dam regulated temperate estuary. Hydrobiologia 2007, 586, 117–134. [Google Scholar] [CrossRef]
- Morais, P.; Chícharo, M.A.; Chícharo, L. Changes in a temperate estuary during the filling of the biggest European dam. Sci. Total Environ. 2009, 407, 2245–2259. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Han, R.; Wang, B.; Tang, X. Zooplankton community in Yangtze River Estuary and adjacent sea areas after the impoundment of the Three Gorges Reservoir. Ann. Limnol. Int. J. Limnol. 2016, 52, 273–284. [Google Scholar] [CrossRef]
- Yen, N.T.M.; Vanreusel, A.; Lins, L.; Thai, T.T.; Nara, B.T.; Quang, N.X. The effect of a dam construction on subtidal nematode communities in the Ba Lai Estuary, Vietnam. Diversity 2020, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Suh, H.L. Salinity and distribution of zooplankton in the estuarine system of Mankyong River and Dongjin River. J. Oceanol. Soc. Korea 1991, 26, 181–192. [Google Scholar]
- Park, C.; Lee, P.G.; Yang, S.R. Variation of zooplankton distribution in the Seomjin River estuary with respect to season and salinity gradients. Sea 2002, 7, 51–59. [Google Scholar]
- Youn, S.H.; Choi, J.K. Distribution pattern of zooplankton in the Han River estuary with respect to tidal cycle. Ocean Sci. J. 2008, 43, 135–146. [Google Scholar] [CrossRef]
- Youn, S.H.; Oh, G.S.; Chung, M.H. Zooplankton community structure and copepod production in the Seomjin River Estuary. J. Korean Soc. Mar. Environ. Saf. 2010, 16, 369–379. [Google Scholar]
- Park, E.O.; Rahman, M.S.; Seo, M.H.; Kim, J.J.; Soh, H.Y. Distribution patterns of calanoid copepods along the Seomjin River estuary in southern Korea during summer. Korean J. Environ. Biol. 2013, 31, 165–171. [Google Scholar] [CrossRef]
- Park, E.O.; Suh, H.L.; Soh, H.Y. Spatio-temporal distribution of Acartia (Copepoda: Calanoida) species along a salinity gradient in the Seomjin River estuary, South Korea. J. Nat. Hist. 2015, 49, 2799–2812. [Google Scholar] [CrossRef]
- Hong, J.T.; Na, B.S.; Kim, J.Y.; Koh, Y.K.; Youn, S.T.; Shin, S.E.; Oh, K.H. Sedimentary geochemical characteristics and environmental impact of sediments in Tamjin River and Doam Bay. J. Environ. Impact Assess. 2007, 16, 393–405. [Google Scholar]
- Chihara, M.; Murano, M. An Illustrated Guide to Marine Plankton in Japan; Tokai University Press: Tokyo, Japan, 1997; pp. 1–260. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; Plymouth Marine Laboratory: Plymouth, UK, 2001; pp. 1–262. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics. R Package Version 2016, 2, 1–189. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2019, 9, 1–287. [Google Scholar]
- Kwak, M.T.; Cho, Y.K. Seasonal Variation in Residence Times of Two Neighboring Bays with Contrasting Topography. Estuaries Coast. 2019, 43, 1–13. [Google Scholar] [CrossRef]
- Jordan, T.E.; Correll, D.L.; Miklas, J.J.; Weller, D.E. Long-term trends in estuarine nutrients and chlorophyll, and short-term effects of variation in watershed discharge. Mar. Ecol. Prog. Ser. 1991, 75, 121–132. [Google Scholar] [CrossRef]
- O’Higgins, T.G.; Wilson, J.G. Impact of the river Liffey discharge on nutrient and chlorophyll concentrations in the Liffey estuary and Dublin Bay (Irish Sea). Estuar. Coast. Shelf Sci. 2005, 64, 323–334. [Google Scholar] [CrossRef]
- National Climate Data Center of Korea Meteorological Administration. Available online: https://data.kma.go.kr/data/grnd/selectAsosRltmList.do?pgmNo=36 (accessed on 14 July 2021).
