
Characterization of Five New Earthworm Mitogenomes (Oligochaeta: Lumbricidae): Mitochondrial Phylogeny of Lumbricidae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction and Sequencing
2.3. Assembly of Mitogenomes
2.4. Data Analysis
3. Results
3.1. Characterization of Mitogenomes
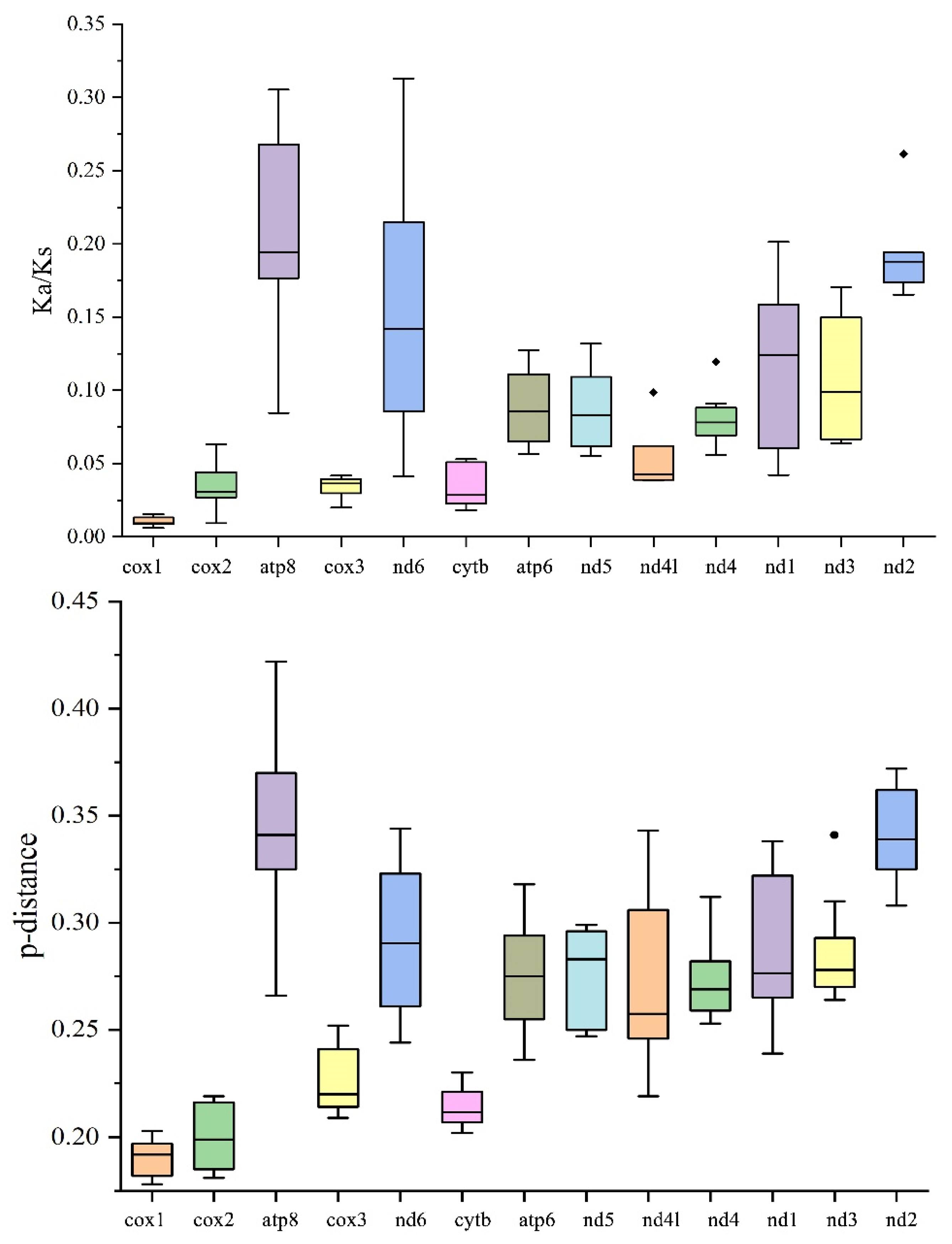
3.2. Variation among PCGs
3.3. A Mitochondrial Genome Phylogeny of Lumbricidae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klarica, J.; Kloss-Brandstätter, A.; Traugott, M.; Juen, A. Comparing four mitochondrial genes in earthworms—Implications for identification, phylogenetics, and discovery of cryptic species. Soil Biol. Biochem. 2012, 45, 23–30. [Google Scholar] [CrossRef]
- Yonemura, S.; Kaneda, S.; Kodama, N.; Sakurai, G.; Yokozawa, M. Dynamic measurements of earthworm respiration. J. Agric. Meteorol. 2019, 75, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Losada, M.; Ricoy, M.; Marshall, J.C.; Domínguez, J. Phylogenetic assessment of the earthworm Aporrectodea caliginosa species complex (Oligochaeta: Lumbricidae) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenetics Evol. 2009, 52, 293–302. [Google Scholar] [CrossRef]
- Richard, B.; Decaëns, T.; Rougerie, R.; James, S.W.; Porco, D.; Hebert, P.D.N. Re-Integrating earthworm juveniles into soil biodiversity studies: Species identification through DNA barcoding. Mol. Ecol. Resour. 2010, 10, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Gabriella, J.E.; Jan, L.; Claudio, M.D.; Alberto, P.C. Identification of Earthworm Species in Uruguay Based on Morphological and Molecular Methods. Agrociencia 2019, 23, 37–46. [Google Scholar]
- Piearce, T.G.; Oates, K.; Carruthers, W.J. A fossil earthworm embryo (Oligochaeta) from beneath a Late Bronze Age midden at Potterne, Wiltshire, UK. J. Zool. 1990, 220, 537–542. [Google Scholar] [CrossRef]
- Briones, M.J.I. A taxonomic study of the Allolobophora caliginosa complex (Oligochaeta, Lumbricidae): A preliminary study. Can. J. Zool. 1996, 74, 240–244. [Google Scholar] [CrossRef]
- Briones, M.J.I. Two ecotypes in Allolobophora caliginosa (Oligochaeta, Lumbricidae). Acta Oecologica 1993, 14, 317–325. [Google Scholar]
