
Breeding Density and Collision Mortality of Loggerhead Shrike (Lanius ludovicianus) in the Altamont Pass Wind Resource Area


Abstract
:1. Introduction
2. Materials and Methods
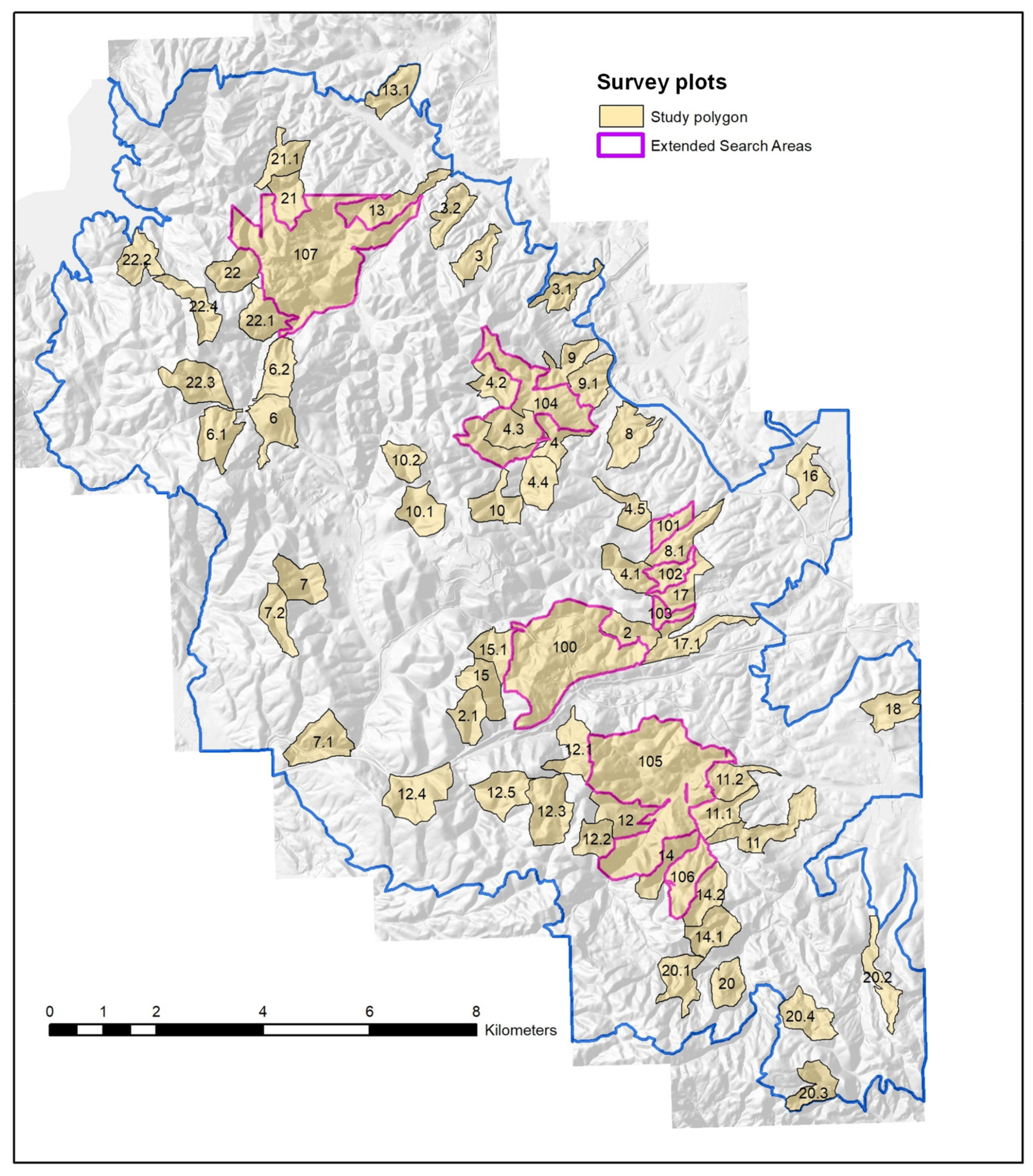
2.1. Study Area
2.2. Field Methods to Estimate Density
2.3. Wind Turbine Fatality Estimates
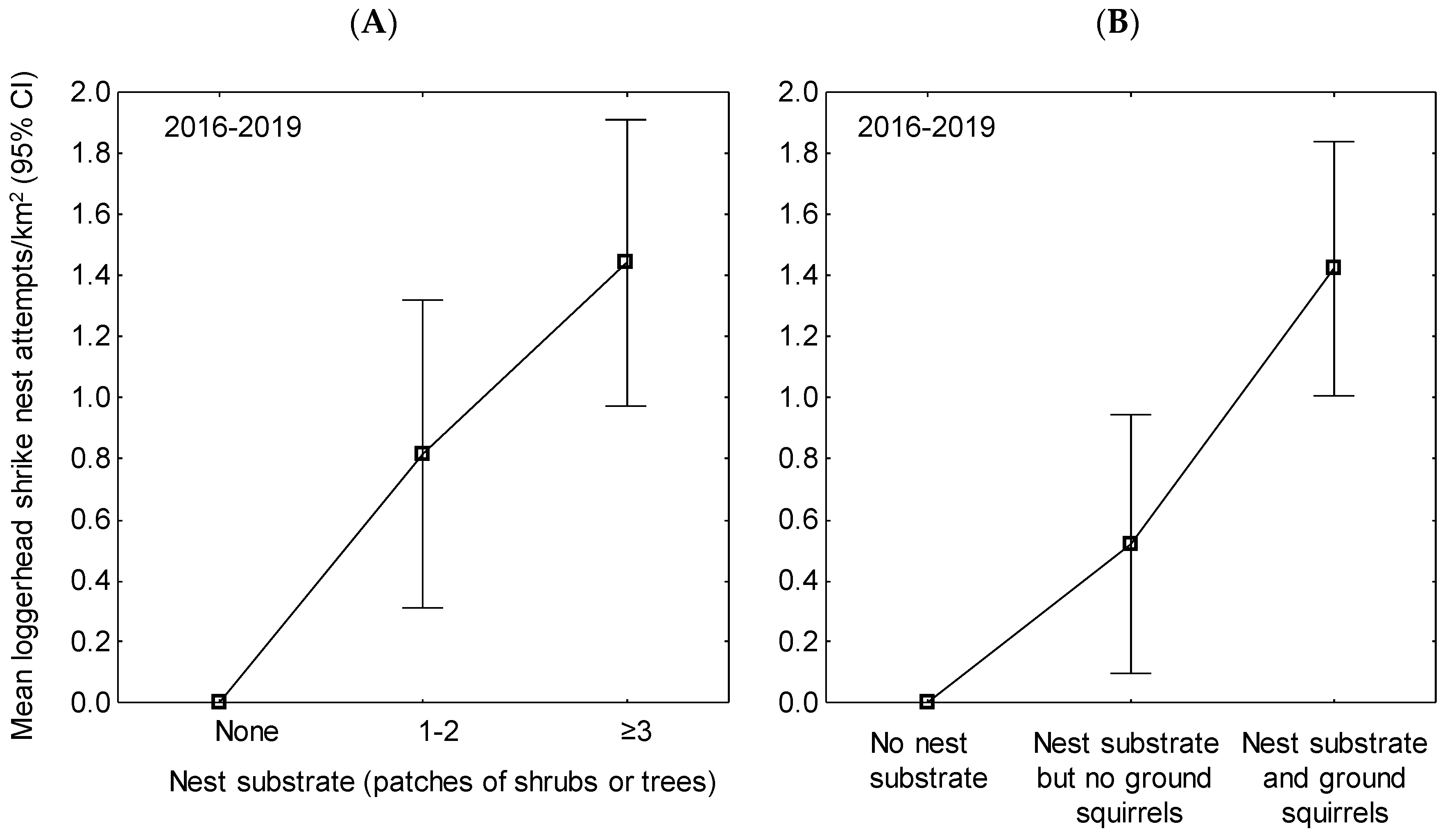
3. Results
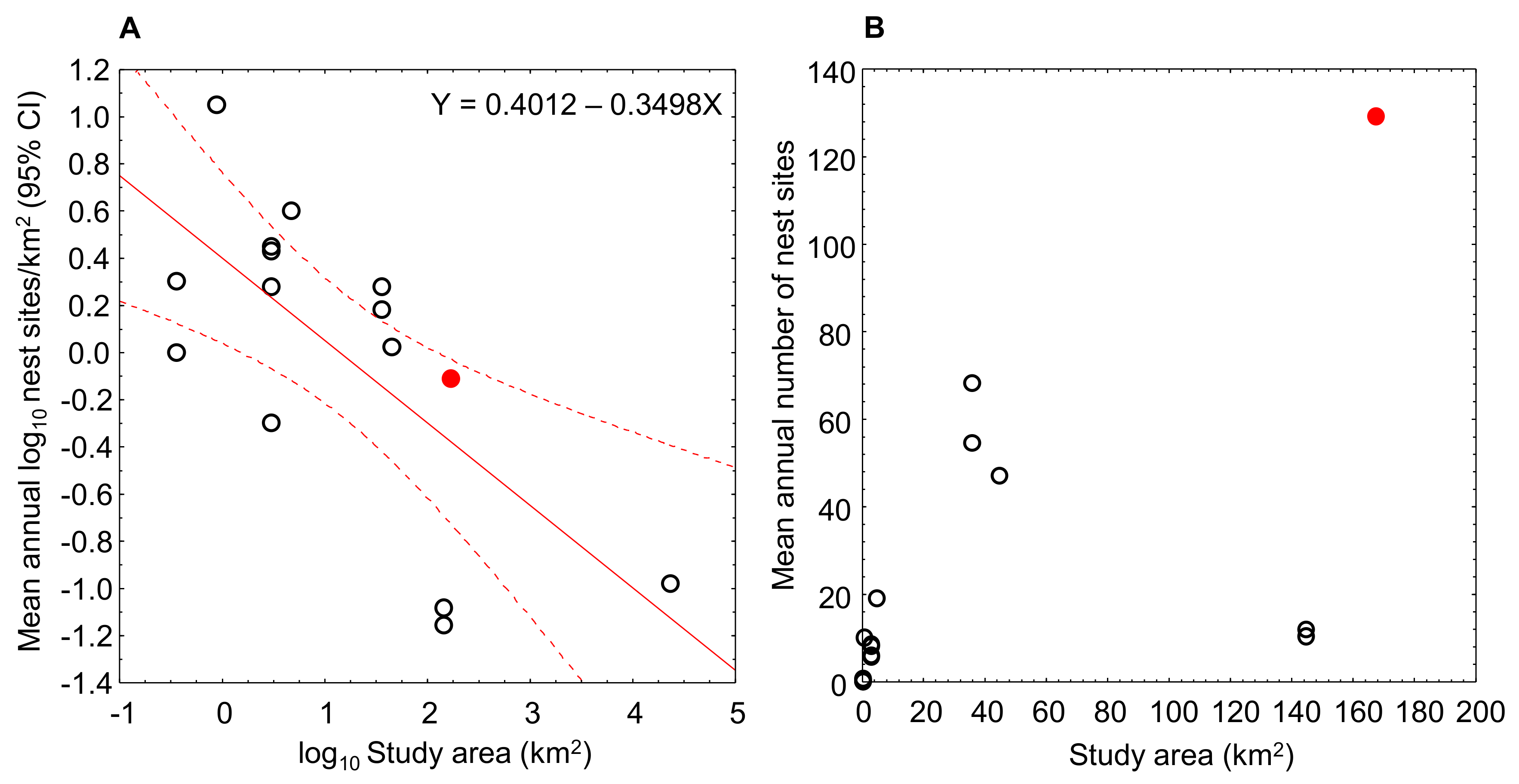
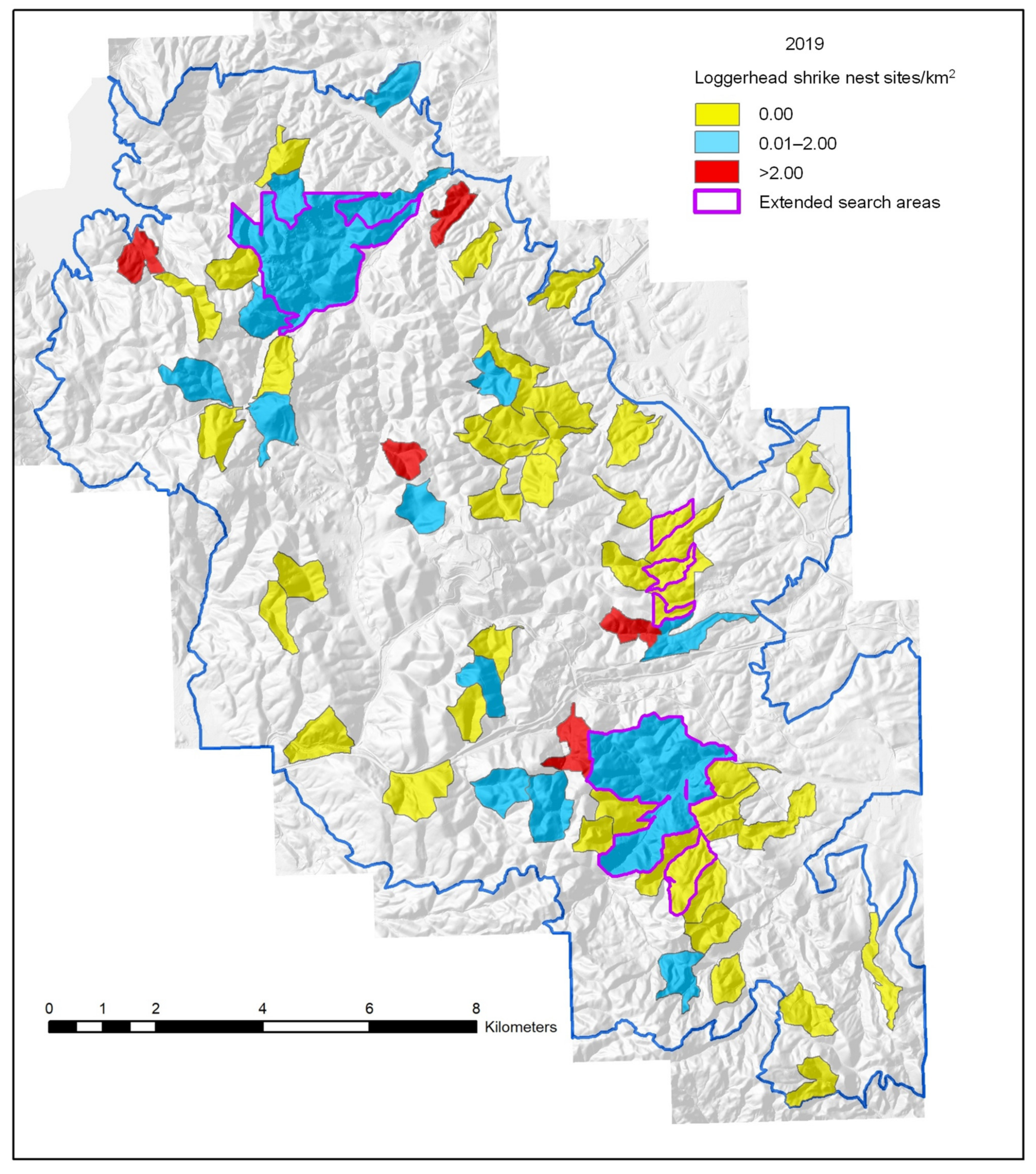
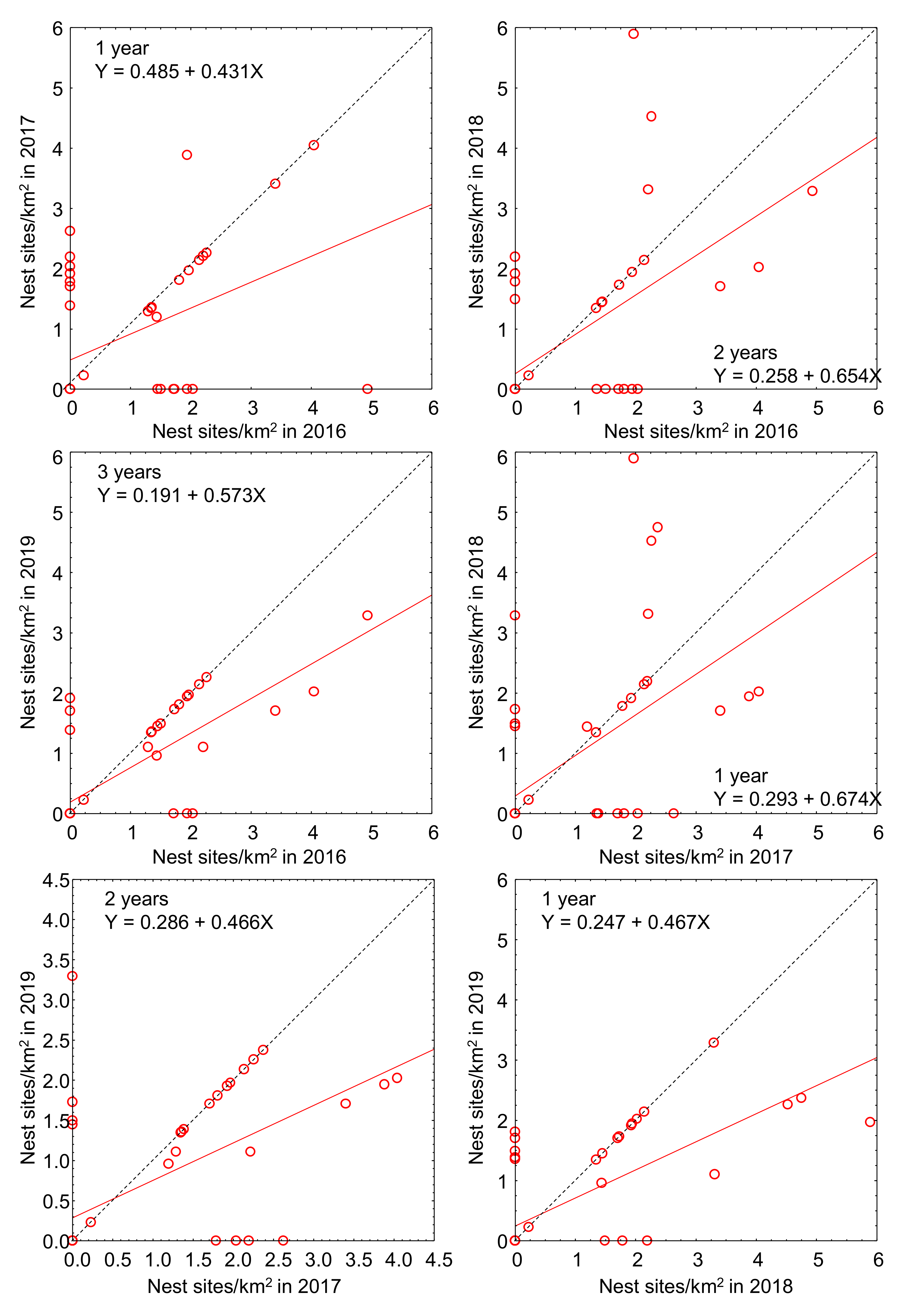
3.1. Breeding Density Estimates