- Delgado, M.; de Jonge, V.N.; Peletier, H. Experiments on resuspension of natural microphytobenthos populations. Mar. Biol. 1991, 108, 321–328. [Google Scholar] [CrossRef]
- Kim, S. Pre-monsoon dynamics of zooplankton community in the estuary of the Tamjin River, Korea. Korean J. Ecol. Environ. 2015, 33, 19–26. [Google Scholar]
- Soh, H.Y.; Kwon, S.W.; Lee, W.; Yoon, Y.H. A new Pseudodiaptomus (Copepoda, Calanoida) from Korea supported by molecular data. Zootaxa 2012, 3368, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Im, D.H.; Suh, H.L. Resource utilization of three copepods along salinity gradient in the Seomjin River estuary, revealed by stable isotope analysis. Ocean Sci. J. 2021, 56, 106–116. [Google Scholar] [CrossRef]
- Kim, S.W.; Han, M.S.; Lee, K.; Choi, Y.K.; Yoo, K.I. Zooplankton and phytoplankton dynamics with the construction of river mouth dam in Kum River estuary, Korea. Korean J. Ecol. Environ. 2002, 35, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.K.; Jeong, J.H.; Nam, E.J.; Jeong, K.M.; Lee, S.W.; Myung, C.S. Zooplankton community and distribution in relation to water quality in the Saemangeum area, Korea: Change in Zooplankton community by the construction of sea Dyke. Ocean Polar Res. 2006, 28, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; Kang, H.K.; Noh, J.H. Temporal and spatial variation of zooplankton community structure post construction of Saemangeum dyke. Ocean Polar Res. 2009, 31, 327–338. [Google Scholar] [CrossRef]
- Wooldridge, T.H. Estuarine zooplankton community structure and dynamics. In Estuaries of South Africa; Allanson, B.R., Baird, D., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 141–166. [Google Scholar]
- Rose, T.H.; Tweedley, J.R.; Warwick, R.M.; Potter, I.C. Influences of microtidal regime and eutrophication on estuarine zooplankton. Estuar. Coast. Shelf Sci. 2020, 238, 106689. [Google Scholar] [CrossRef]
- Soh, H.Y.; Suh, H.L.; Ohtsuka, S.; Yoon, Y.H.; Choi, S.D. Taxonomic studies on brackish copepods in Korean waters—II. Ontogeny and phylogeny of appendages in copepodid stages of Tortanus derjugini Smirnov, 1935 (Copepoda, Calanoida). J. Plankton Res. 2001, 23, 1157–1169. [Google Scholar] [CrossRef]
- Choi, S.Y.; Seo, M.H.; Soh, H.Y. Short-term variation in abundance of four Acartia species (Copepoda, Calanoida) in a eutrophic bay. PeerJ 2021, 9, e10835. [Google Scholar] [CrossRef]
- Jacobs, J. Animal behavior and water movement as co-determinants of plankton distribution in a tidal system. Sarsia 1968, 34, 355–370. [Google Scholar] [CrossRef]
- Wooldridge, T.H.; Erasmus, T. Utilization of tidal currents by estuarine zooplankton. Estuar. Coast. Mar. Sci. 1980, 11, 107–114. [Google Scholar] [CrossRef]
- Ueda, H.; Terao, A.; Tanaka, M.; Hibino, M.; Islam, M.S. How can river-estuarine planktonic copepods survive river floods? Ecol. Res. 2004, 19, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Kimmerer, W.J.; Gross, E.S.; MacWilliams, M.L. Tidal migration and retention of estuarine zooplankton investigated using a particle-tracking model. Limnol. Oceanogr. 2014, 59, 901–916. [Google Scholar] [CrossRef]
- Sullivan, B.K.; McManus, L.T. Factors controlling seasonal succession of the copepods Acartia hudsonica and A. tonsa in Narragansett Bay, Rhode Island: Temperature and resting egg production. Mar. Ecol. Prog. Ser. 1986, 28, 121–128. [Google Scholar] [CrossRef]