- Fang, Y.; Chen, J.; Ruan, H.H.; Xu, N.; Que, Z.T.; Liu, H.Y. Genetic Diversity and Population Structure of Metaphire vulgaris Based on the Mitochondrial COI Gene and Microsatellites. Front. Genet. 2021, 12, 686246. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Ding, J.Y.; Lin, S.P.; Xu, R.F.; Liu, H.Y. Comparative mitogenomes of three species in Moenkhausia: Rare irregular gene rearrangement within Characidae. Int. J. Biol. Macromol. 2021, 183, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Lin, S.P.; Liu, H.Y. Mitochondrial genomes of five Hyphessobrycon tetras and their phylogenetic implications. Ecol. Evol. 2021, 11, 12754–12764. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Xiao, N.W. Terrestrial Earthworms (Oligochaeta: Opisthopora) of China; China Agriculture Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.K.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Braband, A.; Schierwater, B.; Stadler, P.F. Genetic aspects of mitochondrial genome evolution. Mol. Phylogenetics Evol. 2013, 69, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Zhang, H.K.; Gao, S.H.; Lercher, M.J.; Hu, S.N.; Chen, W.H. EvolView, an online tool for visualizing, annotating and managing phylogenetic trees. Nucleic Acids Res. 2012, 40, W569–W572. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Peltek, S.E. The complete mitochondrial genome of Aporrectodea rosea (Annelida: Lumbricidae). Mitochondrial DNA Part B-Resour. 2019, 4, 1752–1753. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.Z.; Liu, H.Y.; Zhang, Y.F.; Ruan, H.H. The complete mitochondrial genome of Lumbricus rubellus (Oligochaeta, Lumbricidae) and its phylogenetic analysis. Mitochondrial DNA Part B-Resour. 2019, 4, 2677–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Boore, J.L.; Brown, W.M. Complete sequence of the mitochondrial DNA of the annelid worm Lumbricus terrestris. Genetics 1995, 141, 305–319. [Google Scholar] [CrossRef]
- Conrado, A.C.; Arruda, H.; Stanton, D.W.G.; James, S.W.; Kille, P.; Brown, G.; Silva, E.; Dupont, L.; Taheri, S.; Morgan, A.J.; et al. The complete mitochondrial DNA sequence of the pantropical earthworm Pontoscolex corethrurus (Rhinodrilidae, Clitellata): Mitogenome characterization and phylogenetic positioning. ZooKeys 2017, 688, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Sechi, P.; Yuan, M.; Jiang, J.; Dong, Y.; Qiu, J. Fifteen new earthworm mitogenomes shed new light on phylogeny within the Pheretima complex. Sci. Rep. 2016, 6, 20096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, J.C.; Wu, C.I. The neutral theory in the genomic era. Curr. Opin. Genet. Dev. 2001, 11, 642–646. [Google Scholar] [CrossRef]
- Yang, Z.; Nielsen, R. Estimating synonymous and nonsynonymous substitution rates under realistic evolutionary models. Mol. Biol. Evol. 2000, 17, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, Z.; Vang, S.; Yu, J.; Wong, G.K.S.; Wang, J. Correlation Between Ka/Ks and Ks is Related to Substitution Model and Evolutionary Lineage. J. Mol. Evol. 2009, 68, 414–423. [Google Scholar] [CrossRef]
