
Assessing the Genetic Diversity of Ilex guayusa Loes., a Medicinal Plant from the Ecuadorian Amazon

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
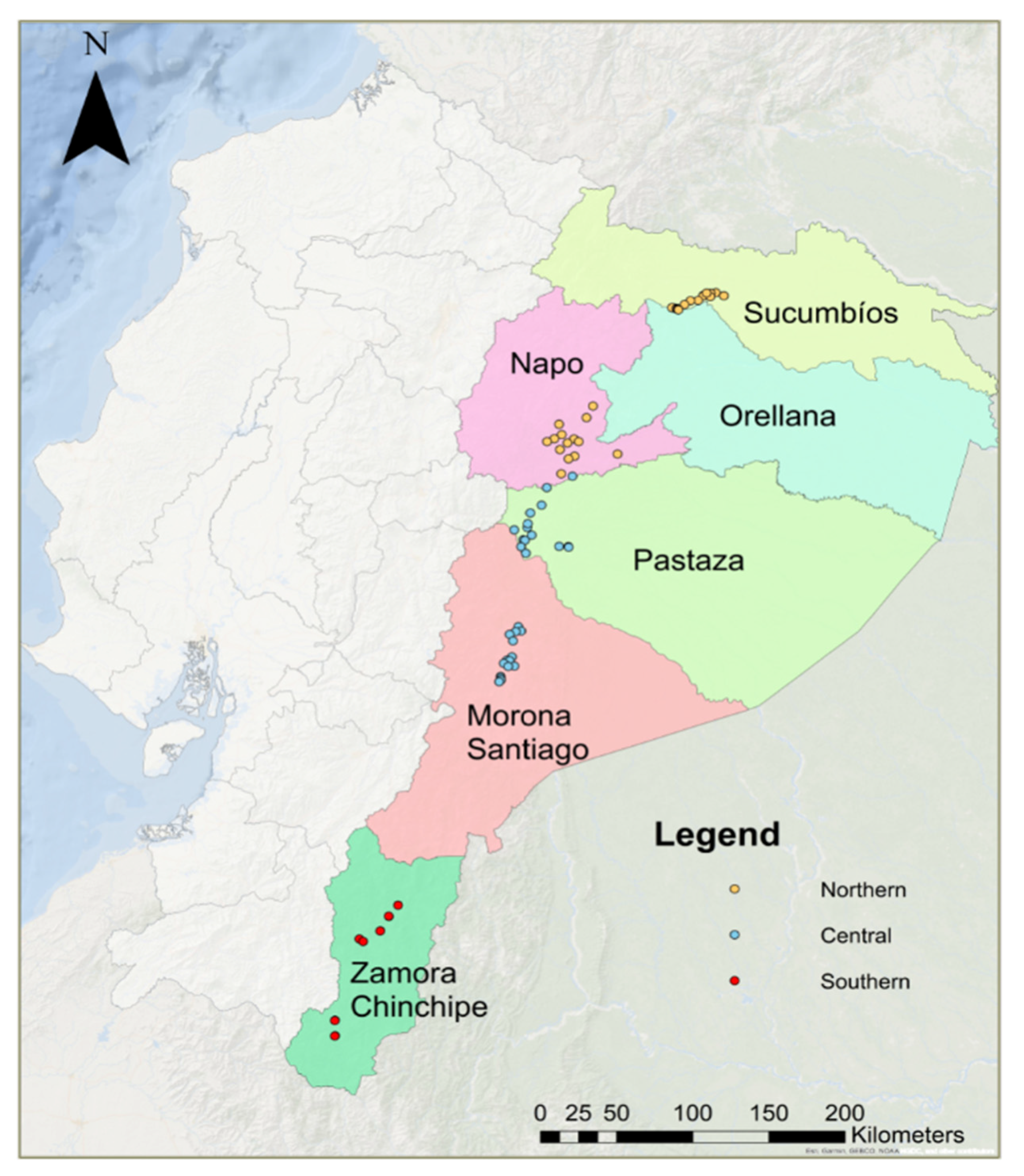
2.1. Sample Collection
2.2. Genomic DNA Isolation
2.3. Development of SSR Markers for Ilex guayusa
2.4. Sample Preparation and Genotyping
2.5. Data Analysis
2.5.1. Genetic Diversity
2.5.2. Population Structure
2.5.3. Testing for Isolation-by-Distance (IBD) and Isolation-by-Environment (IBE)
2.5.4. Niche Characterization Modeling
3. Results
3.1. Clonal and Genetic Diversity of Ilex guayusa in the Ecuadorian Amazon
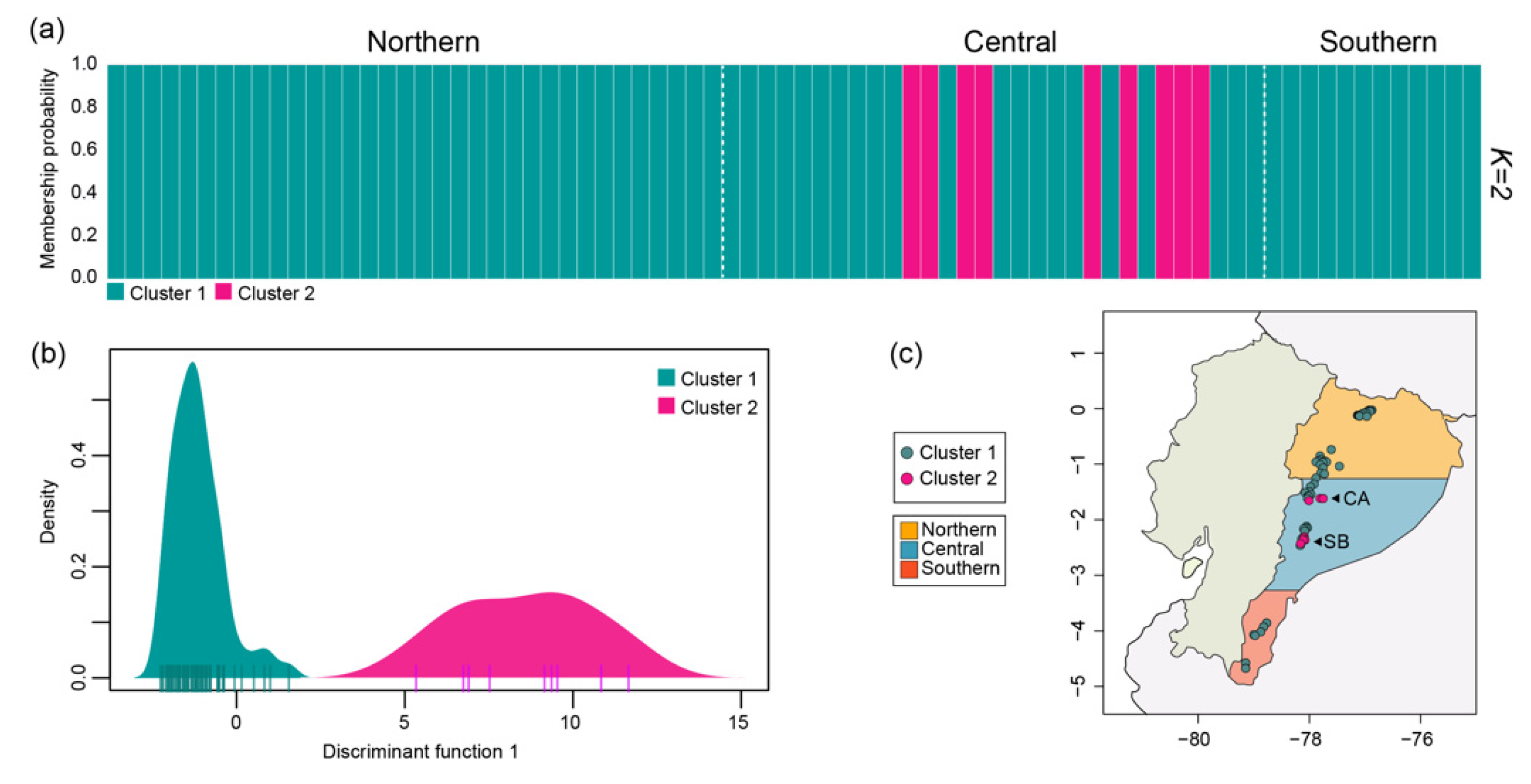
3.2. Population Structure
3.3. Isolation-by-Distance (IBD) and Isolation-by-Environment (IBE)
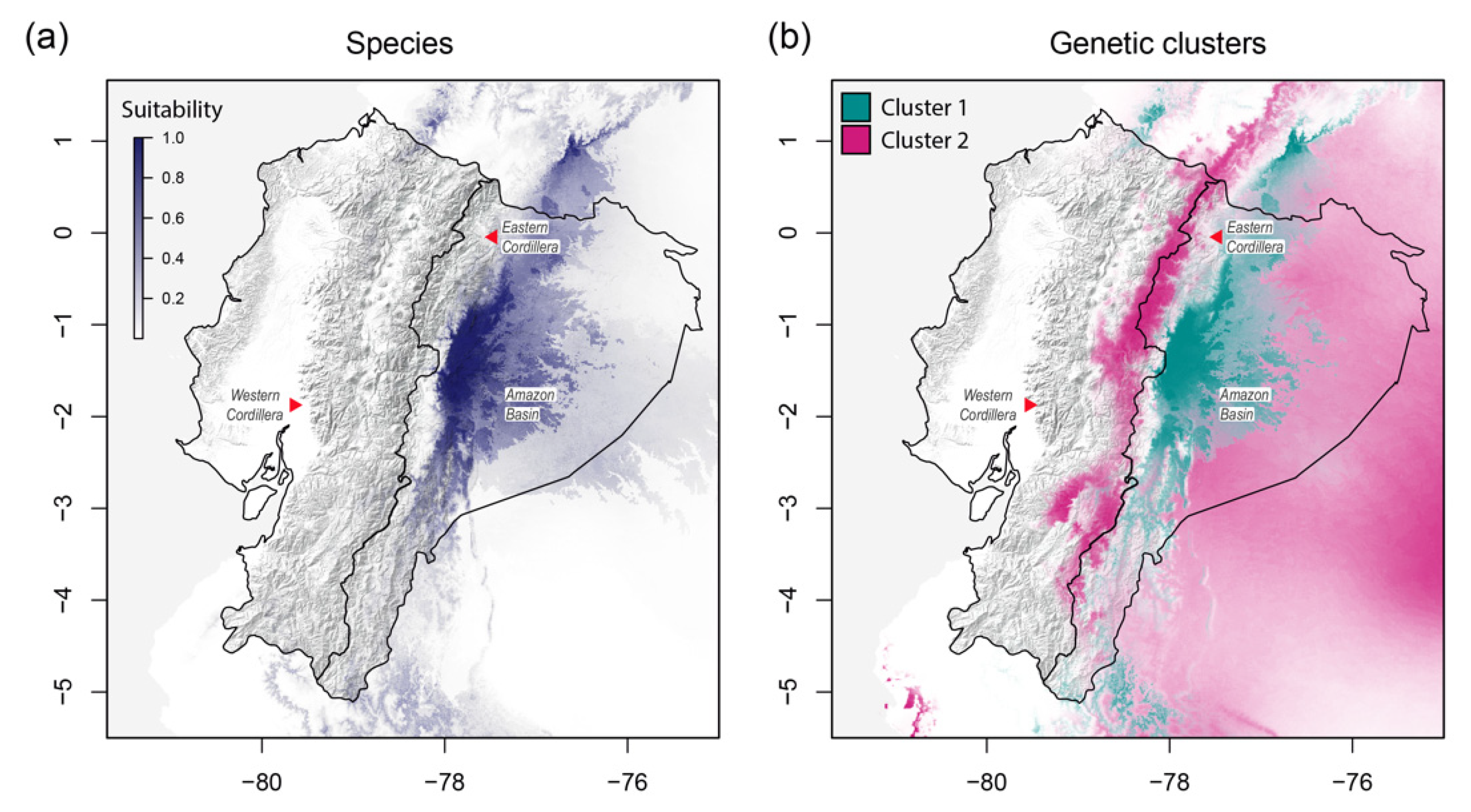
3.4. Niche Characterization Modeling
4. Discussion
4.1. A Moderately Low Genetic Diversity for Ilex guayusa in the Ecuadorian Amazon
4.2. High Clonal Diversity for Ilex guayusa in the Ecuadorian Amazon
4.3. Influence of the Environment and Human Activities as Modulators of Ilex guayusa Population Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, W.H.; Kennelly, E.J.; Bass, G.N.; Wedner, H.J.; Elvin-Lewis, M.P.; Fast, D. Ritualistic use of the holly Ilex guayusa by Amazonian Jivaro Indians. J. Ethnopharmacol. 1991, 33, 25–30. [Google Scholar] [CrossRef]
