Tuber mesentericum and Tuber aestivum Truffles: New Insights Based on Morphological and Phylogenetic Analyses

 , , , , and
, , , , and
Abstract
:
1. Introduction
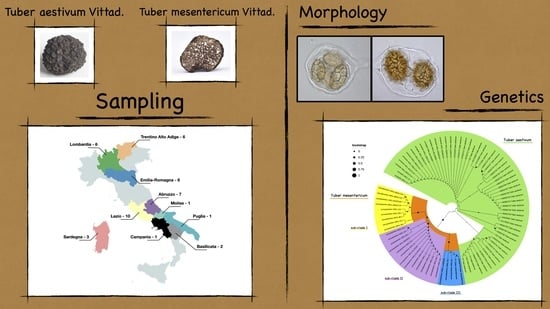
2. Materials and Methods
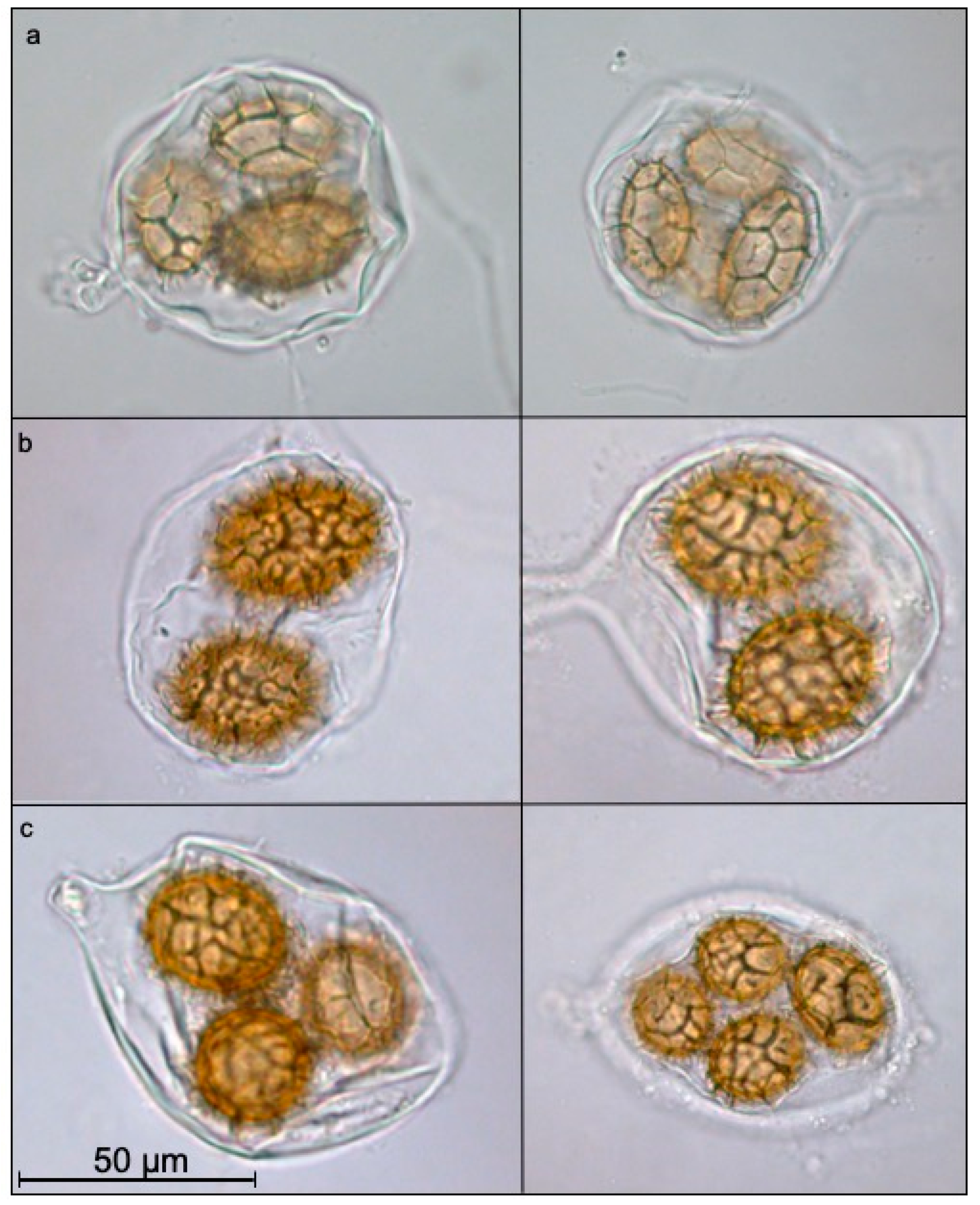
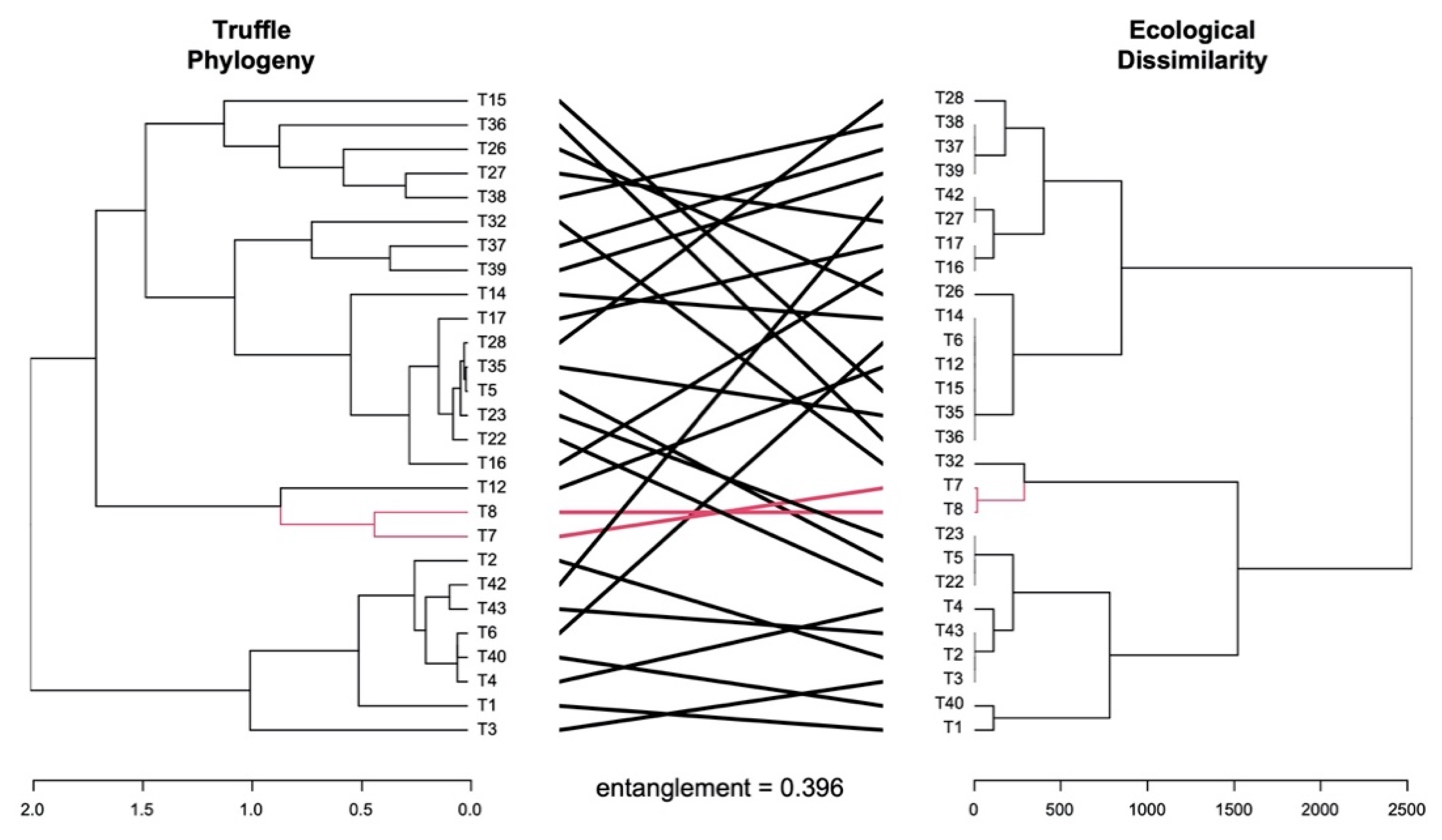
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stobbe, U.; Egli, S.; Tegel, W.; Peter, M.; Sproll, L.; Büntgen, U. Potential and limitations of Burgundy truffle cultivation. Appl. Microbiol. Biotechnol. 2013, 97, 5215–5224. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Frochot, H. Ecology and possibility of culture in Europe of the Burgundy truffle (Tuber uncinatum Chatin). Agric. Ecosyst. Environ. 1990, 28, 71–73. [Google Scholar] [CrossRef]
- Chevalier, G.; Sourzat, P. Soils and Techniques for Cultivating Tuber melanosporum and Tuber aestivum in Europe. In Edible Ectomycorrhizal Mushrooms; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; pp. 163–189. [Google Scholar]
- Venturella, G.; Altobelli, E.; Bernicchia, A.; Di Piazza, S.; Donnini, D.; Gargano, M.L.; Gorjòn, S.P.; Granito, V.M.; Lantieri, A.; Lunghini, D.; et al. Fungal biodiversity and in situ conservation in Italy. Plant Biosyst. 2011, 145, 950–957. [Google Scholar] [CrossRef]
- Boni, C.; Gregori, G.; Puxeddu, M.; Tocci, A. The Tuber aestivum Vitt. in Sardinia: Ecology and Prospects of Cultivation. Plant Biosyst. 1994, 128, 393. [Google Scholar]
