
Reproduction of the Androgenetic Population of the Asian Corbicula Clam (Bivalvia: Cyrenidae) in the Northern Dvina River Basin, Russia

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
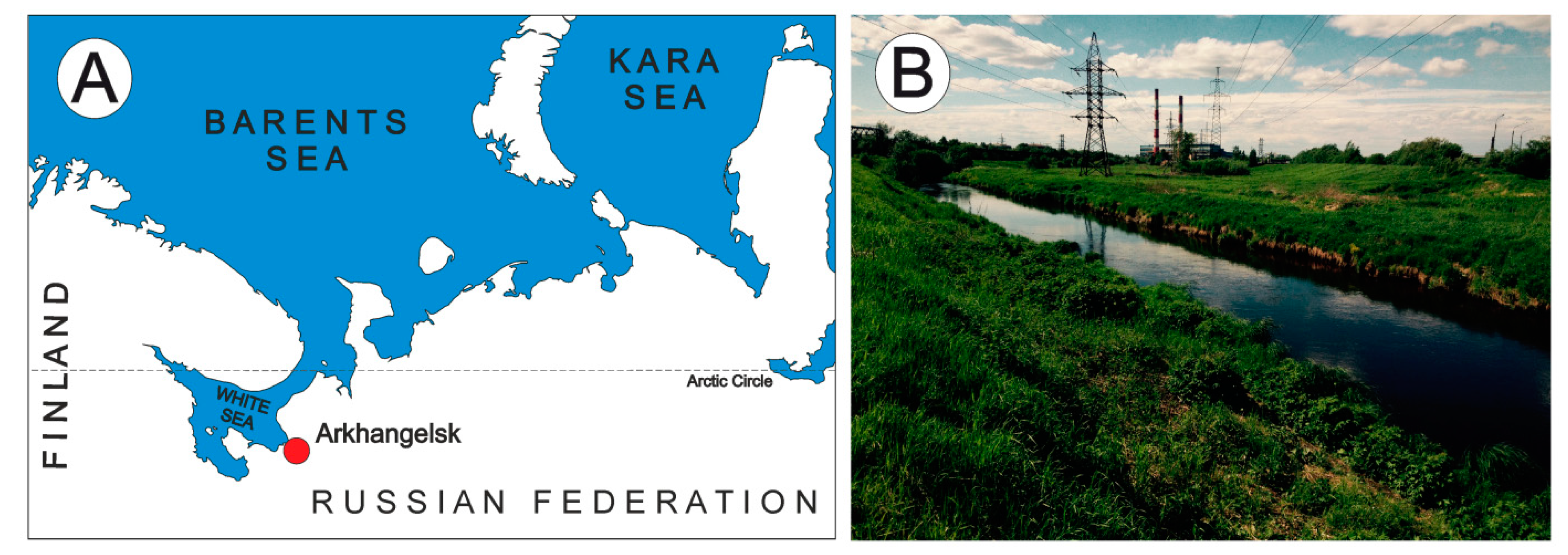
2.1. Study Area
2.2. Field Sampling
2.3. Morphological and Anatomical Studies
2.4. Histological Examination
2.5. Statistical Analyses
3. Results
3.1. Habitat Characteristics
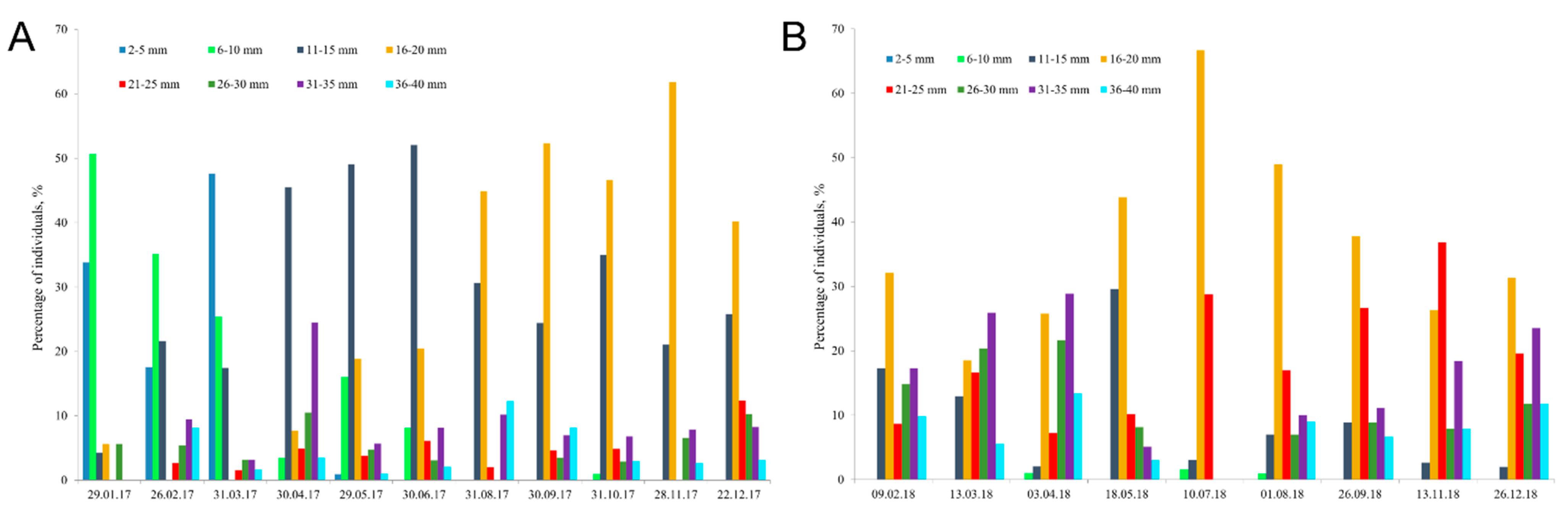
3.2. Shell Length Structure and Brooding
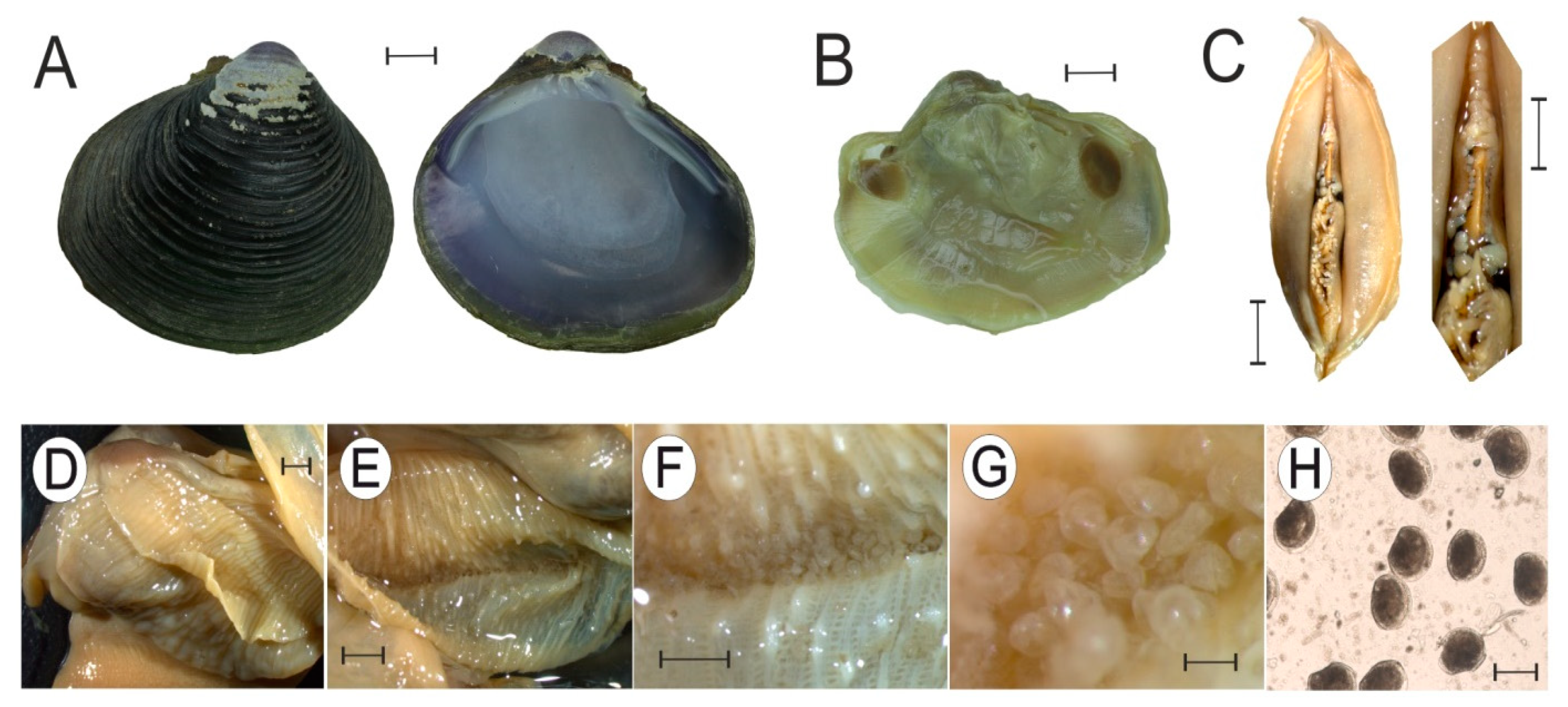
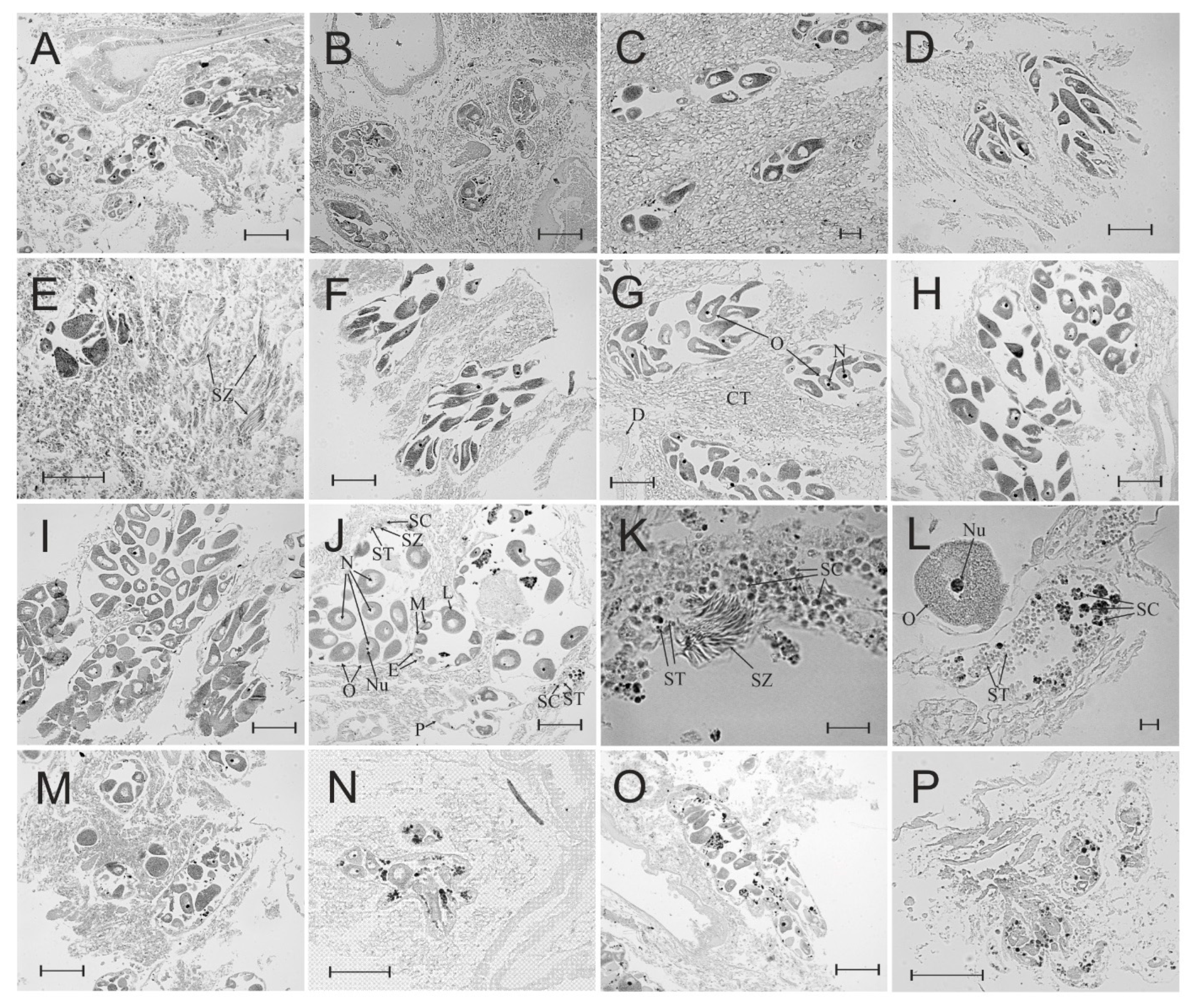
3.3. Gonad Histology
4. Discussion
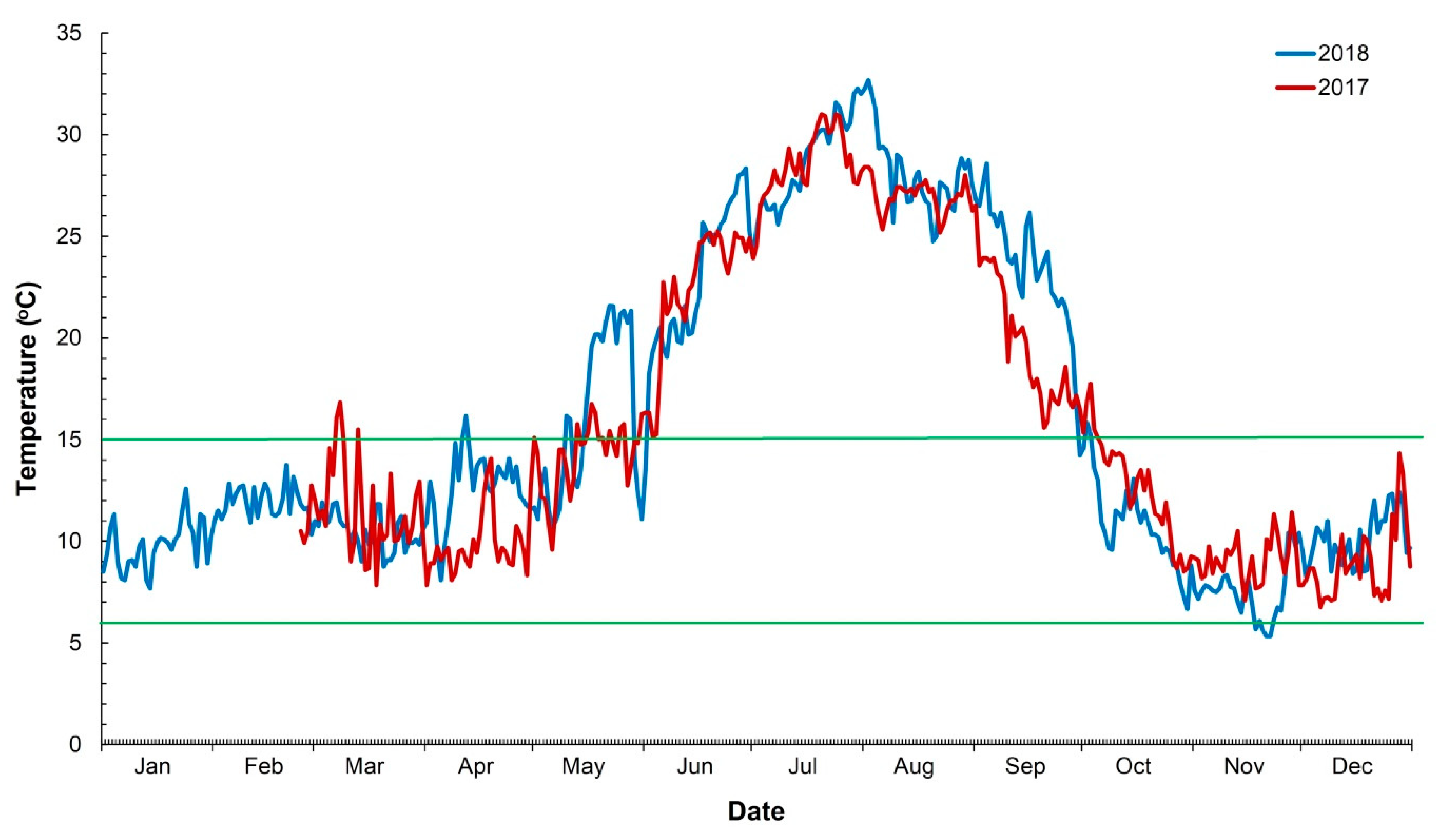
4.1. Reproduction and Influence of Water Temperature
4.2. Reproductive Strategies and Gonadal Histology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Glaubrecht, M.; Von Rintelen, T.; Korniushin, A.V. Towards a systematic revision of brooding freshwater Corbiculidae in southeast Asia (Bivalvia, Veneroida): On shell morphology, anatomy and molecular phylogenetics of endemic taxa from islands in Indonesia. Malacologica 2003, 45, 1–40. [Google Scholar]
- Siripattrawan, S.; Park, J.K.; Foighil, D.O. Two lineages of the introduced Asian freshwater clam Corbicula occur in North America. J. Molluscan Stud. 2000, 66, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Korniushin, A.V. A revision of some Asian and African freshwater clams assigned to Corbicula fluminalis (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae), with a review of anatomical characters and reproductive features based on museum collections. Hydrobiologia 2004, 529, 251–270. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Stanger-hall, K.; Baker, R.J.; Hillis, D.M. All male asexuality: Origin and maintenance of androgenesis in the Asian clam Corbicula. Evolution 2008, 62, 1119–1136. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, L.-M.; Etoundi, E.; Aldridge, D.C.; Marescaux, J.; Yasuda, N.; Van Doninck, K. Genetic uniformity and long-distance clonal dispersal in the invasive androgenetic Corbicula clams. Mol. Ecol. 2014, 23, 5102–5116. [Google Scholar] [CrossRef]
- Crespo, D.; Dolbeth, M.; Leston, S.; Sousa, R.; Pardal, A. Distribution of Corbicula fluminea (Müller, 1774) in the invaded range: A geographic approach with notes on species traits variability. Biol. Invasions 2015, 17, 2087–2101. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Padilla, D.K.; Minchin, D.; Boltovskoy, D.; Burlakova, L.E. Changes in global economies and trade: The potential spread of exotic freshwater bivalves. Biol. Invasions 2007, 9, 161–180. [Google Scholar] [CrossRef]
- Hakenkamp, C.C.; Ribblett, S.G.; Palmer, M.A.; Swan, C.M.; Reid, J.W.; Goodison, M.R. The impact of an introduced bivalve (Corbicula fluminea) on the benthos of a sandy stream. Freshw. Biol. 2001, 46, 491–501. [Google Scholar] [CrossRef]
- Sousa, R.; Antunes, C.; Guilhermo, L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: An overview. Ann. Limnol. Int. J. Limnol. 2008, 44, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Lucy, F.E.; Karatayev, A.Y.; Burlakova, L.E. Predictions for the spread, population density, and impacts of Corbicula fluminea in Ireland. Aquat Invasions 2012, 7, 465–474. [Google Scholar] [CrossRef]
- Sousa, R.; Gutiérrez, J.; Aldridge, D. Non-indigenous invasive bivalves as ecosystem engineers. Biol. Invasions 2009, 11, 2367–2385. [Google Scholar] [CrossRef]