- Sequeda-Castañeda, L.G.; Modesti Costa, G.; Celis, C.; Gamboa, F.; Gutiérrez, S.; Luengas, P. Ilex guayusa Loes (Aquifoliaceae): Amazon and Andean native plant. Pharmacol. OnLine 2016, 3, 193–202. [Google Scholar]
- Dueñas, J.F.; Jarrett, C.; Cummins, I.; Logan-Hines, E. Amazonian Guayusa (Ilex guayusa Loes.): A Historical and Ethnobotanical Overview. Econ. Bot. 2016, 70, 85–91. [Google Scholar] [CrossRef]
- Schultes, R.E. Ilex guayusa from 500 A.D. to the present. In Etnologiska Studier; Göteborgs Etnografiska Museum: Göteborg, Sweden, 1972; Volume 32, pp. 115–138. [Google Scholar]
- Wassen, H. A Medicine-man’s Implements and Plants in a Tiahuanacoid Tomb in Highland Bolivia. In Etnologiska Studier; Göteborgs Etnografiska Museum: Göteborg, Sweden, 1972; Volume 32, pp. 8–114. [Google Scholar]
- Kapp, R.W.; Mendes, O.; Roy, S.; McQuate, R.S.; Kraska, R. General and Genetic Toxicology of Guayusa Concentrate (Ilex guayusa). Int. J. Toxicol. 2016, 35, 222–242. [Google Scholar] [CrossRef] [Green Version]
- Schultes, R.E. Discovery of an ancient guayusa plantation in Colombia. Bot. Mus. Leafl. Harv. Univ. 1979, 27, 143–153. [Google Scholar] [CrossRef]