- Wedén, C.; Pettersson, L.; Danell, E. Truffle cultivation in Sweden: Results from Quercus robur and Corylus avellana field trials on the island of Gotland. Scand. J. For. Res. 2009, 24, 37–53. [Google Scholar] [CrossRef]
- Benucci, G.M.N.; Bonito, G.; Baciarelli Falini, L.; Bencivenga, M. Mycorrhization of Pecan trees (Carya illinoinensis) with commercial truffle species: Tuber aestivum Vittad and Tuber borchii Vittad. Mycorrhiza 2012, 22, 383–392. [Google Scholar] [CrossRef]
- Donnini, D.; Gargano, M.L.; Perini, C.; Savino, E.; Murat, C.; Di Piazza, S.; Altobelli, E.; Salerni, E.; Rubini, A.; Rana, G.L.; et al. Wild and cultivated mushrooms as a model of sustainable development. Plant Biosyst. 2013, 147, 226–236. [Google Scholar] [CrossRef]
- Zambonelli, A.; Donnini, D.; Rana, G.L.; Fascetti, S.; Benucci, G.M.N.; Iotti, M.; Morte, A.; Khabar, L.; Bawadekji, A.; Piattoni, F.; et al. Hypogeous fungi in Mediterranean maquis, arid and semi-arid forests. Plant Biosyst. 2014, 148, 392–401. [Google Scholar] [CrossRef]
- Murat, C. Forty years of inoculating seedlings with truffle fungi: Past and future perspectives. Mycorrhiza 2015, 25, 77–81. [Google Scholar] [CrossRef]
- Molinier, V.; Peter, M.; Stobbe, U.; Egli, S. The Burgundy Truffle (Tuber aestivum syn. uncinatum): A Truffle Species with a Wide Habitat Range over Europe. In True Truffle (Tuber spp.) in the World; Springer: Berlin/Heidelberg, Germany, 2016; pp. 33–47. [Google Scholar]
- Hall, I.; Fitzpatrick, N.; Miros, P.; Zambonelli, A. Counter-season cultivation of truffles in the southern hemisphere: An update. Ital. J. Mycol. 2017, 46, 21–36. [Google Scholar]