3.2. Wind Turbine Fatality Estimates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yosef, R.; Layne, J.N.; Lohrer, F.E. Trends in numbers of loggerhead shrikes on roadside censuses in Peninsular Florida, 1974–1992. Fla. Sci. 1993, 56, 92–97. [Google Scholar]
- Cade, T.J.; Woods, C.P. Changes in distribution and abundance of the Loggerhead Shrike. Conserv. Biol. 1997, 11, 21–31. [Google Scholar] [CrossRef]
- Sauer, J.R.; Hines, J.E.; Fallon, J.E.; Pardieck, K.L.; Ziolkowski, D.J., Jr.; Link, W.A. The North American Breeding Bird Survey, Results and Analysis 1966–2010; Version 12.07.2011; USGS Patuxent Wildlife Research Center: Laurel, MD, USA, 2011.
- Shuford, W.D.; Gardali, T. (Eds.) California Bird Species of Special Concern: A Ranked Assessment of Species, Subspecies, and Distinct Populations of Birds of Immediate Conservation Concern in California; Studies of Western Birds 1; Western Field Ornithologists: Camarillo, CA, USA, 2008. [Google Scholar]
- Smallwood, K.S.; Nakamoto, B. Impacts of West Nile Virus Epizootic on Yellow-Billed Magpie, American Crow, and other Birds in the Sacramento Valley, California. Condor 2009, 111, 247–254. [Google Scholar] [CrossRef]