- García-Ruiz, A.; Baenas, N.; Benítez-González, A.M.; Stinco, C.M.; Meléndez-Martínez, A.J.; Moreno, D.A.; Ruales, J. Guayusa (Ilex guayusa L.) new tea: Phenolic and carotenoid composition and antioxidant capacity. J. Sci. Food Agric. 2017, 97, 3929–3936. [Google Scholar] [CrossRef]
- Radice, M.; Cossio, N.; Scalvenzi, L. Ilex guayusa: A systematic review of its Traditional Uses, Chemical Constituents, Biological Activities and Biotrade Opportunities. In MOL2NET: FROM MOLECULES TO NETWORKS, Proceedings of the MOL2NET 2016, International Conference on Multidisciplinary Sciences, Basel, Switzerland, 15 January–15 December 2016, 2nd ed.; MDPI: Basel, Switzerland, 2017; pp. 1–7. [Google Scholar]
- Patino, V.M. Guayusa, a neglected stimulant from the eastern Andean foothills. Econ. Bot. 1968, 22, 311–316. [Google Scholar] [CrossRef]
- Wise, G.; Santander, D.E. Assessing the History of Safe Use of Guayusa. J. Food Nutr. Res. 2018, 6, 471–475. [Google Scholar] [CrossRef]
- Shemluck, M. The Flowers of Ilex guayusa. Bot. Mus. Leafl. Harv. Univ. 1979, 27, 155–160. [Google Scholar]
- GBIF Backbone Taxonomy. Ilex guayusa Loes. in GBIF Secretariat. Available online: https://www.gbif.org/species/5534620 (accessed on 24 June 2020).