- Marozzi, G.; Benucci, G.M.N.; Baciarelli Falini, L.; Albertini, E.; Donnini, D. Synthesis of Tuber mesentericum ectomycorrhizae with Quercus pubescens: A morphological review and DNA characterization. Sydowia 2018, 70, 81–88. [Google Scholar]
- Benucci, G.M.N.; Csorbai, A.G.; Baciarelli Falini, L.; Marozzi, G.; Suriano, E.; Sitta, N.; Donnini, D. Taxonomy, Biology and Ecology of Tuber macrosporum Vittad. and Tuber mesentericum Vittad. In True Truffle (Tuber spp.) in the World; Springer: Berlin/Heidelberg, Germany, 2016; pp. 69–86. [Google Scholar]
- Marozzi, G.; Baciarelli Falini, L.; Donnini, D. Aspetti biologici e genetici del tartufo mesenterico. In Tuber mesentericum Tartufo mesenterico; Associazione Tartufai del Friuli Venezia Giulia, Corvino edizioni: Udine, Italia, 2019; pp. 25–32. [Google Scholar]
- Pacioni, G.; Pomponi, G. Genotypic patterns of some Italian populations of the Tuber aestivum-T. mesentericum complex. Mycotaxon 1991, 42, 171–179. [Google Scholar]
- Montecchi, A.; Sarasini, M. Funghi Ipogei d’Europa; A.M.B. Fondazione: Trento, Italy, 2000. [Google Scholar]
- Ceruti, A.; Fontana, A.; Nosenzo, C. Le Specie Europee del Genere Tuber: Una Revisione Storica; Museo Reg. Scienze Nat.: Torino, Italy, 2003; Volume 37. [Google Scholar]
- Zhang, J.P.; Liu, P.G.; Chen, J. Tuber sinoaestivum sp. nov., an edible truffle from southwestern China. Mycotaxon 2012, 122, 73–82. [Google Scholar] [CrossRef]
- Mang, S.M.; Zotta, T.; Camele, I.; Racioppi, R.; D’Auria, M.; Rana, G.L. Morphological, physico-chemical and molecular investigations on Tuber bellonae from Basilicata-Italy. J. Anim. Plant Sci. 2017, 27, 528–541. [Google Scholar]