- Woods, C.P. Status of loggerhead shrikes in the sagebrush habitat of southwestern Idaho. Proc. West. Found. Vertebr. Zool. 1995, 6, 150–154. [Google Scholar]
- Luukkonen, D.R. Status and Breeding Ecology of the Loggerhead Shrike in Virginia. Master’s Thesis, Virginia Polytechnical Institute and State University, Blacksburg, VA, USA, 1987. [Google Scholar]
- Novak, P.G. Habitat selection by breeding Loggerhead Shrikes in northern New York. Proc. West. Found. Vertebr. Zool. 1995, 6, 176–181. [Google Scholar]
- Andersen, D.E.; Rongstad, O.J. Surveys for wintering birds of prey in southeastern Colorado: 1983–1988. J. Raptor Res. 1989, 23, 152–156. [Google Scholar]
- Kridelbaugh, A. Nesting ecology of the Loggerhead Shrike in Central Missouri. Wilson Bull. 1983, 95, 303–308. [Google Scholar]
- DeGeus, D.W. Productivity and Habitat Preferences of Loggerhead Shrikes Inhabiting Roadsides in a Midwestern Agroenvironment. Iowa State University Retrospective Theses and Dissertations. 1990. Available online: https://lib.dr.iastate.edu/rtd/16818 (accessed on 27 October 2021).