- Cascales, J.; Bracco, M.; Poggio, L.; Gottlieb, A.M. Genetic diversity of wild germplasm of “yerba mate” (Ilex paraguariensis St. Hil.) from Uruguay. Genetica 2014, 142, 563–573. [Google Scholar] [CrossRef]
- Pereira, M.F.; Ciampi, A.Y.; Inglis, P.W.; Souza, V.A.; Azevedo, V.C.R. Shotgun Sequencing for Microsatellite Identification in Ilex paraguariensis (Aquifoliaceae). Appl. Plant Sci. 2013, 1, 1–3. [Google Scholar] [CrossRef]
- Gauer, L.; Cavalli-Molina, S. Genetic variation in natural populations of maté (Ilex paraguariensis A. St.-Hil., Aquifoliaceae) using RAPD markers. Heredity 2000, 84, 647–656. [Google Scholar] [CrossRef]
- Chen, W.-W.; Xiao, Z.-Z.; Tong, X.; Liu, Y.-P.; Li, Y.-Y. Development and characterization of 25 microsatellite primers for Ilex chinensis (Aquifoliaceae). Appl. Plant Sci. 2015, 3, 1500057. [Google Scholar] [CrossRef] [PubMed]
- Torimaru, T.; Tomaru, N.; Nishimura, N.; Yamamoto, S. Clonal diversity and genetic differentiation in Ilex leucoclada M. patches in an old-growth beech forest. Mol. Ecol. 2003, 12, 809–818. [Google Scholar] [CrossRef]
- Rendell, S.; Ennos, R.A. Chloroplast DNA diversity of the dioecious European tree Ilex aquifolium L. (English holly). Mol. Ecol. 2003, 12, 2681–2688. [Google Scholar] [CrossRef]
- Zhang, J.H.; Gao, Y.Z.; Zhang, B.; Wang, Z.J. Genetic diversity of Ilex L. tree species. Acta Bot. Boreali Occident. Sin. 2011, 31, 504–510. [Google Scholar]
- Xi-Jun, Z.; Dong-Mei, Z.; Yu-Lan, L.; Jin-Le, S.; Jian-Xiang, L. Inter-simple sequence repeats (ISSR) marker analysis of flex plants species and its application. J. Henan Agric. Univ. 2009, 43, 196–200. [Google Scholar]
- Qian, Y.S.; Wang, H.Z.; Shi, N.N.; Zhao, Y.; Li, N.L.; Hu, Z. Studies of genetic diversity among 10 species of Ilex based on RAPD and AFLPs. J. Mol. Cell Biol. 2008, 41, 35–43. [Google Scholar]
- Heck, C.; de Mejia, E. Yerba Mate Tea (Ilex paraguariensis): A Comprehensive Review on Chemistry, Health Implications, and Technological Considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar] [CrossRef]
- Kothiyal, S.K.; Sati, S.C.; Rawat, M.S.M.; Sati, M.D.; Semwal, D.K.; Semwal, R.B.; Sharma, A.; Rawat, B.; Kumar, A. Chemical Constituents and Biological Significance of the Genus Ilex (Aquifoliaceae). Nat. Prod. J. 2012, 2, 212–224. [Google Scholar] [CrossRef]
- Salvador, A.T.; Mosquera, J.; Jaramillo, V.; Arahana, V.; Torres, M.D.L. Preliminary assessment of the degree of genetic diversity of Ecuadorean Ilex guayusa using inter simple sequence repeat (ISSR) markers. ACI Av. Cienc. Ing. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Brzosko, E.; Wróblewska, A.; Ratkiewicz, M. Spatial genetic structure and clonal diversity of island populations of lady’s slipper (Cypripedium calceolus) from the Biebrza National Park (northeast Poland). Mol. Ecol. 2002, 11, 2499–2509. [Google Scholar] [CrossRef]
- Pleasants, J.M.; Wendel, J.F. Genetic Diversity in a Clonal Narrow Endemic, Erythronium propullans, and in Its Widespread Progenitor, Erythronium albidum. Am. J. Bot. 1989, 76, 1136–1151. [Google Scholar] [CrossRef]
- McKey, D.; Elias, M.; Pujol, B.; Duputié, A. The evolutionary ecology of clonally propagated domesticated plants. N. Phytol. 2010, 186, 318–332. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute. ArcGIS Desktop: Release 10. Available online: https://desktop.arcgis.com/es/ (accessed on 18 June 2020).
- Križman, M.; Jakše, J.; Baričevič, D.; Javornik, B.; Prošek, M. Robust CTAB-activated charcoal protocol for plant DNA extraction. Acta Agric. Slov. 2006, 87, 427–433. [Google Scholar]
- Rezadoost, M.H.; Kordrostami, M.; Kumleh, H.H. An efficient protocol for isolation of inhibitor-free nucleic acids even from recalcitrant plants. 3 Biotech 2016, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, S.M.; Fox, G.; Briggs, P.J.; Donaldson, I.J.; Hood, S.; Richardson, P.; Leaver, G.W.; Truelove, N.K.; Preziosi, R.F. A Galaxy-based bioinformatics pipeline for optimised, streamlined microsatellite development from Illumina next-generation sequencing data. Conserv. Genet. Resour. 2016, 8, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data 2010; Babraham Institute: Cambridge, UK, 2014; Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 10 January 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoe, T.A.; Poole, A.W.; de Koning, A.P.J.; Jones, K.L.; Tomback, D.F.; Oyler-McCance, S.J.; Fike, J.A.; Lance, S.L.; Streicher, J.W.; Smith, E.N.; et al. Rapid Microsatellite Identification from Illumina Paired-End Genomic Sequencing in Two Birds and a Snake. PLoS ONE 2012, 7, e30953. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Blacket, M.J.; Robin, C.; Good, R.T.; Lee, S.F.; Miller, A.D. Universal primers for fluorescent labelling of PCR fragments—An efficient and cost-effective approach to genotyping by fluorescence. Mol. Ecol. Resour. 2012, 12, 456–463. [Google Scholar] [CrossRef]
- Hulce, D.; Li, X.; Snyder-Leiby, T.; Liu, C.J. GeneMarker® Genotyping Software: Tools to Increase the Statistical Power of DNA Fragment Analysis. J. Biomol. Tech. 2011, 22, S35–S36. [Google Scholar]
- Meirmans, P.G.; van Tienderen, P.H. Genotype and genodive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Bruvo, R.; Michiels, N.K.; D’Souza, T.G.; Schulenburg, H. A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 2004, 13, 2101–2106. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 29 February 2020).
