
Observations on the Surface Structure of Aurelia solida (Scyphozoa) Polyps and Medusae

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals Collection
2.2. Histochemistry of Polyps
2.3. Sample Preparation for Scanning and Transmission Electron Microscopy
3. Results
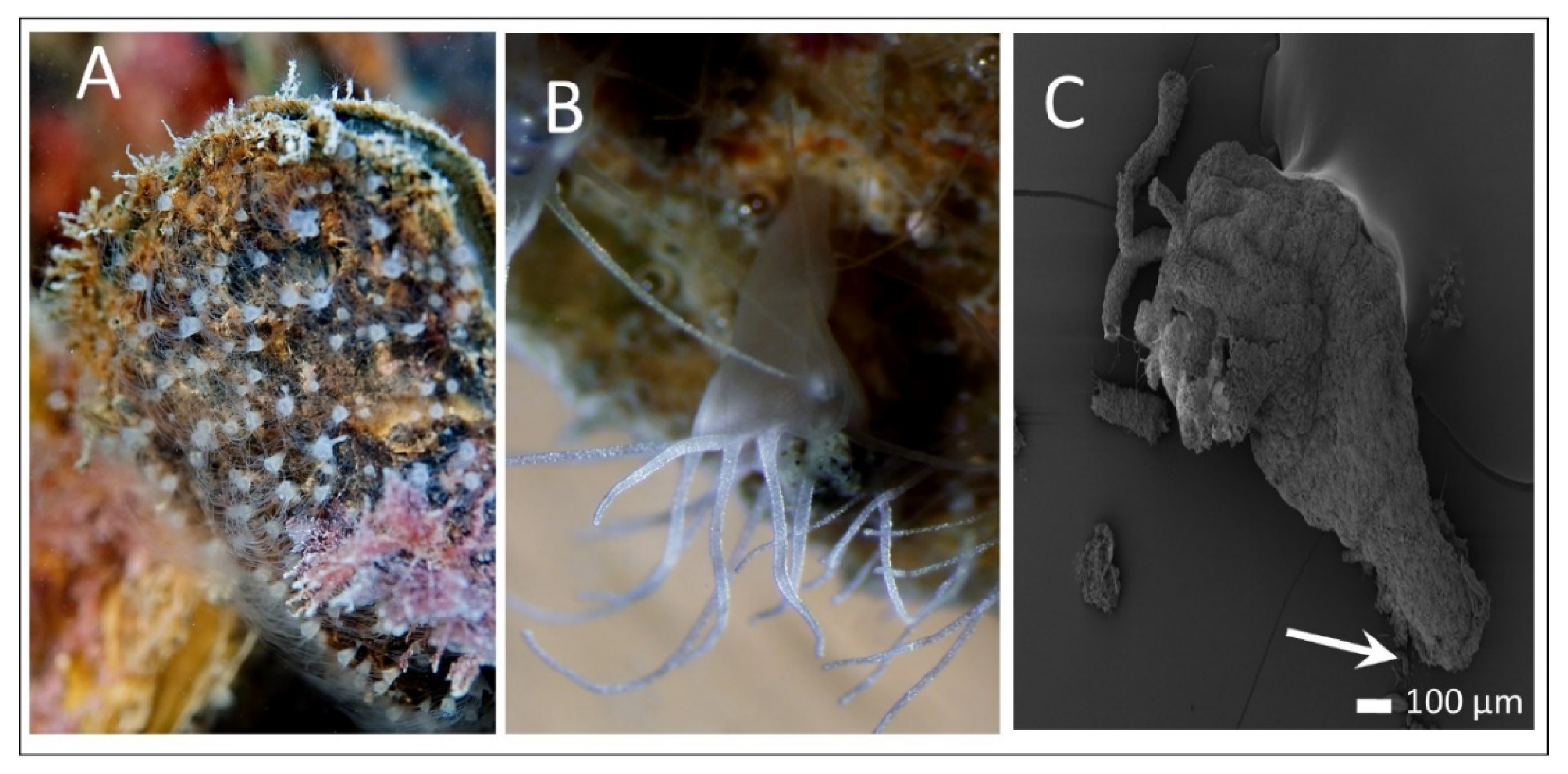
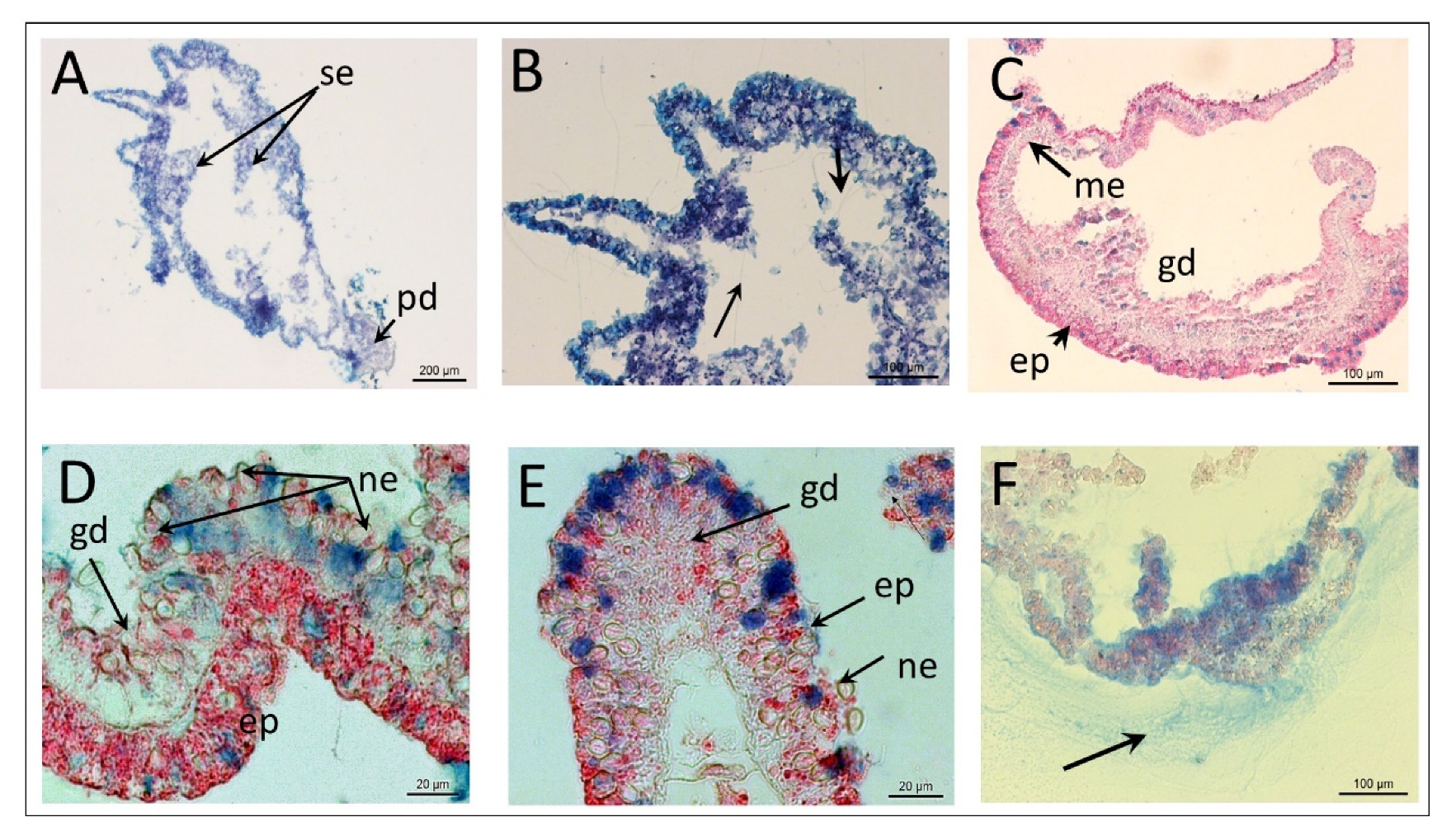
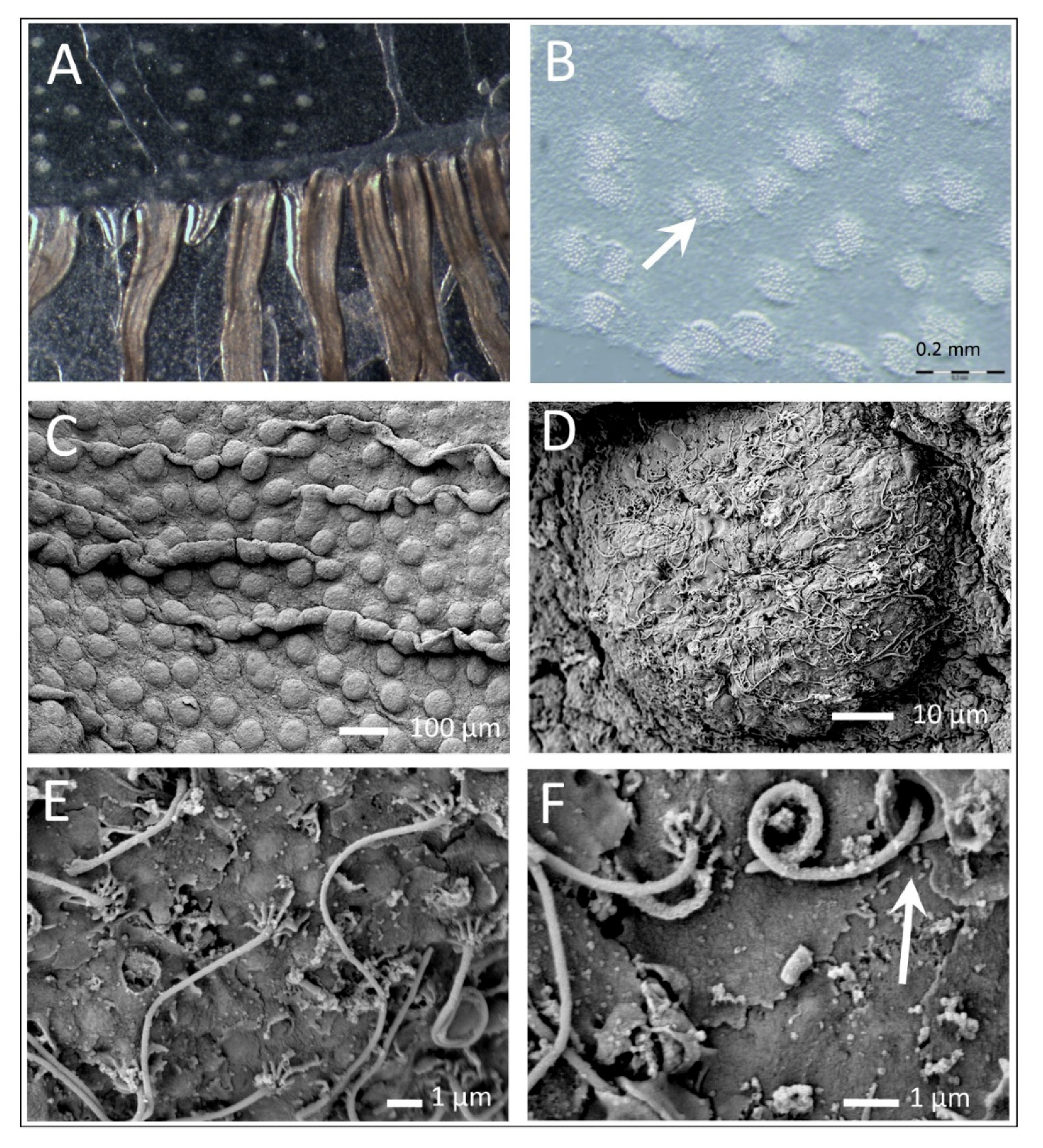
3.1. Polyp Histochemistry and Surface Morphology
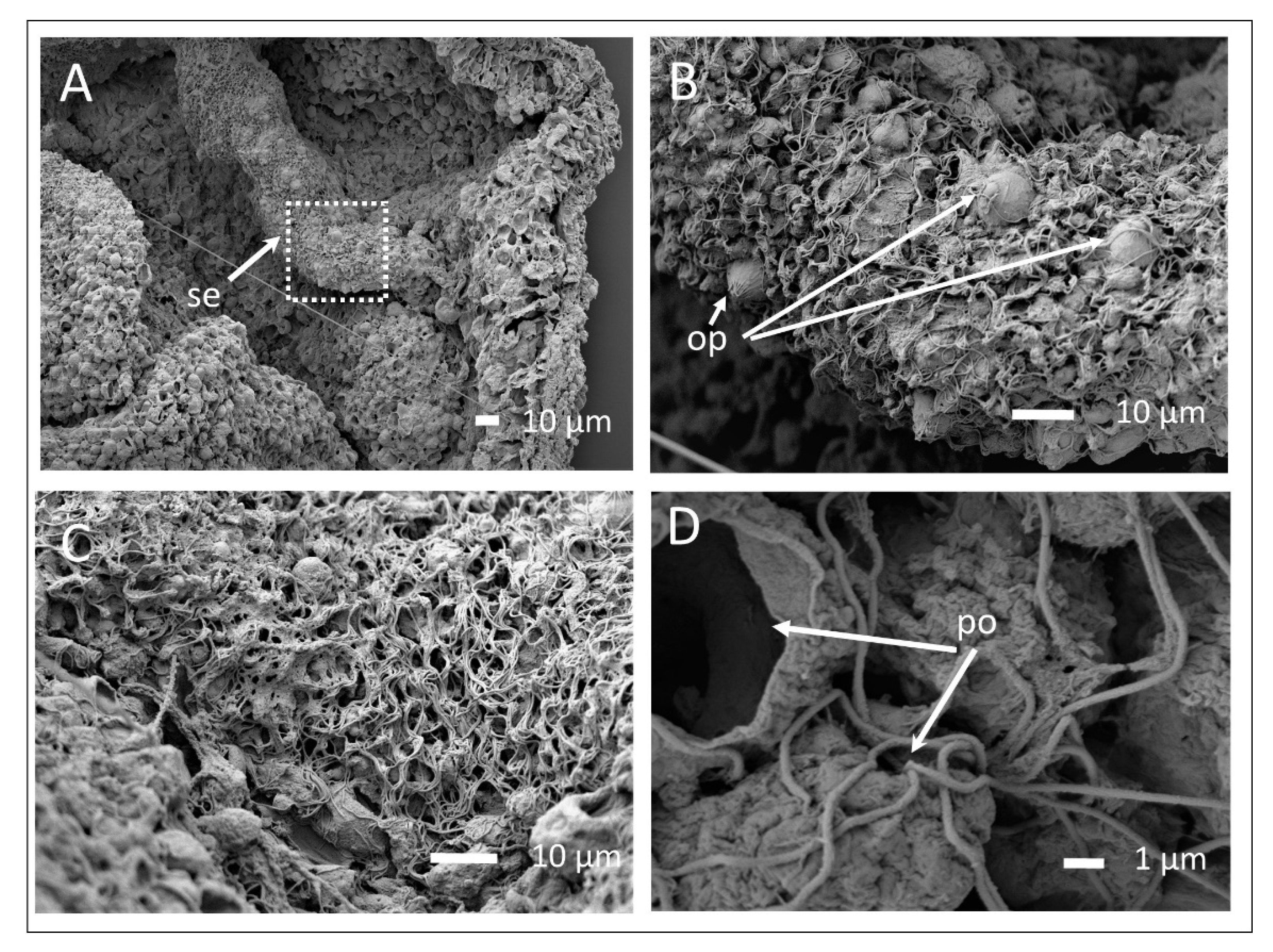