- Scott, T.A.; Morrison, M.L. Natural history and management of the San Clemente loggerhead shrike. Proc. West. Found. Vertebr. Zool. 1990, 4, 23–57. [Google Scholar]
- Thogmartin, W.E.; Howe, F.P.; James, F.C.; Johnson, D.H.; Reed, E.T.; Sauer, J.R.; Thompson, F.R., III. A review of the population estimation approach of the North American Landbird Conservation Plan. Auk 2006, 123, 892–904. [Google Scholar] [CrossRef]
- Smallwood, K.S. Linking habitat restoration to meaningful units of animal demography. Restor. Ecol. 2001, 9, 253–261. [Google Scholar] [CrossRef]
- Smallwood, K.S. Scaling Swainson’s hawk population density for assessing habitat-use across an agricultural landscape. J. Raptor Res. 1995, 29, 172–178. [Google Scholar]
- Smallwood, K.S. On the evidence needed for listing northern goshawks (Accipter gentilis) under the Endangered Species Act: A reply to Kennedy. J. Raptor Res. 1998, 32, 323–329. [Google Scholar]
- Smallwood, K.S.; Morrison, M.L. Nest-site selection in a high-density colony of burrowing owls. J. Raptor Res. 2018, 52, 454–470. [Google Scholar] [CrossRef]
- Smallwood, K.S.; Neher, L.; Mount, J.; Culver, R.C.E. Nesting Burrowing Owl Abundance in the Altamont Pass Wind Resource Area, California. Wildl. Soc. Bull. 2013, 37, 787–795. [Google Scholar] [CrossRef]
- Smallwood, K.S. Long search intervals under-estimate bird and bat fatalities caused by wind turbines. Wildl. Soc. Bull. 2017, 41, 224–230. [Google Scholar] [CrossRef]
- Smallwood, K.S.; Bell, D.A.; Walther, E.L.; Leyvas, E.; Standish, S.; Mount, J.; Karas, B. Estimating wind turbine fatalities using integrated detection trials. J. Wildl. Manag. 2018, 82, 1169–1184. [Google Scholar] [CrossRef]
- Smallwood, K.S. Comparing bird and bat fatality-rate estimates among North American wind-energy projects. Wildl. Soc. Bull. 2013, 37, 19–33. [Google Scholar] [CrossRef]
- Brown, K.; Smallwood, K.S.; Szewczak, J.; Karas, B. Final 2012–2015 Report Avian and Bat Monitoring Project Vasco Winds; LLC. Prepared for NextEra Energy Resources: Livermore, CA, USA, 2016. [Google Scholar]
- T. Harvey & Associates. Golden Hills Wind Energy Center Post-Construction Fatality Monitoring Report: Final 3-Year Report; Prepared for Golden Hills Wind, LLC: Livermore, CA, USA, 2020. [Google Scholar]
- Smallwood, K.S.; Bell, D.A.; Standish, S. Dogs detect larger wind energy impacts on bats and birds. J. Wildl. Manag. 2020, 84, 852–864. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.L.; Kuehler, C.M.; Scott, T.A.; Lieberman, A.A.; Everett, W.T.; Phillips, R.B.; Koehler, C.E.; Aigner, P.A.; Winchell, C.; Burr, T. San Clemente loggerhead shrike: Recovery plan for an endangered species. Proc. West. Found. Vertebr. Zool. 1995, 6, 293–295. [Google Scholar]
- Bjorge, R.R.; Prescott, D.R.C. Population estimate and habitat associations of the loggerhead shrike, Lanius ludovicianus, in southeastern Alberta. Can. Field-Nat. 1996, 110, 445–449. [Google Scholar]