- Pluess, A.R.; Stöcklin, J. Population genetic diversity of the clonal plant Geum reptans (Rosaceae) in the Swiss Alps. Am. J. Bot. 2004, 91, 2013–2021. [Google Scholar] [CrossRef]
- Parks, J.C.; Werth, C.R. A Study of Spatial Features of Clones in a Population of Bracken Fern, Pteridium aquilinum (Dennstaedtiaceae). Am. J. Bot. 1993, 80, 537. [Google Scholar] [CrossRef]
- Goudet, J. hierfstat, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Bateman, A. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.V.; Jasieniuk, M. polysat: An R package for polyploid microsatellite analysis. Mol. Ecol. Resour. 2011, 11, 562–566. [Google Scholar] [CrossRef]
- Chapuis, M.-P.; Estoup, A. Microsatellite Null Alleles and Estimation of Population Differentiation. Mol. Biol. Evol. 2006, 24, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, E. pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, N.J.; Everhart, S.E.; Knaus, B.J.; Kamvar, Z.N. Best Practices for Population Genetic Analyses. Phytopathology 2017, 107, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Beugin, M.; Gayet, T.; Pontier, D.; Devillard, S.; Jombart, T. A fast likelihood solution to the genetic clustering problem. Methods Ecol. Evol. 2018, 9, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. vegan: Community Ecology Package; Version 2.5–6. Available online: https://CRAN.R-project.org/package=vegan (accessed on 19 September 2020).
- Kindt, R.; Coe, R. A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies. In Tree Diversity Analysis, 1st ed.; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005; ISBN 92-9059-179-X. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R. raster: Geographic Data Analysis and Modeling; Version 3.1–5. Available online: https://CRAN.R-project.org/package=raster (accessed on 19 September 2020).
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Luo, M.-X.; Gao, R.-H.; Zhang, W.; Yang, Y.-Z.; Li, Y.-J.; Liao, P.-C. Isolation-by-environment as a driver of genetic differentiation among populations of the only broad-leaved evergreen shrub Ammopiptanthus mongolicus in Asian temperate deserts. Sci. Rep. 2019, 9, 12008–12014. [Google Scholar] [CrossRef]
- Hijmans, R. geosphere: Spherical Trigonometry; Version 1.5–10. Available online: https://CRAN.R-project.org/package=geosphere (accessed on 19 September 2020).
- Venables, W.; Ripley, B. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-21706-2. [Google Scholar]
- Phillips, S.; Dudik, M.; Schapire, R. Maxent Software for Modeling Species Niches and Distributions; Version 3.4.1; American Museum of Natural History: New York, NY, USA, 2021; Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 25 June 2020).
- University of California, Davis. DAV—UC Davis Herbarium; Center for Plant Diversity: Davis, CA, USA, 2020; Available online: https://www.gbif.org/dataset/c267909d-184f-4138-9e67-a074d478d52b (accessed on 24 June 2020).
- Ramirez, J.; Tulig, M.; Watson, K.; Thiers, B. The New York Botanical Garden Herbarium (NY); Version 1.22; The New York Botanical Garden: New York, NY, USA, 2020; Available online: https://www.gbif.org/dataset/d415c253-4d61-4459-9d25-4015b9084fb0 (accessed on 24 June 2020).
- Institute of Botany, University of Hohenheim. Visual Plants (144.41.33.158)—Private Collection of Rainer Bussmann. Available online: https://www.gbif.org/dataset/85cf474e-f762-11e1-a439-00145eb45e9a (accessed on 24 June 2020).
- Crop Wild Relatives Occurrence data consortia. A Global Database for the Distributions of Crop Wild Relatives; Version 1.12; Centro Internacional de Agricultura Tropical—CIAT: Cali, Colombia, 2018; Available online: https://www.gbif.org/dataset/07044577-bd82-4089-9f3a-f4a9d2170b2e (accessed on 24 June 2020).
- Ueda, K. iNaturalist Research-grade Observations; iNaturalist.org. Available online: https://www.gbif.org/dataset/50c9509d-22c7-4a22-a47d-8c48425ef4a7 (accessed on 24 June 2020).
- Orrell, T. NMNH Extant Specimen Records; Version 1.32; National Museum of Natural History, Smithsonian Institution: Washington, DC, USA, 2020; Available online: https://www.gbif.org/dataset/821cc27a-e3bb-4bc5-ac34-89ada245069d (accessed on 24 June 2020).
- Herbarium of the University of Aarhus. The AAU Herbarium Database. Available online: https://www.gbif.org/dataset/833db434-f762-11e1-a439-00145eb45e9a (accessed on 24 June 2020).