- Cao, L.; Damborenea, C.; Penchaszadeh, P.E.; Darrigran, G. Gonadal cycle of Corbicula fluminea (Bivalvia: Corbiculidae) in Pampean streams (Southern Neotropical Region). PLoS ONE 2017, 12, e0186850. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Foighil, D.F. Invasion of the clonal clams: Corbicula lineages in the New World. Am. Malacol. Bull. 2005, 20, 113–122. [Google Scholar]
- Weitere, M.; Yohmann, A.; Schulz, N.; Linn, C.; Dietrich, D.; Arndt, H. Linking environmental warming to the fitness of the invasive clam Corbicula fluminea. Glob. Chang. Biol. 2009, 15, 2838–2851. [Google Scholar] [CrossRef]
- Mcdowell, W.; Mcdowell, W.H.; Byers, J.E. Mass mortality of a dominant invasive species in response to an extreme climate event: Implications for ecosystem function. Limnol. Oceanogr. 2016, 62, 177–188. [Google Scholar] [CrossRef]
- Pauchard, A.; Milbau, A.; Albihn, A. Non-native and native organisms moving into high elevation and high latitude ecosystems in an era of climate change: New challenges for ecology and conservation. Biol. Invasions 2016, 18, 345–353. [Google Scholar] [CrossRef]
- Bódis, E.; Nosek, J.; Oeretel, N.; Toth, B.; Feher, Z. Comparative study of two Corbicula morphs (Bivalvia, Corbiculidae) Inhabiting River Danube. Int. Rev. Hydrobiol. 2011, 96, 257–273. [Google Scholar] [CrossRef]
- Bryan, N.J.; Florence, C.V.; Crail, T.D.; Moorhead, D.L. Freshwater mussel community response to warm water discharge in western Lake Erie. J. Great Lakes Res. 2013, 39, 449–454. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Andreev, N.; Andreeva, S.; Kazantsev, I.E.; Karimov, A.V.; Lazutkina, E.A. Alien mollusk species in the aquatic ecosystems of Western Siberia: A review. Russ. J. Biol. Invasions 2015, 6, 137–147. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Bolotov, I.N.; Aksenova, O.V.; Kondakov, A.V.; Gofarov, M.Y.; Laenko, E.; Sokolova, S.E.; Shevchenko, A.R.; Travina, O.V. Aliens are moving to the Arctic frontiers: An integrative approach reveals selective expansion of androgenic hybrid Corbicula lineages towards the North of Russia. Biol. Invasions 2018, 20, 2227–2243. [Google Scholar] [CrossRef]