- Cui, Z.X.; Liu, Y.; Chu, K.H. Broader pattern of tandem repeats in the mitochondrial control region of Perciformes. Chin. J. Oceanol. Limnol. 2010, 28, 785–794. [Google Scholar] [CrossRef]
- Kornienko, I.V.; Vodolazhsky, D.I. Patterns of single nucleotide polymorphism distribution in hypervariable regions of the D-loop of human mitochondrial DNA. Mol. Biol. 2010, 44, 386–392. [Google Scholar] [CrossRef]
- King, R.A.; Tibble, A.L.; Symondson, W.O. Opening a can of worms: Unprecedented sympatric cryptic diversity within British lumbricide earthworms. Mol. Ecol. 2008, 17, 4684–4698. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Breinholt, J.W.; Stojanovic, M.; James, S.W.; Pérez-Losada, M. Underground evolution: New roots for the old tree of lumbricid earthworms. Mol. Phylogenetics Evol. 2015, 83, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Martinsson, S.; Erséus, C. Hybridisation and species delimitation of Scandinavian Eisenia spp. (Clitellata: Lumbricidae). Eur. J. Soil Biol. 2018, 88, 41–47. [Google Scholar] [CrossRef]
- Martinsson, S.; Erséus, C. Cryptic speciation and limited hybridization within Lumbricus earthworms (Clitellata: Lumbricidae). Mol. Phylogenetics Evol. 2017, 106, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Y.; Wang, C.; Zhang, T.; Al-Farraj, S.A.; Gao, F. Further consideration on the phylogeny of the Ciliophora: Analyses using both mitochondrial and nuclear data with focus on the extremely confused class Phyllopharyngea. Mol. Phylogenetics Evol. 2017, 112, 96–106. [Google Scholar] [CrossRef]
- Katz, L.A.; DeBerardinis, J.; Hall, M.S.; Kovner, A.M.; Dunthorn, M.; Muse, S.V. Heterogeneous Rates of Molecular Evolution Among Cryptic Species of the Ciliate Morphospecies Chilodonella uncinata. J. Mol. Evol. 2011, 73, 266–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Z.; Han, Q.D.; Fu, L.W.; Wang, Y.X.; Sui, Z.H.; Liu, Y.G. Molecular identification and phylogenetic analysis of fungal pathogens isolated from diseased fish in Xinjiang, China. J. Fish Biol. 2021, 1–12. [Google Scholar] [CrossRef]
- Fita, T.; Getu, E.; Wakgari, M.; Woldetsadike, K.; Jones, P.R. Molecular identification of white mango scale, Aulacaspis tubercularis Newstead (Homoptera: Diaspididae), and its associated natural enemies in western Ethiopia. Int. J. Trop. Insect Sci. 2021. [Google Scholar] [CrossRef]