- Collister, D.M.; de Smet, K. Breeders and natural dispersal in the loggerhead shrike. J. Field Ornithol. 1997, 68, 273–282. [Google Scholar]
- Medin, D.E. Grazing and Passerine breeding birds in a Great Basin low-shrub desert. Great Basin Nat. 1986, 46, 567–572. [Google Scholar]
- Gawlik, D.E.; Bildstein, K.L. Reproductive success and nesting habitat of loggerhead shrikes in north-central South Carolina. Wilson Bull. 1990, 102, 37–48. [Google Scholar]
- Poole, L.D. Reproductive Success and Nesting Habitat of Loggerhead Shrikes in Shrubsteppe Communities. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1992. [Google Scholar]
- Collister, D.M.; Wilson, S. Territory size and foraging habitat of loggerhead shrikes (Lanius ludovicianus) in southeastern Alberta. J. Raptor Res. 2007, 41, 130–138. [Google Scholar] [CrossRef]
- Taylor, R.A.J.; Taylor, L.R. A behavioral model for the evolution of spatial dynamics. In Population Dynamics; Anderson, R.M., Turner, B.D., Taylor, L.R., Eds.; Blackwell Scientific Publications: Oxford, UK, 1979; pp. 1–28. [Google Scholar]
- Guo, X.; He, Y.; Wilmshurst, J.F. Managing Loggerhead Shrike Habitat Using Remote Sensing Products. In Proceedings of the American Society of Photogrammetry and Remote Sensing Annual Confernce, ASPRS 2009, Baltimore, MD, USA, 8–13 March 2009; Available online: https://www.asprs.org/a/publications/proceedings/baltimore09/0027.pdf (accessed on 30 September 2021).

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Number of Plots | Nest Attempts/km2 | APWRA Nest Attempts | ||
|---|---|---|---|---|---|
| x | 95% CI | x | 95% CI | ||
| 2016 | 47 | 0.804 | 0.444–1.165 | 134.8 | 74.4–195.2 |
| 2017 | 49 | 0.826 | 0.483–1.168 | 138.4 | 81.0–195.8 |
| 2018 | 51 | 0.814 | 0.412–1.215 | 136.4 | 69.1–203.7 |
| 2019 | 52 | 0.635 | 0.377–0.893 | 106.4 | 63.1–149.6 |
| Plot | Area, Ha | Years | Nest Attempts/km2 | Ground Squirrel Abundance | Nest Substrate | |
|---|---|---|---|---|---|---|
| x | 95% CI | |||||
| 2 | 49.085 | 4 | 2.824 | 1.027–4.622 | Very few | Moderate |
| 2.1 | 44.257 | 4 | 0.509 | 0.000–2.130 | Very few | Scarce |
| 3 | 45.739 | 4 | 0.000 | Many | None | |
| 3.1 | 47.06 | 4 | 0.000 | Moderate | None | |
| 3.2 | 46.737 | 4 | 2.140 | 2.140–2.140 | Many | Scarce |
| 4 | 41.008 | 4 | 0.000 | Moderate | None | |
| 4.1 | 50.75 | 4 | 0.000 | Very few | None | |
| 4.2 | 51.481 | 4 | 2.428 | 0.883–3.974 | Moderate | Moderate |
| 4.3 | 66.65 | 4 | 0.000 | Many | None | |
| 4.4 | 66.94 | 4 | 0.000 | Some | None | |
| 4.5 | 38.05 | 4 | 0.657 | 0.000–2.748 | Some | Moderate |
| 6 | 72.176 | 4 | 0.693 | 0.000–1.966 | Moderate | Moderate |
| 6.1 | 67 | 4 | 0.373 | 0.000–1.561 | None | Moderate |
| 6.2 | 57.227 | 4 | 0.000 | None | Scarce | |
| 7 | 61.185 | 3 | 0.000 | Moderate | None | |
| 7.2 | 48.769 | 3 | 0.000 | Some | None | |
| 8 | 72.792 | 3 | 0.000 | Many | None | |
| 8.1 | 57.411 | 4 | 0.000 | Many | None | |