- Grant, S.; Niezgoda, C. Field Museum of Natural History (Botany) Seed Plant Collection; Version 11.12; Field Museum: Chicago, IL, USA, 2020; Available online: https://www.gbif.org/dataset/90c853e6-56bd-480b-8e8f-6285c3f8d42b (accessed on 24 June 2020).
- Magill, B.; Solomon, J.; Stimmel, H. Tropicos Specimen Data; Missouri Botanical Garden: St. Louis, MO, USA, 2020; Available online: https://www.gbif.org/dataset/7bd65a7a-f762-11e1-a439-00145eb45e9a (accessed on 24 June 2020).
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Jayasinghe, S.L.; Kumar, L. Modeling the climate suitability of tea (Camellia sinensis (L.) O. Kuntze) in Sri Lanka in response to current and future climate change scenarios. Agric. For. Meteorol. 2019, 272–273, 102–117. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Balloux, F.; Lugon-Moulin, N. The estimation of population differentiation with microsatellite markers. Mol. Ecol. 2002, 11, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C.D.F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2010, 177, 309–334. [Google Scholar] [CrossRef]
- Forneck, A. Plant Breeding: Clonality—A Concept for Stability and Variability During Vegetative Propagation. In Progress in Botany; Esser, K., Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 66, pp. 164–183. [Google Scholar]
- Arnaud-Haond, S.; Alberto, F.; Teixeira, S.; Procaccini, G.; Serrão, E.A.; Duarte, C.M. Assessing Genetic Diversity in Clonal Organisms: Low Diversity or Low Resolution? Combining Power and Cost Efficiency in Selecting Markers. J. Hered. 2005, 96, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.A.; Rafii, M.Y.; Mahmud, T.M.M.; Hanafi, M.M.; Miah, G. Genetic Diversity of Torch Ginger (Etlingera elatior) Germplasm Revealed by ISSR and SSR Markers. BioMed Res. Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Serra, I.; Procaccini, G.; Intrieri, M.; Migliaccio, M.; Mazzuca, S.; Innocenti, A. Comparison of ISSR and SSR markers for analysis of genetic diversity in the seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 2007, 338, 71–79. [Google Scholar] [CrossRef]
- Júnior, E.L.C.; Donaduzzi, C.M.; Ferrarese-Filho, O.; Friedrich, J.C.; Gonela, A.; Sturion, J.A. Quantitative genetic analysis of methylxanthines and phenolic compounds in mate progenies. Pesqui. Agropecuária Bras. 2010, 45, 171–177. [Google Scholar] [CrossRef]
- Tarragó, J.; Sansberro, P.; Filip, R.; López, P.; González, A.; Luna, C.; Mroginski, L. Effect of leaf retention and flavonoids on rooting of Ilex paraguariensis cuttings. Sci. Hortic. 2005, 103, 479–488. [Google Scholar] [CrossRef]
- Balloux, F.; Lehmann, L.; de Meeûs, T. The population genetics of clonal and partially clonal diploids. Genetics 2003, 164, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.K.; Kollmann, J. Low genetic diversity in small peripheral populations of a rare European tree (Sorbus torminalis) dominated by clonal reproduction. Conserv. Genet. 2008, 9, 1533–1539. [Google Scholar] [CrossRef]
- Halkett, F.; Simon, J.-C.; Balloux, F. Tackling the population genetics of clonal and partially clonal organisms. Trends Ecol. Evol. 2005, 20, 194–201. [Google Scholar] [CrossRef]
- Elias, M.; Penet, L.; Vindry, P.; McKey, D.; Panaud, O.; Robert, T. Unmanaged sexual reproduction and the dynamics of genetic diversity of a vegetatively propagated crop plant, cassava (Manihot esculenta Crantz), in a traditional farming system. Mol. Ecol. 2001, 10, 1895–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, F.M.D.R.; Lenzi, M.; Ferreira, B.H.D.S.; da Silva, L.V.; Zanella, C.M.; Paggi, G.M. High genetic diversity and moderate genetic structure in the self-incompatible, clonal Bromelia hieronymi (Bromeliaceae). Bot. J. Linn. Soc. 2018, 187, 672–688. [Google Scholar] [CrossRef]
- Jankowska-Wroblewska, S.; Meyza, K.; Sztupecka, E.; Kubera, L.; Burczyk, J. Clonal structure and high genetic diversity at peripheral populations of Sorbus torminalis (L.) Crantz. iForest Biogeosciences For. 2016, 9, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, C.; Cummins, I.; Logan-Hines, E. Adapting Indigenous Agroforestry Systems for Integrative Landscape Management and Sustainable Supply Chain Development in Napo, Ecuador. In Integrating Landscapes: Agroforestry for Biodiversity Conservation and Food Sovereignty; Montagnini, F., Ed.; Springer: Cham, Switzerland, 2017; Volume 12, pp. 283–309. [Google Scholar]
- Ingvarsson, P.K.; Dahlberg, H. The effects of clonal forestry on genetic diversity in wild and domesticated stands of forest trees. Scand. J. For. Res. 2018, 34, 370–379. [Google Scholar] [CrossRef]
