
Revisiting the Diversity of Barbonymus (Cypriniformes, Cyprinidae) in Sundaland Using DNA-Based Species Delimitation Methods

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
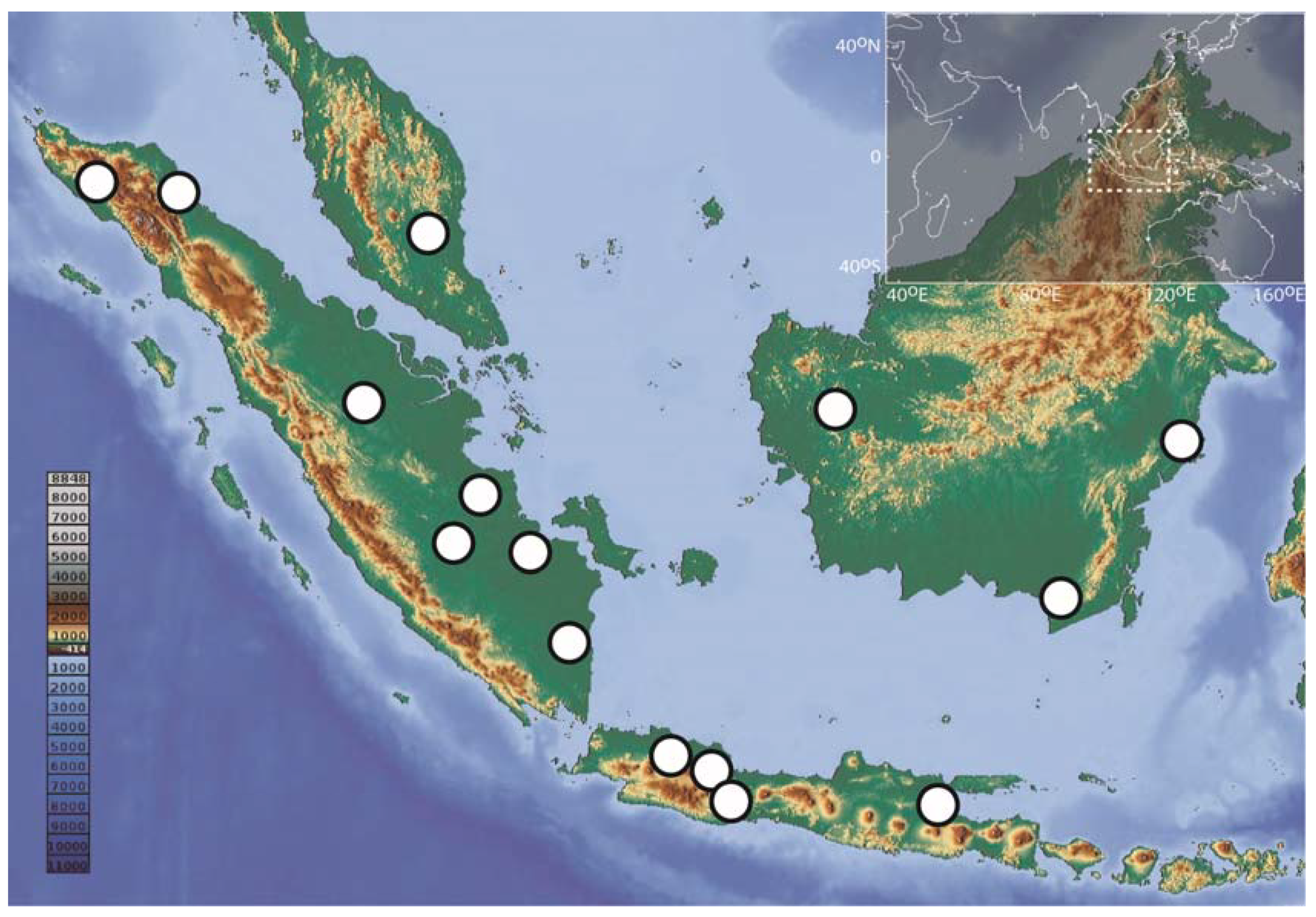
2.1. Sampling and Collection Management
2.2. Sequencing and International Repositories
2.3. Genetic Species Delimitation and Phylogenetic Inferences
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, F. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar]
- Hubert, N.; Wibowo, A.; Busson, F.; Caruso, D.; Sulandari, S.; Nafiqoh, N.; Pouyaud, L.; Rüber, L.; Avarre, J.C.; Herder, F.; et al. DNA barcoding Indonesian freshwater fishes: Challenges and prospects. DNA Barcodes 2015, 3, 144–169. [Google Scholar] [CrossRef] [Green Version]
- Imai, N.; Furukawa, T.; Tsujino, R.; Kitamura, S.; Yumoto, T. Factors affecting forest area change in Southeast Asia during 1980–2010. PLoS ONE 2018, 13, e0197391. [Google Scholar] [CrossRef] [Green Version]
- Laumonier, Y.; Uryu, Y.; Stüwe, M.; Budiman, A.; Setiabudi, B.; Hadian, O. Eco-floristic sectors and deforestation threats in Sumatra: Identifying new conservation area network priorities for ecosystem-based land use planning. Biodivers. Conserv. 2010, 19, 1153–1174. [Google Scholar] [CrossRef]
- Gaveau, D.L.A.; Sloan, S.; Molidena, E.; Yaen, H.; Sheil, D.; Abram, N.K.; Ancrenaz, M.; Nasi, R.; Quinones, M.; Wielaard, N. Four decades of forest persistence, clearance and logging on Borneo. PLoS ONE 2014, 9, e101654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, T.; Hamilton, S.E.; Hochard, J.P.; Kresch, E.P.; Talbot, J. (Not so) gently down the stream: River pollution and health in Indonesia. J. Environ. Econ. Manag. 2018, 92, 35–53. [Google Scholar] [CrossRef]
- Mulligan, M.; van Soesbergen, A.; Sáenz, L. GOODD, a global dataset of more than 38,000 georeferenced dams. Sci. Data 2020, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahruddin, H.; Hutama, A.; Busson, F.; Sauri, S.; Hanner, R.; Keith, P.; Hadiaty, R.; Hubert, N. Revisiting the ichthyodiversity of Java and Bali through DNA barcodes: Taxonomic coverage, identification accuracy, cryptic diversity and identification of exotic species. Mol. Ecol. Resour. 2017, 17, 288–299. [Google Scholar] [CrossRef]