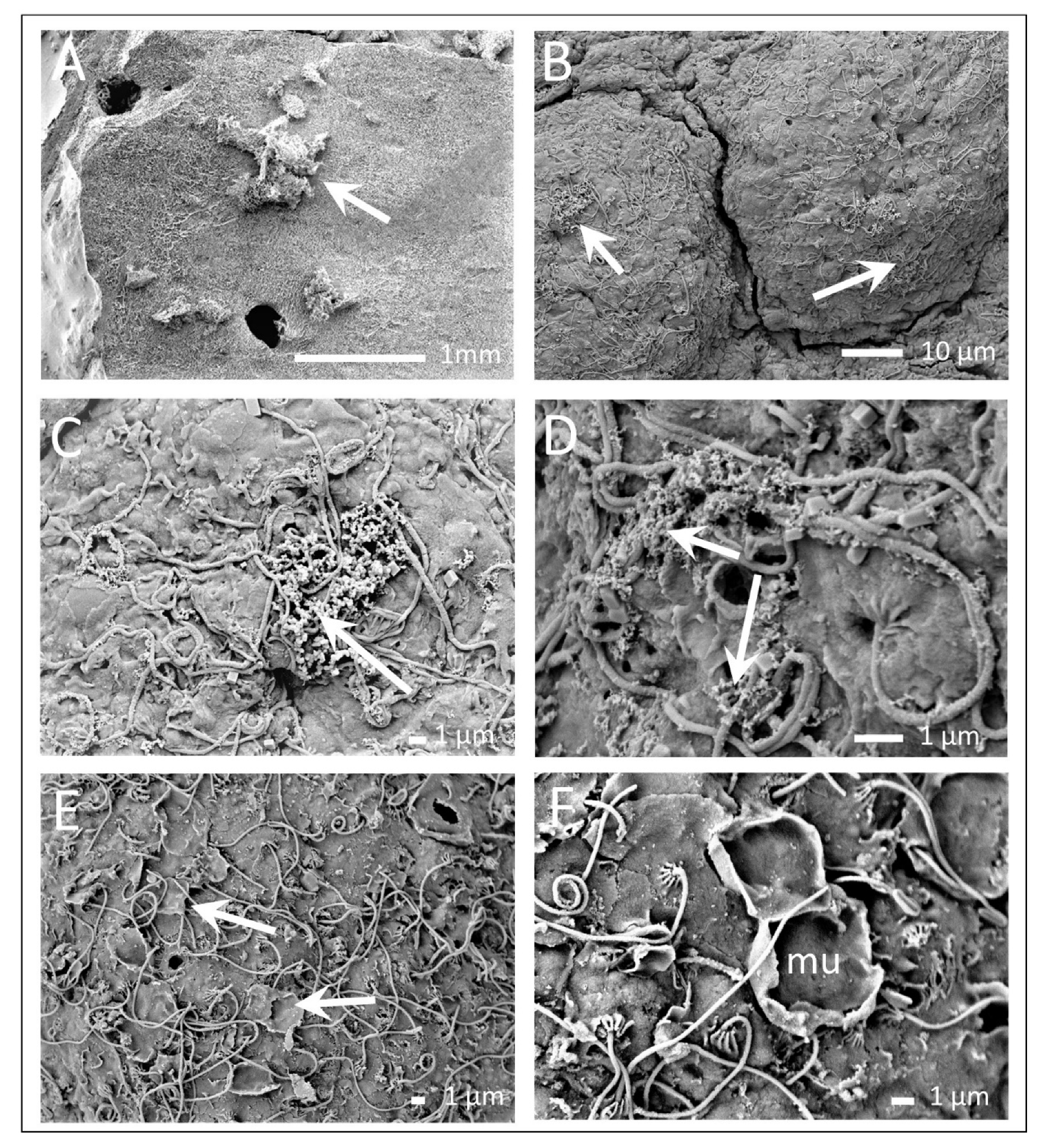
3.2. The Exumbrella Surface of the Medusa
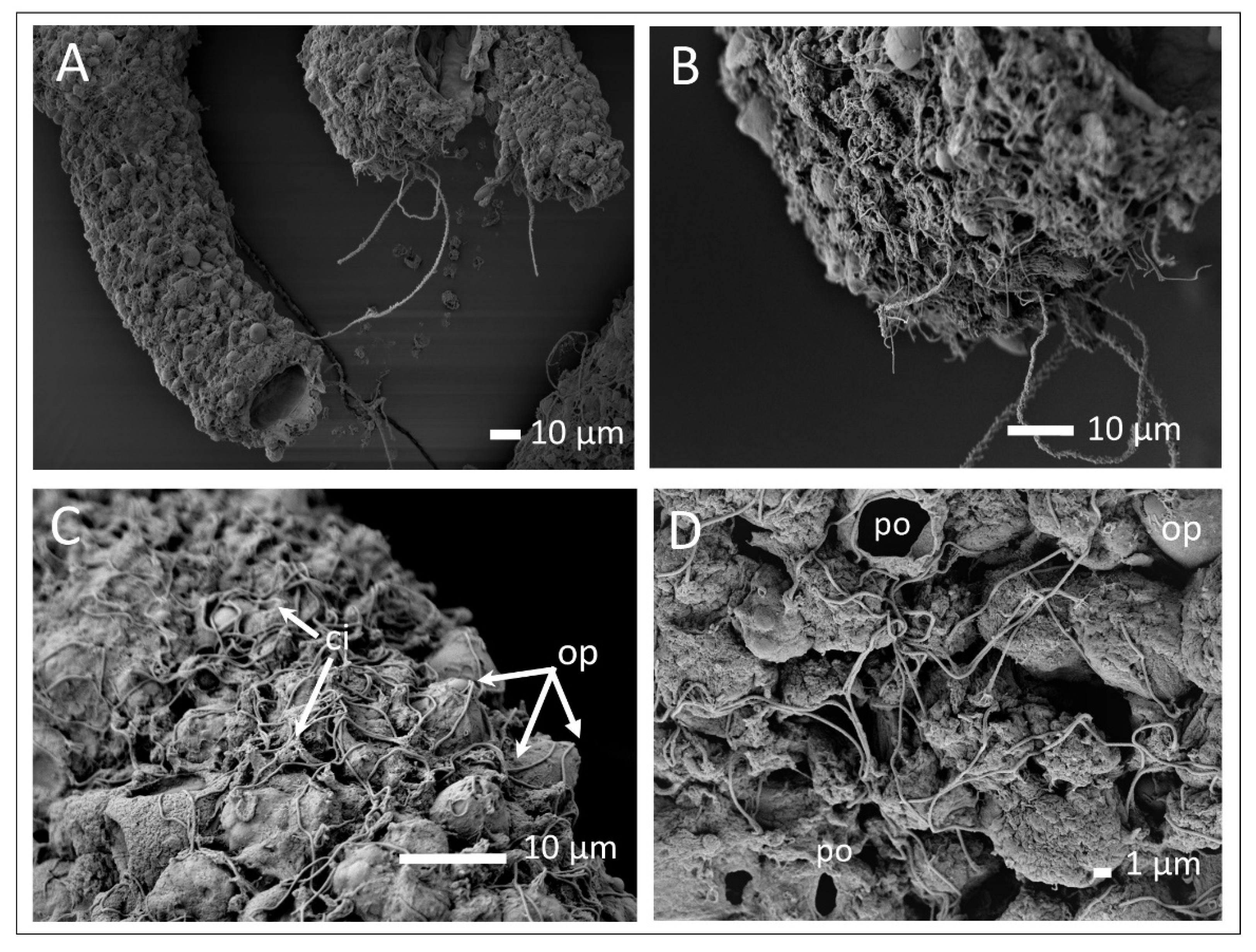
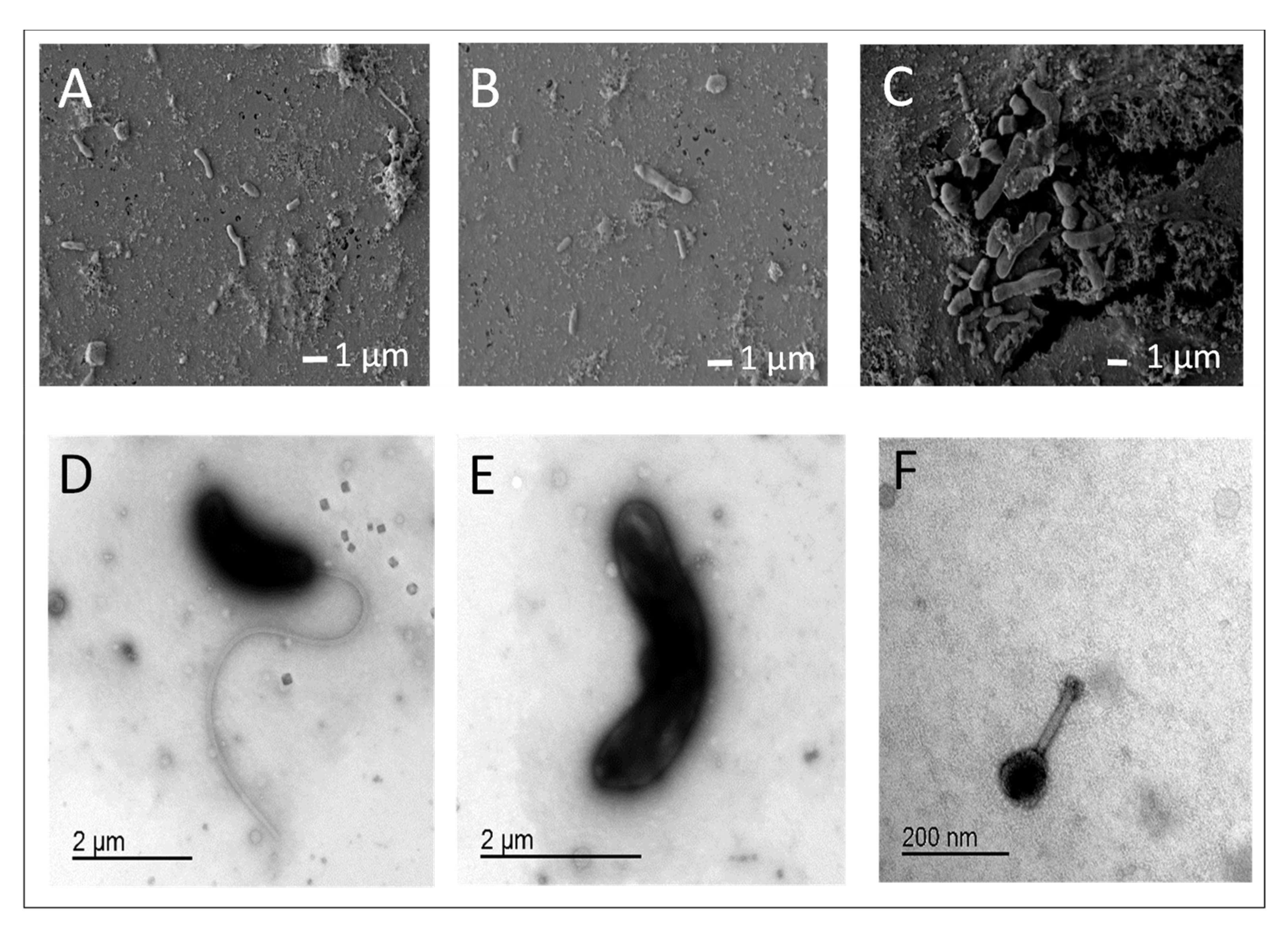
3.3. Mucus and Associated Microbes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dölger, J.; Kiørboe, T.; Andersen, A. Dense Dwarfs versus Gelatinous Giants: The Trade-Offs and Physiological Limits Determining the Body Plan of Planktonic Filter Feeders. Am. Nat. 2019, 194, E30–E40. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, S. Hidden in Plain Sight: The Ecology and Physiology of Organismal Transparency. Biol. Bull. 2001, 201, 301–318. [Google Scholar] [CrossRef] [Green Version]
- Sakai, D.; Kakiuchida, H.; Nishikawa, J.; Hirose, E. Physical Properties of the Tunic in the Pinkish-Brown Salp Pegea confoederata (Tunicata: Thaliacea). Zool. Lett. 2018, 4, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, E.; Sakai, D.; Iida, A.; Obayashi, Y.; Nishikawa, J. Exumbrellar Surface of Jellyfish: A Comparative Fine Structure Study with Remarks on Surface Reflectance. Zool. Sci. 2021, 38, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Sensui, N. Does a Nano-Scale Nipple Array (Moth-Eye Structure) Suppress the Settlement of Ascidian Larvae? J. Mar. Biol. Assoc. 2019, 99, 1393–1397. [Google Scholar] [CrossRef]
- Hirose, E.; Mayama, H.; Miyauchi, A. Does the Aquatic Invertebrate Nipple Array Prevent Bubble Adhesion? An Experiment Using Nanopillar Sheets. Biol. Lett. 2013, 9, 20130552. [Google Scholar] [CrossRef] [Green Version]
- Jarms, G.; Morandini, A.C.; Haeckel, E. World Atlas of Jellyfish: Scyphozoa except Stauromedusae, 1st ed.; Dölling und Galitz Verlag: Hamburg, Germany, 2019; p. 816. ISBN 978-3-86218-082-0. [Google Scholar]
- Collins, A.G.; Jarms, G.; Morandini, A.C. World List of Scyphozoa. Aurelia solida Browne, 1905. 2021. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=287214 (accessed on 24 April 2021).