- Eckert, C.G.; Samis, K.E.; Lougheed, S.C. Genetic variation across species’ geographical ranges: The central-marginal hypothesis and beyond. Mol. Ecol. 2008, 17, 1170–1188. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Sun, G.; Wang, X.; Lu, J.; Wang, I.J.; Wang, Z. Population genetic structure is shaped by historical, geographic, and environmental factors in the leguminous shrub Caragana microphylla on the Inner Mongolia Plateau of China. BMC Plant Biol. 2017, 17, 200. [Google Scholar] [CrossRef] [Green Version]
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A.A. Genetic Isolation by Environment or Distance: Which Pattern of Gene Flow is Most Common? Evolution 2014, 68, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sagarin, R.D.; Gaines, S.D. The ‘abundant centre’ distribution: To what extent is it a biogeographical rule? Ecol. Lett. 2002, 5, 137–147. [Google Scholar] [CrossRef]
- Prugnolle, F.; De Meeûs, T. The impact of clonality on parasite population genetic structure. Parasite 2008, 15, 455–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Meeûs, T.; Balloux, F. Clonal reproduction and linkage disequilibrium in diploids: A simulation study. Infect. Genet. Evol. 2004, 4, 345–351. [Google Scholar] [CrossRef]
- Gitzendanner, M.A.; Weekley, C.W.; Germain-Aubrey, C.C.; Soltis, D.E.; Soltis, P.S. Microsatellite evidence for high clonality and limited genetic diversity in Ziziphus celata (Rhamnaceae), an endangered, self-incompatible shrub endemic to the Lake Wales Ridge, Florida, USA. Conserv. Genet. 2011, 13, 223–234. [Google Scholar] [CrossRef]
- Grimsby, J.L.; Tsirelson, D.; Gammon, M.A.; Kesseli, R. Genetic diversity and clonal vs. sexual reproduction in Fallopia spp. (Polygonaceae). Am. J. Bot. 2007, 94, 957–964. [Google Scholar] [CrossRef]
- Gabrielsen, T.M.; Brochmann, C. Sex after all: High levels of diversity detected in the arctic clonal plant Saxifraga cernuausing RAPD markers. Mol. Ecol. 2002, 7, 1701–1708. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef] [Green Version]
- Kreher, S.A.; Foré, S.A.; Collins, B.S. Genetic variation within and among patches of the clonal species, Vaccinium stamineum L. Mol. Ecol. 2000, 9, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Gargiulo, R.; Ilves, A.; Kaart, T.; Fay, M.F.; Kull, T. High genetic diversity in a threatened clonal species, Cypripedium calceolus (Orchidaceae), enables long-term stability of the species in different biogeographical regions in Estonia. Bot. J. Linn. Soc. 2018, 186, 560–571. [Google Scholar] [CrossRef]
- Aguillon, S.M.; Fitzpatrick, J.W.; Bowman, R.; Schoech, S.J.; Clark, A.G.; Coop, G.; Chen, N. Deconstructing isolation-by-distance: The genomic consequences of limited dispersal. PLoS Genet. 2017, 13, e1006911. [Google Scholar] [CrossRef] [Green Version]
- Foll, M.; Gaggiotti, O. Identifying the Environmental Factors That Determine the Genetic Structure of Populations. Genetics 2006, 174, 875–891. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-R.; Mitchell-Olds, T. Quantifying effects of environmental and geographical factors on patterns of genetic differentiation. Mol. Ecol. 2011, 20, 4631–4642. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef] [PubMed]
- Shafer, A.B.A.; Wolf, J.B.W. Widespread evidence for incipient ecological speciation: A meta-analysis of isolation-by-ecology. Ecol. Lett. 2013, 16, 940–950. [Google Scholar] [CrossRef]
- Temunović, M.; Franjić, J.; Satovic, Z.; Grgurev, M.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Environmental Heterogeneity Explains the Genetic Structure of Continental and Mediterranean Populations of Fraxinus angustifolia Vahl. PLoS ONE 2012, 7, e42764. [Google Scholar] [CrossRef] [Green Version]
- Poljak, I.; Idžojtić, M.; Šapić, I.; Korijan, P.; Vukelić, J. Diversity and structure of Croatian continental and alpine-Dinaric populations of grey alder (Alnus incana /L./ Moench subsp. incana): Isolation by distance and environment explains phenotypic divergence. Šumarski List 2018, 142, 19–32. [Google Scholar] [CrossRef]
- DeWoody, J.; Trewin, H.; Taylor, G. Genetic and morphological differentiation in Populus nigra L.: Isolation by colonization or isolation by adaptation? Mol. Ecol. 2015, 24, 2641–2655. [Google Scholar] [CrossRef] [Green Version]