- FAO. Des Pêches et de L’aquaculture; FAO: Rome, Italy, 2018; ISBN 9789251306925. [Google Scholar]
- Koeshendrajana, S.; Cacho, O.J. Management Options for the Inland Fisheries Resource in South Sumatra, Indonesia: I Bioeconomic Model; Working Papers 12932; University of New England: Armidale, Australia, 2001. [Google Scholar]
- Coates, D. Inland capture fishery statistics of Southeast Asia: Current status and information needs. RAP Publ. 2002, 11, 114. [Google Scholar]
- Welcomme, R.L.; Baird, I.G.; Dudgeon, D.; Halls, A.; Lamberts, D.; Mustafa, M.G. Fisheries of the Rivers of Southeast Asia; Jon Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 363–376. [Google Scholar]
- Muthmainnah, D.; Makmur, H.R.; Sawestri, S.; Supriyadi, F.; Fatah, K. The Features of Inland Fisheries in Southeast Asia: Inland Capture Fisheries and Its Status; Inland Fishery Resources Development and Management Department, Southeast: Samut Prakan, Thailand, 2019; ISBN 6024408161. [Google Scholar]
- Conte-Grand, C.; Britz, R.; Dahanukar, N.; Raghavan, R.; Pethiyagoda, R.; Tan, H.H.; Hadiaty, R.K.; Yaakob, N.S.; Rüber, L. Barcoding snakeheads (Teleostei, Channidae) revisited: Discovering greater species diversity and resolving perpetuated taxonomic confusions. PLoS ONE 2017, 12, e0184017. [Google Scholar] [CrossRef]
- Batubara, A.S.; Muchlisin, Z.A.; Efizon, D.; Elvyra, R.; Fadli, N.; RIZAL, S.; Siti-Azizah, M.N.; Wilkes, M. DNA barcoding (COI genetic marker) revealed hidden diversity of Cyprinid fish (Barbonymus spp.) from Aceh Waters, Indonesia. Biharean Biol. 2013, 15, 1. [Google Scholar]
- Kottelat, M. The fishes of the inland waters of Southeast Asia: A catalog and core bibliography of the fishes known to occur in freshwaters, mangroves and estuaries. Raffles Bull. Zool. 2013, (Suppl. 27), 1–663. [Google Scholar]
- Froese, R.; Pauly, D. Fishbase. Available online: http://www.fishbase.org (accessed on 15 January 2021).