- Pestorić, B.; Lućić, D.; Bojanić, N.; Vodopivec, M.; Kogovšek, T.; Violić, I.; Paliaga, P.; Malej, A. Scyphomedusae and Ctenophora of the Eastern Adriatic: Historical Overview and New Data. Diversity 2021, 13, 186. [Google Scholar] [CrossRef]
- Scorrano, S.; Aglieri, G.; Boero, F.; Dawson, M.N.; Piraino, S. Unmasking Aurelia Species in the Mediterranean Sea: An Integrative Morphometric and Molecular Approach. Zool. J. Linn. Soc. 2017, 180, 243–267. [Google Scholar] [CrossRef]
- Calder, D.R. Nematocysts of Polyps of Aurelia, Chrysaora, and Cyanea, and Their Utility in Identification. Trans. Am. Microsc. Soc. 1971, 90, 269–274. [Google Scholar] [CrossRef]
- Hentschel, J.; Hündgen, M. Morphologie Und Ultrastruktur Des Scyphistoma Aurelia aurita (Scyphozoa, Semaestomae). Zool. Jahrb. 1980, 1980, 295–316. [Google Scholar]
- Lauth, J.; Shepka, S.; Schurr, K. Observations on the Ultrastructure of Scyphistoma. Micron 1983, 14, 95–96. [Google Scholar] [CrossRef]
- Branden, C.; Van De Mieroop, A.; Anthoons, J.; Decleir, W. A Histochemical Study of Pigment in Strobilae and Ephyrae of Aurelia aurita. Lam. Biol. Jahrb. Dodonaea 1980, 48, 149–152. [Google Scholar]
- Chapman, G. Studies of the Mesogloea of Coelenterates. J. Cell Sci. 1953, 1953, 155–176. [Google Scholar] [CrossRef]
- Seipel, K.; Schmid, V. Mesodermal Anatomies in Cnidarian Polyps and Medusae. Int. J. Dev. Biol. 2006, 50, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaposhnikova, T.; Matveev, I.; Napara, T.; Podgornaya, O. Mesogleal Cells of the Jellyfish Are Involved in the Formation of Mesogleal Fibres. Cell. Biol. Int. 2005, 29, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, K.L.; Jamshidi, A.D.; Buckenberger, A.; Satterlie, R.A. Organization of the Subumbrellar Musculature in the Ephyra, Juvenile, and Adult Stages of Aurelia aurita Medusae. Invertebr. Biol. 2019, 138, e12260. [Google Scholar] [CrossRef]
- Leclère, L.; Röttinger, E. Diversity of Cnidarian Muscles: Function, Anatomy, Development and Regeneration. Front. Cell Dev. Biol. 2017, 4, 157. [Google Scholar] [CrossRef]
- Pallasdies, F.; Goedeke, S.; Braun, W.; Memmesheimer, R.-M. From Single Neurons to Behavior in the Jellyfish Aurelia aurita. eLife 2019, 8, e50084. [Google Scholar] [CrossRef]
- Nakanishi, N.; Hartenstein, V.; Jacobs, D.K. Development of the Rhopalial Nervous System in Aurelia sp.1 (Cnidaria, Scyphozoa). Dev. Genes Evol. 2009, 219, 301–317. [Google Scholar] [CrossRef] [Green Version]
- Chapman, D.M. Cnidarian Histology. In Coelenterate Biology. Reviews and New Perspectives; Academy Press: New York, NY, USA, 1974; pp. 1–93. [Google Scholar]
- Chapman, D.M. Microanatomy of the Bell Rim of Aurelia aurita (Cnidaria: Scyphozoa). Can. J. Zool. 1999, 77, 34–46. [Google Scholar] [CrossRef]
- Chapman, D.M. Reextension Mechanism of a Scyphistoma’s Tentacle. Can. J. Zool. 1970, 48, 931–943. [Google Scholar] [CrossRef]
- Shanks, A.; Graham, W. Chemical Defense in a Scyphomedusa. Mar. Ecol. Prog. Ser. 1988, 45, 81–86. [Google Scholar] [CrossRef]
- Brown, B.; Bythell, J. Perspectives on Mucus Secretion in Reef Corals. Mar. Ecol. Prog. Ser. 2005, 296, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Rohwer, F.; Kelley, S. Culture-Independent Analyses of Coral-Associated Microbes. In Coral Health and Disease; Rosenberg, E., Loya, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 265–277. ISBN 978-3-642-05863-9. [Google Scholar]
- Bakshani, C.R.; Morales-Garcia, A.L.; Althaus, M.; Wilcox, M.D.; Pearson, J.P.; Bythell, J.C.; Burgess, J.G. Evolutionary Conservation of the Antimicrobial Function of Mucus: A First Defence against Infection. NPJ Biofilms Microbiomes 2018, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Ducklow, H.W.; Mitchell, R. Composition of Mucus Released by Coral Reef Coelenterates 1: Coral Mucus Composition. Limnol. Oceanogr. 1979, 24, 706–714. [Google Scholar] [CrossRef]
- Liu, W.; Mo, F.; Jiang, G.; Liang, H.; Ma, C.; Li, T.; Zhang, L.; Xiong, L.; Mariottini, G.; Zhang, J.; et al. Stress-Induced Mucus Secretion and Its Composition by a Combination of Proteomics and Metabolomics of the Jellyfish Aurelia coerulea. Mar. Drugs 2018, 16, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daley, M.C.; Urban-Rich, J.; Moisander, P.H. Bacterial Associations with the Hydromedusa Nemopsis bachei and Scyphomedusa Aurelia aurita from the North Atlantic Ocean. Mar. Biol. Res. 2016, 12, 1088–1100. [Google Scholar] [CrossRef]