- Castañeda, A.; Cvetanovska, E.; Kayla, M.; Hamelin, M.; Simard, A.; Ricciardi, A. Distribution, abundance and condition of an invasive bivalve (Corbicula fluminea) along an artificial thermal gradient in the St. Lawrence River. Aquat. Invasions 2018, 13, 379–392. [Google Scholar] [CrossRef]
- Zhivoglyadova, L.A.; Revkov, N.K. Invasion of the bivalve Corbicula fluminea (Müller, 1774) (Bivalvia: Cyrenidae) into the lower Don basin. Aquat. Bioresour. Environ. 2018, 1, 44–50. [Google Scholar] [CrossRef]
- Pryanichnikova, E.G.; Voroshilova, I.S.; Sabitova, R.Z. Introduction of Corbicula fluminea (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae) in the Volga River Basin. Inland Water Biol. 2019, 12, 95–97. [Google Scholar] [CrossRef]
- Etoundi, E.; Marescaux, J.; Yastrade, M.; Debortoli, N.; Hedtke, S.M.; Pigneur, L.-M.; Virgo, J.; Flot, J.F.; Van Doninck, K. Distinct biogeographic origins of androgenetic Corbicula lineages followed by genetic captures. bioRxiv 2019, 590836. [Google Scholar] [CrossRef] [Green Version]
- Pigneur, L.-M.; Marescaux, J.; Roland, K.; Etoundi, E.; Descy, J.-P.; Van Doninck, K. Phylogeny and androgenesis in the invasive Corbicula clams (Bivalvia, Corbiculidae) in Western-Europe. BMC Evol. Biol. 2011, 11, 147. [Google Scholar] [CrossRef] [Green Version]
- Hedtke, S.M.; Glaubrecht, M.; Hillis, D.M. Rare gene capture in predominantly androgenetic species. Proc. Natl. Acad. Sci. USA 2011, 108, 9520–9524. [Google Scholar] [CrossRef] [Green Version]
- Morton, B. The population dynamics of Corbicula fluminea (Bivalvia: Corbiculacea) in Plover Cove Reservoir, Hong Kong. J. Zool. 1997, 181, 21–42. [Google Scholar] [CrossRef]