- Eschmeyer, W.N.; Fricke, R.; van der Laan, R. Catalog of Fishes Electronic Version. Available online: http://www.calacademy.org/research/ichthyology/catalog/fishcatsearch.html (accessed on 15 January 2021).
- Kottelat, M.; Whitten, A.J.; Kartikasari, N.; Wirjoatmodjo, S. Freshwater Fishes of Western Indonesia and Sulawesi; Periplus Publishing Group: Hong Kong, China, 1993; ISBN 0945971605. [Google Scholar]
- Hubert, N.; Lumbantobing, D.; Sholihah, A.; Dahruddin, H.; Delrieu-Trottin, E.; Busson, F.; Sauri, S.; Hadiaty, R.; Keith, P. Revisiting species boundaries and distribution ranges of Nemacheilus spp. (Cypriniformes: Nemacheilidae) and Rasbora spp. (Cypriniformes: Cyprinidae) in Java, Bali and Lombok through DNA barcodes: Implications for conservation in a biodiversity hotspot. Conserv. Genet. 2019, 20, 517–529. [Google Scholar] [CrossRef]
- Sholihah, A.; Delrieu-Trottin, E.; Sukmono, T.; Dahruddin, H.; Risdawati, R.; Elvyra, R.; Wibowo, A.; Kustiati, K.; Busson, F.; Sauri, S.; et al. Disentangling the taxonomy of the subfamily Rasborinae (Cypriniformes, Danionidae) in Sundaland using DNA barcodes. Sci. Rep. 2020, 10, 2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.-C.; Abidin, M.Z.; Pulungan, C.P.; De Bruyn, M.; Mohd Nor, S.A. DNA barcoding reveals high cryptic diversity of freshwater halfbeak genus Hemirhamphodon from Sundaland. PLoS ONE 2016, 11, e0163596. [Google Scholar] [CrossRef] [PubMed]
- Beck, S.V.; Carvalho, G.R.; Barlow, A.; Ruber, L.; Hui Tan, H.; Nugroho, E.; Wowor, D.; Mohd Nor, S.A.; Herder, F.; Muchlisin, Z.A.; et al. Plio-Pleistocene phylogeography of the Southeast Asian Blue Panchax killifish, Aplocheilus panchax. PLoS ONE 2017, 12, e0179557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurul Farhana, S.; Muchlisin, Z.A.; Duong, T.Y.; Tanyaros, S.; Page, L.M.; Zhao, Y.; Adamson, E.A.S.; Khaironizam, M.Z.; de Bruyn, M.; Siti Azizah, M.N. Exploring hidden diversity in Southeast Asia’s Dermogenys spp. (Beloniformes: Zenarchopteridae) through DNA barcoding. Sci. Rep. 2018, 8, 10787. [Google Scholar] [CrossRef] [Green Version]
- Hutama, A.; Dahruddin, H.; Busson, F.; Sauri, S.; Keith, P.; Hadiaty, R.K.; Hanner, R.; Suryobroto, B.; Hubert, N. Identifying spatially concordant evolutionary significant units across multiple species through DNA barcodes: Application to the conservation genetics of the freshwater fishes of Java and Bali. Glob. Ecol. Conserv. 2017, 12, 170–187. [Google Scholar] [CrossRef]
- Delrieu-Trottin, E.; Durand, J.; Limmon, G.; Sukmono, T.; Sugeha, H.Y.; Chen, W.; Busson, F.; Borsa, P.; Dahruddin, H.; Sauri, S. Biodiversity inventory of the grey mullets (Actinopterygii: Mugilidae) of the Indo-Australian Archipelago through the iterative use of DNA-based species delimitation and specimen assignment methods. Evol. Appl. 2020, 13, 1451–1467. [Google Scholar] [CrossRef] [Green Version]
- Sholihah, A.; Delrieu-Trottin, E.; Condamine, F.L.; Wowor, D.; Rüber, L.; Pouyaud, L.; Agnèse, J.-F.; Hubert, N. Impact of Pleistocene Eustatic Fluctuations on Evolutionary Dynamics in Southeast Asian Biodiversity Hotspots. Syst. Biol. 2021. [Google Scholar] [CrossRef]
- Rahman, M.M.; Norén, M.; Mollah, A.R.; Kullander, S.O. Building a DNA barcode library for the freshwater fishes of Bangladesh. Sci. Rep. 2019, 9, 9382. [Google Scholar] [CrossRef] [PubMed]