- Ueda, H. Temporal and spatial distribution of the two closely related Acartia species A. omorii and A. hudsonica (Copepoda, Calanoida) in a small inlet water of Japan. Estuar. Coast. Shelf Sci. 1987, 24, 691–700. [Google Scholar] [CrossRef]
- Lawrence, D.; Valiela, I.; Tomasky, G. Estuarine calanoid copepod abundance in relation to season, salinity, and land-derived nitrogen loading, Waquoit Bay, MA. Estuar. Coast. Shelf Sci. 2004, 2004. 61, 547–557. [Google Scholar] [CrossRef]
- Lee, E.H.; Seo, M.H.; Yoon, Y.H.; Choi, S.D.; Soh, H.Y. Environmental factors affecting zooplankton community in Gwangyang Bay. Korean J. Environ. Biol. 2017, 2017. 35, 631–639. [Google Scholar] [CrossRef]
- Sun, X.H.; Sun, S.; Li, C.L.; Zhang, G.T. Seasonal and spatial variation in abundance and egg production of Paracalanus parvus (Copepoda: Calanoida) in/out Jiaozhou Bay, China. Estuar. Coast. Shelf Sci. 2008, 79, 637–643. [Google Scholar] [CrossRef]
- Islam, M.S.; Ueda, H.; Tanaka, M. Spatial and seasonal variations in copepod communities related to turbidity maximum along the Chikugo estuarine gradient in the upper Ariake Bay, Japan. Estuar. Coast. Shelf Sci. 2006, 68, 113–126. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | 2018 | 2019 | ||
|---|---|---|---|---|
| Spring | Summer | Fall | Winter | |
| Trachylinae | ||||
| Unid. Trachymedusae | 0.30 | 1.34 | 5.96 | |
| Cladocera | ||||
| Pleopis polyphemoides | 0.02 | |||
| Calanoida (Copepoda) | ||||
| Acartia erythraea | 0.40 | |||
| Acartia hongi | 0.14 | 0.15 | 0.02 | |
| Acartia hudsonica | 38.97 | 43.10 | 75.53 | |
| Acartia ohtsukai | 14.08 | 6.75 | 10.84 | |
| Acartia forticrusa | 14.89 | |||
| Bestiolina coreana | 0.14 | |||
| Calanopia thompsoni | 0.03 | |||
| Calanus sinicus | 0.02 | |||
| Centropages abdominalis | 0.05 | |||
| Centropages tenuiremis | 0.01 | 0.04 | ||
| Eurytemora pacifica | 0.07 | 0.49 | ||
| Labidocera pavo | 0.00 * | |||
| Labidocera rotunda | 0.04 | |||
| Paracalanus parvus s. l. | 1.00 | 2.63 | 23.68 | 0.68 |
| Parvocalanus crassirostris | 0.06 | |||
| Pseudodiaptomus japonicus | 0.03 | 0.04 | 0.03 | |
| Pseudodiaptomus marinus | 16.55 | 0.07 | 0.11 | 0.87 |
| Tortanus derjugini | 1.56 | |||
| Tortanus dextrilobatus | 0.24 | 0.53 | 0.85 | |
| Tortanus forcipatus | 2.03 | 0.04 | ||
| Cyclopoida (Copepoda) | ||||
| Ditrichocorycaeus affinis | 0.02 | 0.07 | 0.26 | 0.12 |
| Oithona spp. | 0.04 | 2.12 | 8.00 | 0.04 |
| Harpacticoida (Copepoda) | ||||
| Unid. Harpacticoida | 0.29 | 0.18 | 0.87 | 14.71 |
| Multicrustacea | ||||
| Unid. Amphipods | 0.07 | 0.03 | ||
| Unid. Mysids | 0.03 | |||
| Unid. Isopods | 0.28 | 0.02 | ||
| Chaetognatha | ||||
| Aidanosagitta crassa | 0.04 | 0.41 | 0.09 | |
| Larvae | ||||
| Cirripedia larvae | 17.83 | 35.93 | 3.47 | 0.54 |
| Decapoda larvae | 3.19 | 2.83 | 0.08 | |
| Bivalvia larvae | 2.50 | 3.13 | 0.05 | |
| Gastropoda larvae | 1.18 | 6.39 | 0.02 | 0.04 |
| ophiopluteus larvae | 8.70 | 0.07 | ||
| Polychaeta larvae | 0.49 | 0.47 | 0.41 | 0.81 |
| Appendicularia | ||||
| Oikopleura dioica | 11.24 | 7.44 | ||
| Total abundance (ind./m3) | 2793 ± 2924 | 5578 ± 6469 | 1122 ± 2324 | 2091 ± 2140 |
| Copepoda(ind./m3) | 2039 ± 2350 | 1668 ± 2678 | 988 ± 2274 | 1934 ± 2032 |