- Breka, K.; Krizmanić, I.; Vukov, T.; Stamenković, S. A procedure for taxon assessment based on morphological variation in European water frogs (Pelophylax esculentus complex). Turk. J. Zool. 2020, 44, 215–223. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Sample ID | Coordination |
|---|---|---|
| Bimastos parvus | GS1 | 39°46′ N, 98°19′ E |
| Dendrobaena octaedra | CBS1 | 44°2′ N, 127°55′ E |
| Eisenia andrei | LFWA | 38°87′ N, 116°47′ E |
| Eisenia nordenskioldi | LSFL | 47°10′ N, 128°53′ E |
| Octolasion tyrtaeum | XZLS | 29°46′ N, 91°55′ E |
| Family | Species | Accession no. | Length (bp) | References |
|---|---|---|---|---|
| Lumbricidae | Aporrectodea rosea | MK573632.1 | 15,086 | [22] |
| Bimastos parvus | MZ857199 | 15,194 | ||
| Dendrobaena octaedra | MZ857197 | 15,715 | ||
| Eisenia andrei | MZ857198 | 15,656 | ||
| Eisenia balatonica | MK642872.1 | 14,589 | [23] | |
| Eisenia nana | MK618511.1 | 14,599 | [23] | |
| Eisenia nordenskioldi | MZ857200 | 15,152 | ||
| Eisenia spelaea | MK642870.1 | 14,738 | [23] | |
| Eisenia tracta | MK642871.1 | 14,589 | [23] | |
| Lumbricus rubellus | MN102127.1 | 15,464 | [24] | |
| Lumbricus terrestris | U24570.1 | 14,998 | [25] | |
| Octolasion tyrtaeum | MZ857201 | 14,977 | ||
| Glossoscolecidae | Pontoscolex corethrurus | KT988053.1 | 14,835 | [26] |
| Model | cox1 | cox2 | atp8 | cox3 | nd6 | cytb | atp6 | nd5 | nd4l | nd4 | nd1 | nd3 | nd2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BI | GTR+F+ I+G4 | GTR+F+ I+G4 | HKY+F+ I+G4 | GTR+F+ I+G4 | HKY+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | HKY+F+ I+G4 |
| ML | TIM2+F+ I+G4 | TIM2+F+ I+G4 | HKY+F+ I+G4 | TIM2+F+ I+G4 | HKY+F+ I+G4 | TIM2+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | GTR+F+ I+G4 | HKY+F+ I+G4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Zhang, Y.; Xu, W.; Fang, Y.; Ruan, H. Characterization of Five New Earthworm Mitogenomes (Oligochaeta: Lumbricidae): Mitochondrial Phylogeny of Lumbricidae. Diversity 2021, 13, 580. https://doi.org/10.3390/d13110580
Liu H, Zhang Y, Xu W, Fang Y, Ruan H. Characterization of Five New Earthworm Mitogenomes (Oligochaeta: Lumbricidae): Mitochondrial Phylogeny of Lumbricidae. Diversity. 2021; 13(11):580. https://doi.org/10.3390/d13110580
Chicago/Turabian StyleLiu, Hongyi, Yufeng Zhang, Wei Xu, Yu Fang, and Honghua Ruan. 2021. "Characterization of Five New Earthworm Mitogenomes (Oligochaeta: Lumbricidae): Mitochondrial Phylogeny of Lumbricidae" Diversity 13, no. 11: 580. https://doi.org/10.3390/d13110580