- Hanaoka, K.; Ohno, H.; Wada, N.; Ueno, S.; Goessler, W.; Kuehnelt, D.; Schlagenhaufen, C.; Kaise, T.; Irgolic, K.J. Occurrence of Organo-Arsenicals in Jellyfishes and Their Mucus. Chemosphere 2001, 44, 743–749. [Google Scholar] [CrossRef]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The Second Skin: Ecological Role of Epibiotic Biofilms on Marine Organisms. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the Coral Microbiome: Underpinning the Health and Resilience of Reef Ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The Sponge Holobiont in a Changing Ocean: From Microbes to Ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Kos Kramar, M.; Tinta, T.; Lučić, D.; Malej, A.; Turk, V. Bacteria Associated with Moon Jellyfish during Bloom and Post-Bloom Periods in the Gulf of Trieste (Northern Adriatic). PLoS ONE 2019, 14, e0198056. [Google Scholar] [CrossRef] [Green Version]
- Tinta, T.; Kogovšek, T.; Malej, A.; Turk, V. Jellyfish Modulate Bacterial Dynamic and Community Structure. PLoS ONE 2012, 7, e39274. [Google Scholar] [CrossRef]
- Tinta, T.; Kogovšek, T.; Klun, K.; Malej, A.; Herndl, G.J.; Turk, V. Jellyfish-Associated Microbiome in the Marine Environment: Exploring Its Biotechnological Potential. Mar. Drugs 2019, 17, 94. [Google Scholar] [CrossRef] [Green Version]
- Dinasquet, J.; Granhag, L.; Riemann, L. Stimulated Bacterioplankton Growth and Selection for Certain Bacterial Taxa in the Vicinity of the Ctenophore Mnemiopsis leidyi. Front. Microbiol. 2012, 3, 302. [Google Scholar] [CrossRef] [Green Version]
- Di Camillo, C.G.; Luna, G.M.; Bo, M.; Giordano, G.; Corinaldesi, C.; Bavestrello, G. Biodiversity of Prokaryotic Communities Associated with the Ectoderm of Ectopleura crocea (Cnidaria, Hydrozoa). PLoS ONE 2012, 7, e39926. [Google Scholar] [CrossRef] [Green Version]
- Weiland-Bräuer, N.; Neulinger, S.C.; Pinnow, N.; Künzel, S.; Baines, J.F.; Schmitz, R.A. Composition of Bacterial Communities Associated with Aurelia aurita Changes with Compartment, Life Stage, and Population. Appl. Environ. Microbiol. 2015, 81, 6038–6052. [Google Scholar] [CrossRef] [Green Version]
- Weiland-Bräuer, N.; Pinnow, N.; Langfeldt, D.; Roik, A.; Güllert, S.; Chibani, C.M.; Reusch, T.B.H.; Schmitz, R.A. The Native Microbiome Is Crucial for Offspring Generation and Fitness of Aurelia Aurita. mBio 2020, 11, e02336-20. [Google Scholar] [CrossRef] [PubMed]
- Weiland-Bräuer, N.; Prasse, D.; Brauer, A.; Jaspers, C.; Reusch, T.B.H.; Schmitz, R.A. Cultivable Microbiota Associated with Aurelia aurita and Mnemiopsis leidyi. MicrobiologyOpen 2020, 9, e1094. [Google Scholar] [CrossRef] [PubMed]
- Hočevar, S.; Malej, A.; Boldin, B.; Purcell, J. Seasonal Fluctuations in Population Dynamics of Aurelia aurita Polyps in Situ with a Modelling Perspective. Mar. Ecol. Prog. Ser. 2018, 591, 155–166. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill, L., Ed.; Elsevier: Philadelphia, PA, USA, 2008; Volume 2008, ISBN 978-0-443-10279-0. [Google Scholar]
- Pearson, R.; Tellam, R.; Xu, B.; Zhao, Z.; Willcox, M.; Kongsuwan, K. Isolation, Biochemical Characterization and Anti-Adhesion Property of Mucin from the Blue Blubber Jellyfish (Catostylus mosaicus). Biosci. Methods 2011, 2. [Google Scholar] [CrossRef]
- Parracho, T.; Morais, Z. Catostylus Tagi: Partial RDNA Sequencing and Characterisation of Nematocyte Structures Using Two Improvements in Jellyfish Sample Preparation. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 40. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.; Varki, N.M.; Jankowski, M.D.; Gagneux, P. Using Unfixed, Frozen Tissues to Study Natural Mucin Distribution. JoVE 2012, 21, e3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, D.A.; Nakanishi, N.; Hensley, N.M.; Cozzolino, K.; Tabatabaee, M.; Martin, M.; Hartenstein, V.; Jacobs, D.K. Structural and Developmental Disparity in the Tentacles of the Moon Jellyfish Aurelia sp.1. PLoS ONE 2015, 10, e0134741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeger, T.; Möller, H. Ultrastructural Observations on Prey Capture and Digestion in the Scyphomedusa Aurelia aurita. Mar. Biol. 1987, 96, 391–400. [Google Scholar] [CrossRef]
- Ruppert, E.E.; Fox, R.S.; Barnes, R.D. Invertebrate Zoology: A Functional Evolutionary Approach, 7th ed.; Thomson-Brooks/Cole: Belmont, CA, USA, 2004; ISBN 978-0-03-025982-1. [Google Scholar]
- Folino-Rorem, N.C. Phylum Cnidaria. In Thorp and Covich’s Freshwater Invertebrates; Elsevier: London, UK, 2015; pp. 159–179. ISBN 978-0-12-385026-3. [Google Scholar]