- Aquino, L.M.G.; Tango, J.M.; Canoy, R.J.C.; Fontanilla, I.K.C.; Basiao, Z.U.; Ong, P.S.; Quilang, J.P. DNA barcoding of fishes of Laguna de Bay, Philippines. Mitochondrial DNA 2011, 22, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Meganathan, P.; Austin, C.M.; Tam, S.M.; Chew, P.C.; Siow, R.; Rashid, Z.A.; Song, B.K. An Application of DNA Barcoding to the Malaysian Freshwater Fish Fauna: mtDNA COI Sequences Reveal Novel Haplotypes, Cryptic Species and Field-Based Misidentification; Monash University Malaysia: Selangor, Malaysia, 2015. [Google Scholar]
- Panprommin, D.; Soontornprasit, K.; Tuncharoen, S.; Iamchuen, N. Efficacy of DNA barcoding for the identification of larval fish species in the Upper and Middle Ing River, Thailand. Gene Rep. 2021, 23, 101057. [Google Scholar] [CrossRef]
- Barman, A.S.; Singh, M.; Singh, S.K.; Saha, H.; Singh, Y.J.; Laishram, M.; Pandey, P.K. DNA barcoding of freshwater fishes of Indo-Myanmar biodiversity hotspot. Sci. Rep. 2018, 8, 8579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barman, A.S.; Singh, M.; Pandey, P.K. DNA barcoding and genetic diversity analyses of fishes of Kaladan River of Indo-Myanmar biodiversity hotspot. Mitochondrial DNA Part A 2018, 29, 367–378. [Google Scholar] [CrossRef]
- Esa, Y.B.; Siraj, S.S.; Daud, S.K.; Ryan, J.J.R.; Rahim, K.A.A.; Tan, S.G. Molecular systematics of mahseers (Cyprinidae) in Malaysia inferred from sequencing of a mitochondrial Cytochrome C Oxidase I (COI) gene. Pertanika J. Trop. Agric. Sci. 2008, 31, 263–269. [Google Scholar]
- Lakra, W.S.; Singh, M.; Goswami, M.; Gopalakrishnan, A.; Lal, K.K.; Mohindra, V.; Sarkar, U.K.; Punia, P.P.; Singh, K.V.; Bhatt, J.P. DNA barcoding Indian freshwater fishes. Mitochondrial DNA Part A 2016, 27, 4510–4517. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hébert, P.D.N. Universal primers cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; deWaard, J.R.; Zakharov, E.; Prosser, S.W.J.; Sones, J.E.; McKeown, J.T.A.; Mantle, B.; La Salle, J. A DNA “barcode blitz”: Rapid digitization and sequencing of a natural history collection. PLoS ONE 2013, 8, e68535. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The barcode index number (BIN) system. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapli, P.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson Tree Processes for single-locus species delimitation under Maximum Likelihood and Markov Chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, A.; Ling, C.; Ho, S.Y.W.; Zhu, C.-D. Comparison of methods for molecular species delimitation across a range of speciation scenarios. Syst. Biol. 2018, 67, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Kekkonen, M.; Mutanen, M.; Kaila, L.; Nieminen, M.; Hebert, P.D.N. Delineating Species with DNA Barcodes: A Case of Taxon Dependent Method Performance in Moths. PLoS ONE 2015, 10, e0122481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kekkonen, M.; Hebert, P.D.N. DNA barcode-based delineation of putative species: Efficient start for taxonomic workflows. Mol. Ecol. Resour. 2014, 14, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Hubert, N.; Huang, Y.; Wang, X.; Gan, X.; Peng, Z.; He, S. DNA barcoding the ichthyofauna of the Yangtze River: Insights from the molecular inventory of a mega-diverse temperate fauna. Mol. Ecol. Resour. 2019, 19, 1278–1291. [Google Scholar] [CrossRef]