- DeSilva, R.; Dodd, R.S. Fragmented and isolated: Limited gene flow coupled with weak isolation by environment in the paleoendemic giant sequoia (Sequoiadendron giganteum). Am. J. Bot. 2019, 107, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Hellwig, T.; Abbo, S.; Sherman, A.; Coyne, C.J.; Saranga, Y.; Lev-Yadun, S.; Main, D.; Zheng, P.; Ophir, R. Limited divergent adaptation despite a substantial environmental cline in wild pea. Mol. Ecol. 2020, 29, 4322–4336. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, S.; Meirmans, P.G.; Aitken, S.N.; Ritland, K.; Isabel, N. The challenge of separating signatures of local adaptation from those of isolation by distance and colonization history: The case of two white pines. Ecol. Evol. 2016, 6, 8649–8664. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Wu, J.; Wang, Z.; Hou, H.; Gao, Y.; Han, W.; Ru, W.; Sun, G.; Wang, Y. Population genetic variation characterization of the boreal tree Acer ginnala in Northern China. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Marcer, A.; Méndez-Vigo, B.; Alonso-Blanco, C.; Picó, F.X. Tackling intraspecific genetic structure in distribution models better reflects species geographical range. Ecol. Evol. 2016, 6, 2084–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Griffin, T.S.; Kraner, D.; Schaffner, M.K.; Sharma, D.; Hazel, M.; Leitch, A.R.; Orians, C.M.; Han, W.; Stepp, J.R.; et al. Environmental Factors Variably Impact Tea Secondary Metabolites in the Context of Climate Change. Front. Plant Sci. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cansian, R.L.; Mossi, A.; Mosele, S.H.; Toniazzo, G.; Treichel, H.; Paroul, N.; Oliveira, J.V.D.; Mazutti, M.; Echeverrigaray, S. Genetic conservation and medicinal properties of mate (Ilex paraguariensis St Hil.). Pharmacogn. Rev. 2008, 2, 326–338. [Google Scholar]
- Gan, R.-Y.; Zhang, D.; Wang, M.; Corke, H. Health Benefits of Bioactive Compounds from the Genus Ilex, a Source of Traditional Caffeinated Beverages. Nutrients 2018, 10, 1682. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.K.; Mukherjee, A.; Guo, W.; Ng, W.L.; Huang, Y. Combining ecological niche modeling with genetic lineage information to predict potential distribution of Mikania micrantha Kunth in South and Southeast Asia under predicted climate change. Glob. Ecol. Conserv. 2019, 20, e00800. [Google Scholar] [CrossRef]
- Lathrap, D.W. The antiquity and importance of long-distance trade relationships in the moist tropics of pre-Columbian South America. World Archaeol. 1973, 5, 170–186. [Google Scholar] [CrossRef]
- Renner, S.S. A History of Botanical Exploration in Amazonian Ecuador, 1739–1988. Smithson. Contrib. Bot. 1993, 82, 1–39. [Google Scholar] [CrossRef]
- Spruce, R. Notes of a Botanist on the Amazon and Andes; Macmillan and Company: London, UK, 1908. [Google Scholar]
- Li, W.; Wang, B.; Wang, J. Lack of genetic variation of an invasive clonal plant Eichhornia crassipes in China revealed by RAPD and ISSR markers. Aquat. Bot. 2006, 84, 176–180. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | N | G | effG | D | E | ShW |
|---|---|---|---|---|---|---|
| Northern | 44 | 34 | 27.657 | 0.986 | 0.813 | 1.924 a |
| Central | 31 | 30 | 29.121 | 0.998 | 0.971 | 2.670 b |
| Southern | 13 | 12 | 11.267 | 0.987 | 0.939 | 1.893 a |
| Regions | N | A | PA | AR | Ho | He | FIS |
|---|---|---|---|---|---|---|---|
| Northern | 34 | 60 | 23 | 2.483 | 0.452 | 0.356 | −0.106 |
| Central | 30 | 66 | 30 | 3.193 | 0.435 | 0.444 | 0.162 |
| Southern | 12 | 36 | 3 | 2.118 | 0.455 | 0.308 | −0.379 |
| Overall | 76 | 95 | - | 4.981 | 0.448 | 0.396 | −0.185 |
| Source of Variation | df | Sum Sq | Mean Sq | Est. Var | % of Variative |
|---|---|---|---|---|---|
| Among regions | 2 | 19.247 | 9.624 | 0.125 | 3.00 |
| Within regions | 149 | 557.483 | 3.741 | 3.741 | 97.00 |
| Overall | 151 | 576.730 | - | 3.867 | 100.00 |
| Model | Cluster 1 | Cluster 2 | Species |
|---|---|---|---|
| Cluster 1 | 1.000 | 0.426 | 0.875 |
| Cluster 2 | - | 1.000 | 0.475 |
| Species | - | - | 1.000 |
| Bioclimatic Variable * | Species | Cluster 1 | Cluster 2 |
|---|---|---|---|
| Bio 2 | 2.5 | 2.8 | 0.0 |
| Bio 3 | 3.4 | 1.0 | 0.0 |
| Bio 4 | 5.8 | 4.1 | 8.4 |
| Bio 8 | 42.6 | 46.1 | 15.4 |
| Bio 15 | 1.5 | 2.2 | 76.2 |
| Bio 17 | 44.2 | 43.8 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erazo-Garcia, M.P.; Guadalupe, J.J.; Rowntree, J.K.; Borja-Serrano, P.; Espinosa de los Monteros-Silva, N.; Torres, M.d.L. Assessing the Genetic Diversity of Ilex guayusa Loes., a Medicinal Plant from the Ecuadorian Amazon. Diversity 2021, 13, 182. https://doi.org/10.3390/d13050182
Erazo-Garcia MP, Guadalupe JJ, Rowntree JK, Borja-Serrano P, Espinosa de los Monteros-Silva N, Torres MdL. Assessing the Genetic Diversity of Ilex guayusa Loes., a Medicinal Plant from the Ecuadorian Amazon. Diversity. 2021; 13(5):182. https://doi.org/10.3390/d13050182
Chicago/Turabian StyleErazo-Garcia, Maria P., Juan José Guadalupe, Jennifer K. Rowntree, Pamela Borja-Serrano, Nina Espinosa de los Monteros-Silva, and Maria de Lourdes Torres. 2021. "Assessing the Genetic Diversity of Ilex guayusa Loes., a Medicinal Plant from the Ecuadorian Amazon" Diversity 13, no. 5: 182. https://doi.org/10.3390/d13050182