- Hornbach, D.J. Life history traits of a riverine population of the Asian clam Corbicula fluminea. Am. Midl. Nat. 1992, 127, 248–257. [Google Scholar] [CrossRef]
- Rajagopal, S.; Van Der Velde, G.; Bij De Vaate, D. Reproductive biology of the Asiatic clams Corbicula fluminalis and Corbicula fluminea in the River Rhine. Arch. Fur Hydrobiol. 2000, 149, 403–420. [Google Scholar] [CrossRef]
- Kraemer, L.; Galloway, M. Larval development of Corbicula fluminea (Muller) (Bivalvia: Corbiculacea): An appraisal of its heterochrony. Am. Malacol. Bull. 1986, 4, 61–79. [Google Scholar]
- Mouthon, J. Life cycle and population dynamics of the Asian clam Corbicula fluminea (Bivalvia: Corbiculidae) in the Rhône River at Creys-Malville (France). Arch. Fur Hydrobiol. 2001, 151, 571–589. [Google Scholar] [CrossRef]
- Cataldo, D.; Boltovskoy, D. Population dynamics of Corbicula fluminea (Bivalvia) in the Paraná river delta (Argentina). Hydrobiologia 1999, 380, 153–163. [Google Scholar] [CrossRef]
- McMahon, R.F. Evolutionary and physiological adaptations of aquatic invasive animals: R selection versus resistance. Can. J. Fish. Aquat. Sci. 2002, 59, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- French, J., III; Schloesser, D. Growth and overwinter survival of the Asiatic clam, Corbicula fluminea, in the St. Clair River, Michigan. Hydrobiologia 1991, 219, 165–170. [Google Scholar] [CrossRef]
- Denton, M.E.; Chandra, S.; Wittmann, M.E.; Reuter, J.; Baguley, J.G. Reproduction and Population Structure of Corbicula fluminea in an Oligotrophic Subalpine Lake. J. Shellfish Res. 2012, 31, 145–152. [Google Scholar] [CrossRef]