- Chia, F.-S.; Amerongen, H.M.; Peteya, D.J. Ultrastructure of the Neuromuscular System of the Polyp of Aurelia aurita L., 1758 (Cnidaria, Scyphozoa). J. Morphol. 1984, 180, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Killi, N.; Bonello, G.; Mariottini, G.L.; Pardini, P.; Pozzolini, M.; Cengiz, S. Nematocyst Types and Venom Effects of Aurelia aurita and Velella velella from the Mediterranean Sea. Toxicon 2020, 175, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Avian, M.; Malej, A. Aurelia Polyps and Medusae (Scyphozoa; Semaeostomeae; Ulmaridae) in the Northern Adriatic: Their Cnidome and Ecology. In Proceedings of the PERSEUS International Jellyfish Workshop, Cadiz, Spain, 2–3 March 2015; p. 18. [Google Scholar]
- Avian, M.; Del Negro, P.; Sandrini, L.R. A Comparative Analysis of Nematocysts in Pelagia noctiluca and Rhizostoma pulmo from the North Adriatic Sea. Hydrobiologia 1991, 216–217, 615–621. [Google Scholar] [CrossRef]
- D’Ambra, I.; Lauritano, C. A Review of Toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef]
- Hündgen, M. Cnidaria: Cell Types. In Biology of the Integument; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 47–56. ISBN 978-3-642-51595-8. [Google Scholar]
- Patwa, A.; Thiéry, A.; Lombard, F.; Lilley, M.K.S.; Boisset, C.; Bramard, J.-F.; Bottero, J.-Y.; Barthélémy, P. Accumulation of Nanoparticles in “Jellyfish” Mucus: A Bio-Inspired Route to Decontamination of Nano-Waste. Sci. Rep. 2015, 5, 11387. [Google Scholar] [CrossRef] [Green Version]
- Garren, M.; Azam, F. Corals Shed Bacteria as a Potential Mechanism of Resilience to Organic Matter Enrichment. ISME J. 2012, 6, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, O.H.; Fernandez, V.I.; Garren, M.; Guasto, J.S.; Debaillon-Vesque, F.P.; Kramarsky-Winter, E.; Vardi, A.; Stocker, R. Vortical Ciliary Flows Actively Enhance Mass Transport in Reef Corals. Proc. Natl. Acad. Sci. USA 2014, 111, 13391–13396. [Google Scholar] [CrossRef] [Green Version]
- Jordano, M. de A.; Morandini, A.C.; Nagata, R.M. Is Phenotypic Plasticity Determined by Temperature and Fluid Regime in Filter-Feeding Gelatinous Organisms? J. Exp. Mar. Biol. Ecol. 2020, 522, 151238. [Google Scholar] [CrossRef]
- Johnston, I.S.; Rohwer, F. Microbial Landscapes on the Outer Tissue Surfaces of the Reef-Building Coral Porites compressa. Coral Reefs 2007, 26, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, K. Regulation of Microbial Populations by Coral Surface Mucus and Mucus-Associated Bacteria. Mar. Ecol. Prog. Ser. 2006, 322, 1–14. [Google Scholar] [CrossRef]
- Daniels, C.; Zeifman, A.; Heym, K.; Ritchie, K.; Watson, C.; Berzins, I.; Breitbart, M. Spatial Heterogeneity of Bacterial Communities in the Mucus of Montastraea annularis. Mar. Ecol. Prog. Ser. 2011, 426, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Daniels, C.; Breitbart, M. Bacterial Communities Associated with the Ctenophores Mnemiopsis leidyi and Beroe ovata. FEMS Microbiol. Ecol. 2012, 82, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, K.B.; Smith, G.W. Microbial Communities of Coral Surface Mucopolysaccharide Layers. In Coral Health and Disease; Rosenberg, E., Loya, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 259–264. ISBN 978-3-642-05863-9. [Google Scholar]
- Teplitski, M.; Krediet, C.J.; Meyer, J.L.; Ritchie, K.B. Microbial Interactions on Coral Surfaces and Within the Coral Holobiont. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 331–346. ISBN 978-3-319-31303-0. [Google Scholar]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, F. Oceanography: Microbes, Molecules, and Marine Ecosystems. Science 2004, 303, 1622–1624. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turk, V.; Fortič, A.; Kos Kramar, M.; Tušek Žnidarič, M.; Štrus, J.; Kostanjšek, R.; Malej, A. Observations on the Surface Structure of Aurelia solida (Scyphozoa) Polyps and Medusae. Diversity 2021, 13, 244. https://doi.org/10.3390/d13060244
Turk V, Fortič A, Kos Kramar M, Tušek Žnidarič M, Štrus J, Kostanjšek R, Malej A. Observations on the Surface Structure of Aurelia solida (Scyphozoa) Polyps and Medusae. Diversity. 2021; 13(6):244. https://doi.org/10.3390/d13060244
Chicago/Turabian StyleTurk, Valentina, Ana Fortič, Maja Kos Kramar, Magda Tušek Žnidarič, Jasna Štrus, Rok Kostanjšek, and Alenka Malej. 2021. "Observations on the Surface Structure of Aurelia solida (Scyphozoa) Polyps and Medusae" Diversity 13, no. 6: 244. https://doi.org/10.3390/d13060244