Phycological Herbaria as a Useful Tool to Monitor Long-Term Changes of Macroalgae Diversity: Some Case Studies from the Mediterranean Sea



Abstract
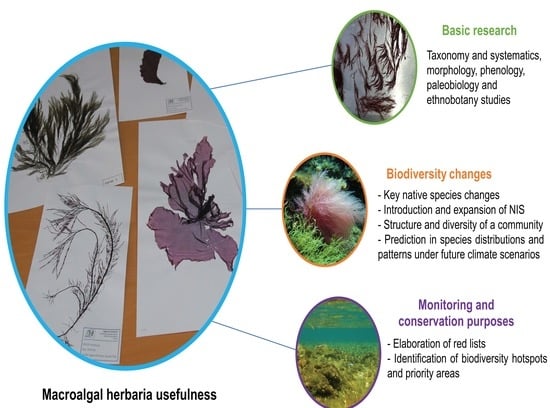
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Overview of European Phycological Collections
3.2. Case Studies from the Mediterranean Sea
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for Future Research. Mar. Poll. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudouresque, C.F. Marine biodiversity in the Mediterranean: Status of species, populations and communities. Sci. Rep. Port-Cros Natl. Park 2004, 20, 97–146. [Google Scholar]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Cinar, M.E.; Almogi- Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Patterns in introduction trends and pathways. Medit. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Zenetos, A.; Belchior, C.; Cardoso, A.C. Invading European Seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manag. 2013, 76, 64–74. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Corsini-Foka, M.; Morri, C.; Zenetos, A. Thirty years after: Dramatic change in the coastal marine ecosystems of Kos Island (Greece), 1981–2013. Mediterr. Mar. Sci. 2014, 15, 482–497. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–481. [Google Scholar] [CrossRef]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.; Pérez, T. Climate change effects on a miniature ocean: The highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef]
- Primack, R.B.; Miller-Rushing, A.J. The role of botanical gardens in climate change research. New Phytol. 2009, 182, 303–313. [Google Scholar] [CrossRef]
- Lavoie, C. Biological collections in an ever-changing world: Herbaria as tools for biogeographical and environmental studies. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 68–76. [Google Scholar] [CrossRef]
- Vellend, M.; Baeten, L.; Myers-Smith, I.H.; Elmendorf, S.C.; Beauséjour, R.; Brown, C.D.; De Frenne, P.; Verheyen, K.; Wipf, S. Global meta-analysis reveals no net change in local-scale plant biodiversity over time. Proc. Natl. Acad. Sci. USA 2013, 110, 19456–19459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meineke, E.K.; Davies, T.J.; Daru, B.H.; Davis, C.C. Biological collections for understanding biodiversity in the Anthropocene. Phil. Trans. R. Soc. B 2018, 374, 20170386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greve, M.; Lykke, A.M.; Fagg, C.W.; Gereau, R.E.; Lewis, G.P.; Marchant, R.; Marshall, A.R.; Ndayishimiye, J.; Bogaert, J.; Svenning, J.-C. Realising the potential of herbarium records for conservation biology. S. Afr. J. Bot. 2016, 105, 317–323. [Google Scholar] [CrossRef]
- Taylor, J.W.; Swann, E.C. DNA from herbarium specimens. In Ancient DNA; Hermann, B., Hummel, S., Eds.; Springer-Berlag: Berlin, Germany, 1994; pp. 167–181. [Google Scholar]
- Ruhling, A.; Tyler, G. Ecology of heavy metals—A regional and historical study. Bot. Not. 1969, 122, 248–259. [Google Scholar]
- Lang, P.L.M.; Willems, F.M.; Scheepens, J.F.; Burbano, H.A.; Bossdorf, O. Using herbaria to study global environmental change. New Phytol. 2019, 221, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Funk, V.A. The importance of Herbaria. Plant Sci. Bull. 2003, 49, 94–95. [Google Scholar]
- Johnson, K.G.; Brooks, S.J.; Fenberg, F.B.; Glover, A.G.; James, K.E.; Lister, A.M.; Michel, E.; Spencer, M.; Todd, J.A.; Valsami-Jones, E.; et al. Climate change and biosphere response: Unlocking the collections vault. BioScience 2011, 61, 147–153. [Google Scholar] [CrossRef]
- Krupnick, G.A.; Kress, W.J.; Wagner, W.L. Achieving Target 2 of the Global Strategy for Plant Conservation: Building a preliminary assessment of vascular plant species using data from herbarium specimens. Biodivers. Conserv. 2009, 18, 1459–1474. [Google Scholar] [CrossRef]
- Kricsfalusy, V.V.; Trevisan, N. Prioritizing regionally rare plant species for conservation using herbarium data. Biodivers. Conserv. 2014, 23, 39–61. [Google Scholar] [CrossRef]