- Mello, A.; Cantisani, A.; Vizzini, A.; Bonfante, P. Genetic variability of Tuber uncinatum and its relatedness to other black truffles. Environ. Microbiol. 2002, 4, 584–594. [Google Scholar] [CrossRef]
- Gryndler, M.; Hršelová, H.; Soukupová, L.; Streiblová, E.; Valda, S.; Borovička, J.; Gryndlerová, H.; Gažo, J.; Miko, M. Detection of summer truffle (Tuber aestivum Vittad.) in ectomycorrhizae and in soil using specific primers. FEMS Microbiol. Lett. 2011, 318, 84–91. [Google Scholar] [CrossRef]
- Sica, M.; Gaudio, L.; Aceto, S. Genetic structure of Tuber mesentericum Vitt. based on polymorphisms at the ribosomal DNA ITS. Mycorrhiza 2007, 17, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Marozzi, G.; Sánchez, S.; Benucci, G.M.N.; Bonito, G.; Baciarelli Falini, L.; Albertini, E.; Donnini, D. Mycorrhization of pecan (Carya illinoinensis) with black truffles: Tuber melanosporum and Tuber brumale. Mycorrhiza 2017, 2, 303–309. [Google Scholar] [CrossRef]
- Bonito, G.; Smith, M.E.; Nowak, M.; Healy, R.A.; Guevara, G.; Cázares, E.; Kinoshita, A.; Nouhra, E.R.; Domínguez, L.S.; Tedersoo, L.; et al. Historical Biogeography and Diversification of Truffles in the Tuberaceae and Their Newly Identified Southern Hemisphere Sister Lineage. PLoS ONE 2013, 8, e52765. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) [Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. 1988. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 6 June 2020).