- Penk, M.R.; Williams, M.A. Thermal effluents from power plants boost performance of the invasive clam Corbicula fluminea in Ireland’s largest river. Sci. Total Environ. 2019, 693, 133546. [Google Scholar] [CrossRef] [PubMed]
- Marescaux, J.; Pigneur, L.-M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat Invasions 2010, 5, 35–39. [Google Scholar] [CrossRef]
- Agafonova, S.A.; Frolova, N.L. Specific features of ice regime in rivers of the Northern Dvina basin. Water Resour. 2007, 34, 123–131. [Google Scholar] [CrossRef]
- Den Hartog, C.; Van Den Brink, F.B.W.; Van Der Velde, G. Why was the invasion of the River Rhine by Corophium curvispinum and Corbicula species so successful? J. Shellfish Res. 1992, 26, 1121–1129. [Google Scholar]
- Reyna, P.B.; Morán, A.G.; Tatián, M. Taxonomy, distribution and population structure of invasive Corbiculidae (Mollusca, Bivalvia) in the Suquía River basin, Córdoba, Argentina. Iheringia Série Zool. 2013, 103, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Baba, K.; Tada, M.; Kawajiri, Y. Effects of temperature and salinity on spawning of the brackish water bivalve Corbicula japonica in Lake Abashiri, Hokkaido, Japan. Mar. Ecol. Prog. Ser. 1999, 180, 213–221. [Google Scholar] [CrossRef]
- Rybalkina (Dzyuba), S.M.; Maiorova, M.A.; Anisimov, A.P.; Kravchenko, D.N. The Gametogenesis and Sexual Cycle of the Bivalve Corbicula japonica Prime (1864) in the Mouth of the Kievka River (Sea of Japan). Russ. J. Mar. Biol. 2013, 39, 253–264. [Google Scholar] [CrossRef]
- Morton, B. Some aspects of the population structure and sexual strategy of Corbicula cf. fluminalis (Bivalvia: Corbiculacea) from the Pearl River, Peoples Republic of China. J. Molluscan Stud. 1982, 48, 1–23. [Google Scholar]