- Limmon, G.; Delrieu-Trottin, E.; Patikawa, J.; Rijoly, F.; Dahruddin, H.; Busson, F.; Steinke, D.; Hubert, N. Assessing species diversity of Coral Triangle artisanal fisheries: A DNA barcode reference library for the shore fishes retailed at Ambon harbor (Indonesia). Ecol. Evol. 2020, 10, 3356–3366. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the generalized mixed Yule coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermingham, E.; McCafferty, S.; Martin, A.P. Fish biogeography and molecular clocks: Perspectives from the Panamanian isthmus. In Molecular Systematics of Fishes; Kocher, T.D., Stepien, C.A., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 113–128. [Google Scholar]
- Ogilvie, H.A.; Bouckaert, R.R.; Drummond, A.J. StarBEAST2 brings faster species tree inference and accurate estimates of substitution rates. Mol. Biol. Evol. 2017, 34, 2101–2114. [Google Scholar] [CrossRef]
- Ho, S.Y.W.; Larson, G. Molecular clocks: When timesare a-changin’. Trends Genet. 2006, 22, 79–83. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide-Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Brown, S.D.J.; Collins, R.A.; Boyer, S.; Lefort, C.; Malumbres-Olarte, J.; Vink, C.J.; Cruickshank, R.H. Spider: An R package for the analysis of species identity and evolution, with particular reference to DNA barcoding. Mol. Ecol. Resour. 2012, 12, 562–565. [Google Scholar] [CrossRef]
- Paradis, E. pegas: An {R} package for population genetics with an integrated--modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, C.; Bryson, J.R.W. Cryptic diversity and discordance in single-locus species delimitation methods within horned lizards (Phrynosomatidae: Phrynosoma). Mol. Ecol. Resour. 2017, 17, 1168–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Liu, S.; Yu, D.; Liu, H.; Danley, P.D. Mitochondrial capture and incomplete lineage sorting in the diversification of balitorine loaches (Cypriniformes, Balitoridae) revealed by mitochondrial and nuclear genes. Zool. Scr. 2012, 41, 233–247. [Google Scholar] [CrossRef]
- Hopkins, R.L.; Eisenhour, D.J. Hybridization of Lythrurus fasciolaris and Lythrurus umbratilis (Cypriniformes: Cyprinidae) in the Ohio River basin. Copeia 2008, 2008, 162–171. [Google Scholar] [CrossRef]
- Atsumi, K.; Nomoto, K.; Machida, Y.; Ichimura, M.; Koizumi, I. No reduction of hatching rates among F1 hybrids of naturally hybridizing three Far Eastern daces, genus Tribolodon (Cypriniformes, Cyprinidae). Ichthyol. Res. 2018, 65, 165–167. [Google Scholar] [CrossRef]
- Hubert, N.; Hanner, R. DNA barcoding, species delineation and taxonomy: A historical perspective. DNA Barcodes 2015, 3, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Wibowo, A.; Atminarso, D.; Baumgartner, L.; Vasemagi, A. High prevalence of non-native fish species in a remote region of the Mamberamo River, Indonesia. Pac. Conserv. Biol. 2020, 26, 293–300. [Google Scholar] [CrossRef]
- Keith, P.; Lord, C.; Darhuddin, H.; Limmon, G.; Sukmono, T.; Hadiaty, R.; Hubert, N. Schismatogobius (Gobiidae) from Indonesia, with description of four new species. Cybium 2017, 41, 195–211. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Original Description | Authors | Eschmeyer Catalog of Fished | Fishbase | Status |
|---|---|---|---|---|
| Barbus altus | Günther 1868 | Barbonymus altus | Barbonymus altus | Valid as Barbonymus altus (Günther 1868) |
| Barbus amblycephalus | Bleeker 1855 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus balleroides | Valenciennes 1842 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus boulengerii | Popta 1905 | Barbonymus collingwoodii | Barbonymus collingwoodii | Valid as Barbonymus collingwoodii (Günther 1868) |