| 10 | 53.519 | 4 | 0.000 | Few | None | |
| 10.1 | 58.643 | 4 | 0.853 | 0.000–2.419 | Moderate | Scarce |
| 10.2 | 42.148 | 3 | 3.163 | 0.000–6.566 | Moderate | Moderate |
| 11 | 84.234 | 4 | 0.000 | Very few | None | |
| 11.1 | 53.701 | 4 | 0.000 | Very few | None | |
| 11.2 | 53.38 | 4 | 0.000 | None | None | |
| 12 | 61.759 | 4 | 0.000 | Some | None | |
| 12.1 | 60.738 | 4 | 2.881 | 0.000–6.178 | Very few | Moderate |
| 12.2 | 41.261 | 4 | 0.000 | Very few | None | |
| 12.3 | 90.475 | 4 | 2.211 | 0.775–3.647 | Few | Moderate |
| 12.5 | 93.027 | 4 | 0.749 | 0.000–2.125 | Many | Moderate |
| 13 | 74.084 | 4 | 1.350 | 1.350–1.350 | Few | Moderate |
| 13.1 | 50.885 | 4 | 2.948 | 0.000–6.075 | Moderate | Scarce |
| 14 | 51.331 | 4 | 0.487 | 0.000–2.037 | Few | Scarce |
| 14.1 | 58.149 | 4 | 0.430 | 0.000–1.798 | Few | Scarce |
| 14.2 | 53.197 | 4 | 0.000 | Few | None | |
| 15 | 55.36 | 4 | 1.355 | 0.000–2.792 | Few | Moderate |
| 15.1 | 48.773 | 4 | 0.000 | Few | Scarce | |
| 16 | 52.773 | 3 | 0.000 | Moderate | Scarce | |
| 17 | 57.953 | 4 | 0.000 | Few | None | |
| 17.1 | 69.087 | 4 | 1.086 | 0.000–2.237 | Moderate | Scarce |
| 18 | 54.597 | 1 | 0.000 | None | None | |
| 20 | 45.531 | 4 | 1.098 | 0.000–3.116 | None | Moderate |
| 20.1 | 58.718 | 4 | 2.555 | 0.990–4.119 | Few | Moderate |
| 20.2 | 63.108 | 4 | 0.000 | None | Scarce | |
| 20.3 | 49.819 | 4 | 0.000 | Few | None | |
| 20.4 | 62.357 | 4 | 0.000 | None | None | |
| 21 | 51.993 | 4 | 1.443 | 0.000–2.973 | Very few | Scarce |
| 21.1 | 49.031 | 4 | 0.510 | 0.000–2.133 | Moderate | Scarce |
| 22 | 56.015 | 4 | 0.893 | 0.000–2.533 | None | Moderate |
| 22.1 | 57.817 | 4 | 1.297 | 0.000–2.673 | None | Moderate |
| 22.2 | 49.445 | 4 | 3.034 | 1.176–4.892 | Very few | Moderate |
| 22.3 | 73.676 | 4 | 1.018 | 0.000–2.098 | None | Scarce |
| 22.4 | 55.193 | 1 | 0.000 | None | Moderate | |
| 101 | 32 | 2 | 0.000 | Moderate | None | |
| 102 | 34 | 2 | 0.000 | Many | None | |
| 103 | 22 | 2 | 0.000 | Many | None | |
| 104 | 250 | 3 | 0.000 | Few | None | |
| 105 | 439 | 4 | 0.228 | 0.228–0.228 | Moderate | Moderate |
| 106 | 71 | 2 | 0.000 | Few | None | |
| 107 | 456 | 4 | 1.262 | 0.896–1.629 | Very few | Moderate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smallwood, K.S.; Smallwood, N.L. Breeding Density and Collision Mortality of Loggerhead Shrike (Lanius ludovicianus) in the Altamont Pass Wind Resource Area. Diversity 2021, 13, 540. https://doi.org/10.3390/d13110540
Smallwood KS, Smallwood NL. Breeding Density and Collision Mortality of Loggerhead Shrike (Lanius ludovicianus) in the Altamont Pass Wind Resource Area. Diversity. 2021; 13(11):540. https://doi.org/10.3390/d13110540
Chicago/Turabian StyleSmallwood, K. Shawn, and Noriko L. Smallwood. 2021. "Breeding Density and Collision Mortality of Loggerhead Shrike (Lanius ludovicianus) in the Altamont Pass Wind Resource Area" Diversity 13, no. 11: 540. https://doi.org/10.3390/d13110540