| Season | Species | AF | AH | AO | PP | PM | TDER | TDEX | TF | OS | Harp | Temp | Sal | Chl-a | Phyto | Dia | Dino | Turb |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spring | Acartia hudsonica (AH) | - | 1 | 0.848 ** | - | −0.090 | −0.383 | - | - | - | - | −0.233 | 0.282 | −0.243 | −0.021 | −0.022 | - | −0.241 |
| Acartia ohtsukai (AO) | - | 0.848 ** | 1 | - | −0.119 | −0.335 | − | - | - | - | −0.220 | 0.265 | −0.149 | −0.114 | −0.115 | - | −0.135 | |
| Pseudodiaptomus marinus (PM) | - | −0.090 | −0.119 | - | 1 | 0.479 * | - | - | - | - | 0.160 | −0.060 | 0.132 | −0.269 | −0.268 | - | 0.048 | |
| Tortanus derjugini (TDER) | - | −0.383 | −0.335 | - | 0.479 * | 1 | - | - | - | - | 0.582 ** | −0.513 * | 0.544 * | −0.423 | −0.420 | - | 0.487 * | |
| Summer | Acartia ohtsukai (AO) | 0.120 | - | 1 | - | - | - | 0.116 | 0.194 | - | - | −0.041 | −0.015 | −0.121 | −0.104 | −0.105 | 0.426 * | −0.044 |
| Acartia forticrusa (AF) | 1 | - | 0.120 | - | - | - | 0.979 ** | −0.211 | - | - | 0.578 ** | −0.851 ** | 0.647 ** | −0.462 * | −0.462 * | −0.195 | 0.455 * | |
| Tortanus dextrilobatus (TDEX) | 0.979 ** | - | 0.116 | - | - | - | 1 | −0.169 | - | - | 0.493 * | −0.751 ** | 0.599 ** | −0.442 * | −0.442 * | −0.186 | 0.432 * | |
| Tortanus forcipatus (TF) | −0.211 | − | 0.194 | - | - | - | −0.169 | 1 | - | - | −0.118 | 0.242 | −0.213 | 0.089 | 0.090 | −0.041 | −0.195 | |
| Fall | Acartia hudsonica (AH) | - | 1 | 0.989 ** | 0.948 ** | - | - | - | - | 0.984 ** | - | −0.328 | −0.264 | 0.169 | −0.105 | −0.105 | −0.103 | 0.329 |
| Acartia ohtsukai (AO) | - | 0.989 ** | 1 | 0.955 ** | - | - | - | - | 0.975 ** | - | −0.225 | −0.143 | 0.112 | −0.075 | −0.075 | −0.067 | 0.224 | |
| Paracalanus parvus s. l. (PP) | - | 0.948 ** | 0.955 ** | 1 | - | - | - | - | 0.971 ** | - | −0.237 | −0.160 | 0.095 | −0.072 | −0.072 | −0.047 | 0.187 | |
| Oithona spp. (OS) | - | 0.984 ** | 0.975 ** | 0.971 ** | - | - | - | - | 1 | - | −0.291 | −0.220 | 0.122 | −0.114 | −0.114 | −0.104 | 0.274 | |
| Winter | Acartia hudsonica (AH) | - | 1 | - | −0.125 | - | - | - | - | - | 0.313 | −0.580 ** | −0.746 ** | 0.760 ** | 0.216 | 0.214 | 0.663 ** | 0.656 ** |
| Paracalanus parvus s. l. (PP) | - | −0.125 | - | 1 | - | - | - | - | - | −0.070 | 0.316 | 0.359 | −0.373 | −0.251 | −0.250 | −0.224 | −0.281 | |
| Harpacticoida (Harp) | - | 0.313 | - | −0.070 | - | - | - | - | - | 1 | −0.366 | −0.423 * | 0.422 * | 0.371 | 0.371 | 0.006 | 0.442 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, M.-H.; Kim, H.-J.; Lee, S.-J.; Kim, S.-Y.; Yoon, Y.-H.; Han, K.-H.; Choi, S.-D.; Kwak, M.-T.; Jeong, M.-K.; Soh, H.-Y. Environmental Factors Affecting the Spatiotemporal Distribution of Copepods in a Small Mesotidal Inlet and Estuary. Diversity 2021, 13, 389. https://doi.org/10.3390/d13080389
Seo M-H, Kim H-J, Lee S-J, Kim S-Y, Yoon Y-H, Han K-H, Choi S-D, Kwak M-T, Jeong M-K, Soh H-Y. Environmental Factors Affecting the Spatiotemporal Distribution of Copepods in a Small Mesotidal Inlet and Estuary. Diversity. 2021; 13(8):389. https://doi.org/10.3390/d13080389
Chicago/Turabian StyleSeo, Min-Ho, Hyeon-Jung Kim, Seok-Ju Lee, So-Yeon Kim, Yang-Ho Yoon, Kyeong-Ho Han, Sang-Duck Choi, Myeong-Taek Kwak, Man-Ki Jeong, and Ho-Young Soh. 2021. "Environmental Factors Affecting the Spatiotemporal Distribution of Copepods in a Small Mesotidal Inlet and Estuary" Diversity 13, no. 8: 389. https://doi.org/10.3390/d13080389