| Barbus bramoides | Valenciennes 1842 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus erythropterus | Bleeker 1849 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus foxi | Fowler 1937 | Barbonymus altus | Barbonymus altus | Valid as Barbonymus altus (Günther 1868) |
| Barbus gonionotus | Bleeker 1849 | Barbonymus gonionotus | Barbonymus gonionotus | Valid as Barbonymus gonionotus (Bleeker 1849) |
| Barbus hypsylonotus | Valenciennes 1842 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus javanicus | Bleeker 1855 | Barbonymus gonionotus | Barbonymus gonionotus | Valid as Barbonymus gonionotus (Bleeker 1849) |
| Barbus koilometopon | Bleeker 1857 | Barbonymus gonionotus | Barbonymus gonionotus | Valid as Barbonymus gonionotus (Bleeker 1849) |
| Barbus macrophthalmus | Bleeker 1855 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Barbus mahakkamensis | Ahl, 1922 | Barbonymus mahakkamensis | Barbodes mahakkamensis | Valid as Barbus mahakkamensis (Ahl 1922) |
| Barbus platysoma | Bleeker 1855 | Barbonymus platysoma | Barbodes platysoma | Valid as Barbodes platysoma (Bleeker 1855) |
| Barbus schwanefeldi rubra | Vaillant 1902 | Barbonymus schwanefeldii | Barbonymus schwanenfeldii | Valid as Barbonymus schwanefeldii (Bleeker 1854) |
| Barbus schwanenfeldii | Bleeker 1854 | Barbonymus schwanefeldii | Barbonymus schwanenfeldii | Valid as Barbonymus schwanefeldii (Bleeker 1854) |
| Barbus strigatus | Boulenger 1894 | Barbonymus strigatus | Barbodes strigatus | Valid as Barbonymus strigatus (Boulenger 1894) |
| Barbus wadon | Bleeker 1849 | Barbonymus balleroides | Barbonymus balleroides | Valid as Barbonymus balleroides (Valenciennes 1842) |
| Puntius jolamarki | Smith 1934 | Barbonymus gonionotus | Barbonymus gonionotus | Valid as Barbonymus gonionotus (Bleeker 1849) |
| Puntius viehoeveri | Fowler 1943 | Barbonymus gonionotus | Barbonymus gonionotus | Valid as Barbonymus gonionotus (Bleeker 1849) |
| Systomus belinka | Bleeker 1860 | Barbonymus belinka | Puntius belinka | Valid as Barbonymus belinka (Bleeker 1860) |
| K2P Genetic Distance | ||||
|---|---|---|---|---|
| Species | N | BIN | Max. Intraspecific | Min. Interspecific |
| Barbonymus altus | 17 | BOLD:AAE2136 | 0.010 | 0.026 |
| 1 | BOLD:AEB4313 | - | 0.026 | |
| Barbonymus belinka | 5 | BOLD:AED2516 | 0.000 | 0.069 |
| Barbonymus gonionotus | 98 | BOLD:AAD1940 | 0.018 | 0.080 |
| Barbonymus mahakkamensis | 2 | BOLD:ADN2907 | 0.000 | 0.050 |
| Barbonymus schwanefeldii | 50 | BOLD:AAU0688 | 0.013 | 0.050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahruddin, H.; Sholihah, A.; Sukmono, T.; Sauri, S.; Nurhaman, U.; Wowor, D.; Steinke, D.; Hubert, N. Revisiting the Diversity of Barbonymus (Cypriniformes, Cyprinidae) in Sundaland Using DNA-Based Species Delimitation Methods. Diversity 2021, 13, 283. https://doi.org/10.3390/d13070283
Dahruddin H, Sholihah A, Sukmono T, Sauri S, Nurhaman U, Wowor D, Steinke D, Hubert N. Revisiting the Diversity of Barbonymus (Cypriniformes, Cyprinidae) in Sundaland Using DNA-Based Species Delimitation Methods. Diversity. 2021; 13(7):283. https://doi.org/10.3390/d13070283
Chicago/Turabian StyleDahruddin, Hadi, Arni Sholihah, Tedjo Sukmono, Sopian Sauri, Ujang Nurhaman, Daisy Wowor, Dirk Steinke, and Nicolas Hubert. 2021. "Revisiting the Diversity of Barbonymus (Cypriniformes, Cyprinidae) in Sundaland Using DNA-Based Species Delimitation Methods" Diversity 13, no. 7: 283. https://doi.org/10.3390/d13070283