- Juhel, G.; Culloty, S.; O’riordan, R.; O’connor, J.; Faoite, L.; McNamara, R. A histological study of the gametogenic cycle of the freshwater mussel Dreissena polymorpha (Pallas, 1771) in Lough Derg, Ireland. J. Molluscan Stud. 2003, 69, 365–373. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Doherty, F.G.; Cherry, D.S.; Cairns, J.R. Spawning periodicity of the Asiatic clam Corbicula fluminea in the New River, Virginia. Am. Midl. Nat. 1987, 117, 71–82. [Google Scholar] [CrossRef]
- Park, G.; Chung, E.Y. Histological studies on hermaphroditism, gametogenesis and cyclic changes in the structures of marsupial gills of the introduced Asiatic clam, Corbicula fluminea and the Korean clam, Corbicula leana. J. Shellfish Res. 2004, 23, 179–184. [Google Scholar]
- Gomes, C.; Sousa, R.; Mendes, T.; Borges, R.; Vilares, P.; Vasconcelos, V.; Guilhermino, L.; Antunes, A. Low genetic diversity and high invasion success of Corbicula fluminea (Bivalvia, Corbiculidae) (Müller, 1774) in Portugal. PLoS ONE 2016, 11, e0158108. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, V.S.; Huekelem, L.V. Gametogenesis and larval production in a population of the introduced Asiatic Clam Corbicula sp. (Bivalvia: Corbiculidae), in Maryland. Biol. Bull. 1985, 168, 50–60. [Google Scholar] [CrossRef]
- Byrne, M.; Phelps, H.; Church, T.; Adair, V.; Selvakumaraswamy, P.; Potts, J. Reproduction and development of the freshwater clam Corbicula australis in southeast Australia. Hydrobiologia 2000, 418, 185–197. [Google Scholar] [CrossRef]
- Labecka, A.M.; Domagala, J. Continuous reproduction of Sinanodonta woodiana (Lea, 1824) females: An invasive mussel species in a female-biased population. Hydrobiologia 2018, 810, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Beninger, P. Caveat observator: The many faces of pre-spawning atresia in marine bivalve reproductive cycles. Mar. Biol. 2017, 164, 163. [Google Scholar] [CrossRef]