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Benucci, G.M.N.; Burnard, D.; Shepherd, L.D.; Bonito, G.; Munkacsi, A.B. Evidence for co-evolutionary history of early diverging Lycopodiaceae plants with fungi. Front. Microbiol. 2020, 10, 2944. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D. Vegan: Community Ecology Package; R Package Version 2.5-4. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 August 2020).
- Granetti, B.; De Angelis, A.; Materozzi, G. Umbria Terra di Tartufi; Umbriagraf: Terni, Italy, 2005. [Google Scholar]
- Breitenbach, J.; Kränzlin, F. Champignons de Suisse: Les Ascomycètes; Edition Mykologia: Lucerne, Switzerland, 1981; Volume 1. [Google Scholar]
- Read, N.D.; Beckett, A. Ascus and ascospore morphogenesis. Mycol. Res. 1996, 100, 1281–1314. [Google Scholar] [CrossRef]
- Zacchi, L.; Vaughan-Martini, A.; Angelini, P. Yeast distribution in a truffle-field ecosystem. Ann. Microbiol. 2003, 53, 275–282. [Google Scholar]
- Pacioni, G.; Leonardi, M.; Aimola, P.; Ragnelli, A.M.; Rubini, A.; Paolocci, F. Isolation and characterization of some mycelia inhabiting Tuber ascomata. Mycol. Res. 2007, 111, 1450–1460. [Google Scholar] [CrossRef]
- Rivera, C.S.; Blanco, D.; Oria, R.; Venturini, M.E. Diversity of culturable microorganisms and occurrence of Listeria monocytogenes and Salmonella spp. in Tuber aestivum and Tuber melanosporum ascocarps. Food Microbiol. 2010, 27, 286–293. [Google Scholar] [CrossRef]
- Stielow, B.; Menzel, W. Complete nucleotide sequence of TaV1, a novel totivirus isolated from a black truffle ascocarp (Tuber aestivum Vittad.). Arch. Virol. 2010, 155, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Gryndler, M.; Soukupová, L.; Hršelová, H.; Gryndlerová, H.; Borovička, J.; Streiblová, E.; Jansa, J. A quest for indigenous truffle helper prokaryotes. Environ. Microbiol. Rep. 2013, 5, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Benucci, G.M.N.; Bonito, G.M. The truffle microbiome: Species and geography effects on bacteria associated with fruiting bodies of hypogeous Pezizales. Microb. Ecol. 2016, 72, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.; Ascione, S.; Pacioni, G.; Cesare, P.; Pacioni, M.L.; Miranda, M. The challenge for identifying the fungi living inside mushrooms: The case of truffle inhabiting mycelia. Plant Biosyst. 2017, 152, 1002–1010. [Google Scholar] [CrossRef]
- Culleré, L.; Ferreira, V.; Chevret, B.; Venturini, M.E.; Sánchez-Gimeno, A.C.; Blanco, D. Characterisation of aroma active compounds in black truffles (Tuber melanosporum) and summer truffles (Tuber aestivum) by gas chromatography-olfactometry. Food Chem. 2010, 122, 300–306. [Google Scholar] [CrossRef]
- Splivallo, R.; Deveau, A.; Valdez, N.; Kirchhoff, N.; Frey-Klett, P.; Karlovsky, P. Bacteria associated with truffle-fruiting bodies contribute to truffle aroma. Environ. Microbiol. 2015, 17, 2647–2660. [Google Scholar] [CrossRef]