- Dorange, G.; Pennec, M.L.E. Ultrastructural study of oogenesis and oocytic degeneration in Pecten maxiums from the Bay of St. Brieuc. Mar. Biol. 1989, 103, 339–348. [Google Scholar] [CrossRef]
- Lima, P.; Monteiro, S.M.; Sousa, M.; Machado, J. A histological study of oogenesis in the freshwater mussel Anodonta cygnea (Linnaeus, 1758) in Mira Lagoon, Portugal. Malacologia 2012, 55, 251–261. [Google Scholar] [CrossRef]
- Darriba, S.; San Juan, F.; Guerra, A. Reproductive cycle of the razor clam Ensis arcuatus (Jeffreys, 1865) in northwest Spain and its relation to environmental conditions. J. Exp. Mar. Biol. Ecol. 2004, 311, 101–115. [Google Scholar] [CrossRef]
- Şehriban, C.; Şereflişan, H. Certain reproductive characteristics of the freshwater mussel Unio terminalis delicates (Lea, 1863) in Gölbaşi. Aquat. Sci. 2002, 59, 1235–1244. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bespalaya, Y.V.; Aksenova, O.V.; Kropotin, A.V.; Shevchenko, A.R.; Travina, O.V. Reproduction of the Androgenetic Population of the Asian Corbicula Clam (Bivalvia: Cyrenidae) in the Northern Dvina River Basin, Russia. Diversity 2021, 13, 316. https://doi.org/10.3390/d13070316
Bespalaya YV, Aksenova OV, Kropotin AV, Shevchenko AR, Travina OV. Reproduction of the Androgenetic Population of the Asian Corbicula Clam (Bivalvia: Cyrenidae) in the Northern Dvina River Basin, Russia. Diversity. 2021; 13(7):316. https://doi.org/10.3390/d13070316
Chicago/Turabian StyleBespalaya, Yulia V., Olga V. Aksenova, Alexander V. Kropotin, Alexander R. Shevchenko, and Oksana V. Travina. 2021. "Reproduction of the Androgenetic Population of the Asian Corbicula Clam (Bivalvia: Cyrenidae) in the Northern Dvina River Basin, Russia" Diversity 13, no. 7: 316. https://doi.org/10.3390/d13070316