- Splivallo, R.; Vahdatzadeh, M.; Maciá-vicente, J.G.; Molinier, V.; Peter, M.; Egli, S.; Uroz, S.; Paolocci, F.; Deveau, A.; Huber, K.J. Orchard conditions and fruiting body characteristics drive the microbiome of the black truffle Tuber aestivum. Front. Microbiol. 2019, 10, 1437. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Wedén, C.; Danell, E.; Tibell, L. Species recognition in the truffle genus Tuber—The synonyms Tuber aestivum and Tuber uncinatum. Environ. Microbiol. 2005, 7, 1535–1546. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Riccioni, C.; Rubini, A.; Türkoğlu, A.; Belfiori, B.; Paolocci, F. Ribosomal DNA polymorphisms reveal genetic structure and a phylogeographic pattern in the Burgundy truffle Tuber aestivum Vittad. Mycologia 2019, 111, 26–39. [Google Scholar] [CrossRef]
- Rubini, A.; Paolocci, F.; Granetti, B.; Arcioni, S. Single step molecular characterization of morphologically similar black truffle species. FEMS Microbiol. Lett. 1998, 164, 7–12. [Google Scholar] [CrossRef]
- Rubini, A.; Paolocci, F.; Granetti, B.; Arcioni, S. Morphological characterization of molecular-typed Tuber magnatum ectomycorrhizae. Mycorrhiza 2001, 11, 179–185. [Google Scholar] [CrossRef]
- Seifert, K.A. Progress towards DNA barcoding of fungi. Mol. Ecol. Resour. 2009, 9, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Molinier, V.; van Tuinen, D.; Chevalier, G.; Gollotte, A.; Wipf, D.; Redecker, D. A multigene phylogeny demonstrates that Tuber aestivum and Tuber uncinatum are conspecific. Org. Divers. Evol. 2013, 13, 503–512. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID/Query Accession Number | Recognized Species (Ascocarp) | Collection Date (Year) | Collection Site (Italian Region) | Spore Morphology Classification | Consensus Taxa/GenBank Accession Number |
|---|---|---|---|---|---|
| T1/MH992040 | T. aestivum | 2013 | Molise | T. aestivum | T. aestivum/AY226026 |
| T2/MH992046 | T. aestivum | 2013 | Sardegna | T. aestivum | T. aestivum/AY226026 |
| T3/MH992052 | T. aestivum | 2013 | Sardegna | T. aestivum | T. aestivum/AY170350 |
| T4/MH992059 | T. aestivum | 2013 | Emilia Romagna | T. aestivum | T. aestivum/AY226022 |
| T5 */MH992063 | T. uncinatum | 2013 | Trentino Alto Adige | T. mesentericum | T. mesentericum/AF516775 |
| T6/MH992064 | T. uncinatum | 2014 | Lazio | T. aestivum | T. aestivum/AY226022 |
| T7/MH992065 | T. mesentericum | 2013 | Puglia | T. mesentericum | T. mesentericum/AF516778 |
| T8/MH992066 | T. mesentericum | 2013 | Lazio | T. mesentericum | T. mesentericum/AF516778 |
| T9 | T. mesentericum | 2013 | Lazio | T. mesentericum | |
| T10 | T. mesentericum | 2013 | Lazio | T. mesentericum | |
| T11 | T. mesentericum | 2013 | Lazio | T. mesentericum | |
| T12/MH992041 | T. mesentericum | 2013 | Abruzzo | Uncertain | T. mesentericum/AF516778 |
| T13 | T. mesentericum | 2013 | Abruzzo | T. mesentericum | |
| T14/MH992042 | T. mesentericum | 2013 | Abruzzo | T. mesentericum | T. mesentericum/AF516775 |
| T15/MH992043 | T. mesentericum | 2013 | Abruzzo | T. mesentericum | T. mesentericum/AF516769 |
| T16/MH992044 | T. mesentericum | 2013 | Lazio | T. mesentericum | T. mesentericum/AF516775 |
| T17/MH992045 | T. mesentericum | 2013 | Lazio | T. mesentericum | T. mesentericum/AF516775 |
| T18 | T. mesentericum | 2013 | Lazio | T. mesentericum | |
| T19 | T. mesentericum | 2013 | Lazio | T. mesentericum | |
| T20 | T. mesentericum | 2013 | Trentino Alto Adige | T. mesentericum | |
| T21 | T. mesentericum | 2013 | Trentino Alto Adige | T. mesentericum | |
| T22/MH992047 | T. mesentericum | 2013 | Trentino Alto Adige | T. mesentericum | T. mesentericum/AF516775 |
| T23/MH992048 | T. mesentericum | 2013 | Trentino Alto Adige | Uncertain | T. mesentericum/AF516775 |
| T24 | T. mesentericum | 2013 | Trentino Alto Adige | T. mesentericum | |
| T25 | T. mesentericum | 2013 | Basilicata | T. mesentericum | |
| T26/MH992049 | T. mesentericum | 2013 | Basilicata | T. mesentericum | T. mesentericum/AF516769 |
| T27/MH992050 | T. mesentericum | 2013 | Campania | T. mesentericum | T. mesentericum/AF516769 |
| T28/MH992051 | T. mesentericum | 2013 | Lombardia | T. mesentericum | T. mesentericum/AF516775 |
| T29 * | T. mesentericum | 2013 | Emilia Romagna | T. aestivum | |
| T30 * | T. mesentericum | 2013 | Emilia Romagna | T. aestivum | |
| T31 * | T. mesentericum | 2013 | Emilia Romagna | T. aestivum | |
| T32*/MH992053 | T. mesentericum | 2013 | Emilia Romagna | T. aestivum | T. mesentericum/AF516775 |
| T33 * | T. mesentericum | 2013 | Emilia Romagna | T. aestivum | |
| T34 | T. mesentericum | 2014 | Abruzzo | T. mesentericum | |
| T35/MH992054 | T. mesentericum | 2014 | Abruzzo | Uncertain | T. mesentericum/AF516775 |
| T36/MH992055 | T. mesentericum | 2014 | Abruzzo | T. mesentericum | T. mesentericum/AF516769 |
| T37/MH992056 | T. mesentericum | 2014 | Lombardia | T. mesentericum | T. mesentericum/AF516775 |
| T38/MH992057 | T. mesentericum | 2014 | Lombardia | T. mesentericum | T. mesentericum/AF516769 |
| T39/MH992058 | T. mesentericum | 2014 | Lombardia | T. mesentericum | T. mesentericum/AF516775 |
| T40 */MH992060 | T. mesentericum | 2014 | Lombardia | T. aestivum | T. aestivum/AY226022 |
| T41 * | T. mesentericum | 2014 | Lombardia | T. aestivum | |
| T42/MH992061 | T. uncinatum | 2013 | Lazio | T. aestivum | T. aestivum/AY226026 |
| T43/MH992062 | T. aestivum | 2013 | Sardegna | T. aestivum | T. aestivum/AY226026 |
| T. aestivum Clade | Sub-Clade_III * | Sub-Clade_I * | Sub-Clade_II * | Outgroup | |
|---|---|---|---|---|---|
| T. aestivum clade | - | 0.0077 | 0.0077 | 0.0088 | 0.0118 |
| sub-clade_III * | 0.0334 | - | 0.0055 | 0.0066 | 0.0117 |
| sub-clade_I * | 0.0338 | 0.0205 | - | 0.0063 | 0.0121 |
| sub-clade_II * | 0.0409 | 0.0274 | 0.0225 | - | 0.0130 |
| Outgroup | 0.0651 | 0.0625 | 0.0664 | 0.0708 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marozzi, G.; Benucci, G.M.N.; Suriano, E.; Sitta, N.; Raggi, L.; Lancioni, H.; Baciarelli Falini, L.; Albertini, E.; Donnini, D. Tuber mesentericum and Tuber aestivum Truffles: New Insights Based on Morphological and Phylogenetic Analyses. Diversity 2020, 12, 349. https://doi.org/10.3390/d12090349
Marozzi G, Benucci GMN, Suriano E, Sitta N, Raggi L, Lancioni H, Baciarelli Falini L, Albertini E, Donnini D. Tuber mesentericum and Tuber aestivum Truffles: New Insights Based on Morphological and Phylogenetic Analyses. Diversity. 2020; 12(9):349. https://doi.org/10.3390/d12090349
Chicago/Turabian StyleMarozzi, Giorgio, Gian Maria Niccolò Benucci, Edoardo Suriano, Nicola Sitta, Lorenzo Raggi, Hovirag Lancioni, Leonardo Baciarelli Falini, Emidio Albertini, and Domizia Donnini. 2020. "Tuber mesentericum and Tuber aestivum Truffles: New Insights Based on Morphological and Phylogenetic Analyses" Diversity 12, no. 9: 349. https://doi.org/10.3390/d12090349