- IUCN. An Introduction to the IUCN Red List of Ecosystems: The Categories and Criteria for Assessing Risks to Ecosystems; IUCN: Gland, Switzerland, 2016. [Google Scholar] [CrossRef]
- Davy, A.J. Museum specimens breathe life into plant conservation? Trends Ecol. Evol. 2005, 20, 285–286. [Google Scholar] [CrossRef]
- Seregin, A.P. Making the Russian flora visible: Fast digitisation of the Moscow University herbarium (MW) in 2015. Taxon 2016, 65, 203–209. [Google Scholar] [CrossRef]
- Armeli Minicante, A.; Birello, G.; Sigovini, M.; Minuzzo, T.; Perin, A.; Ceregato, A. Building a Natural and Cultural Heritage Repository for the Storage and Dissemination of Knowledge: The Algarium Veneticum and the Archivio di Studi Adriatici Case Study. J. Libr. Metadata 2017, 17, 1–15. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Anderson, J.G.T.; Bakker, J.D.; Billo, T.J.; Dunwiddie, P.W.; Groom, M.J.; Hampton, S.E.; Herman, S.G.; Levey, D.J.; Machnicki, N.J.; et al. Natural history’s place in science and society. BioScience 2014, 64, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.A.; Dalen, J.; Neill, K.F. Insights from natural history collections: Analysing the New Zealand macroalgal flora using herbarium data. PhytoKeys 2013, 30, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Thiers, B. The World’s Herbaria 2018: A Summary Report Based on Data from Index Herbariorum. 2019. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 15 May 2020).
- COST 49. Algae Directory: Algologists, Companies, Culture Collections and Herbaria in European Countries; Office for Official Publications of European Communities: Luxembourg, 1997; p. 235.
- Giaccone, T.; Catra, M.; Serio, D.; Giaccone, G. A review of Mediterranean macrophytobenthos collections present in Italy: A contribution to the Mediterranean Initiative on Taxonomy. Chem. Ecol. 2008, 24, 175–184. [Google Scholar] [CrossRef]
- Sales, M.; Ballesteros, E. Long-term comparison of algal assemblages dominated by Cystoseira crinita (Fucales, Heterokontophyta) from Cap Corse (Corsica, North Western Mediterranean). Eur. J. Phycol. 2010, 45, 404–412. [Google Scholar] [CrossRef]
- Sales, M.; Ballesteros, E.; Anderson, M.J.; Ivesa, I.; Cardona, E. Biogeographical patterns of algal communities in the Mediterranean Sea: Cystoseira crinita-dominated assemblages as a case study. J. Biogeogr. 2012, 39, 140–152. [Google Scholar] [CrossRef]
- Gianni, F.; Bartolini, F.; Airoldi, L.; Ballesteros, E.; Francour, P.; Guidetti, P.; Meinesz, A.; Thibaut, T.; Mangialajo, L. Conservation and restoration of marine forests in the Mediterranean Sea and the potential role of marine protected areas. Adv. Oceanogr. Limnol. 2013, 4, 83–101. [Google Scholar] [CrossRef]
- Mineur, F.; Arenas, F.; Assis, J.; Davies, A.J.; Engelen, A.H.; Fernandes, F.; Malta, E.-J.; Thibaut, T.; Van Nguyen, T.; Vaz-Pinto, F.; et al. European seaweeds under pressure: Consequences for communities and ecosystem functioning. J. Sea Res. 2015, 98, 91–108. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in French Riviera: The harbinger of future extinctions? Medit. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef] [Green Version]
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 2008, 358, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 2016, 181, 196–208. [Google Scholar] [CrossRef]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-term decline of the populations of Fucales (Cystoseira, Sargassum) in the Albères coast (north-western Mediterranean). Mar. Poll. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Sauvageau, C. A propos des Cystoseira de Banyuls et de Guéthary. Bull. Stat. Biol. Arcachon 1912, 14, 1–424. [Google Scholar]
- Feldmann, J. Les Algues Marines de ea Côte des Albères. I.-III; Cyanophycées, Chlorophycées et Phéophycées de la Côte des Albères; Imprimerie Wolf: Rouen, France, 1937; p. 197. [Google Scholar]
- Feldmann, J. Recherches sur la Végétation Marine de la Méditerranée. La Côte Des Albères; Imprimerie Wolf: Rouen, France, 1937; p. 339. [Google Scholar]
- Gros, C. Le genre Cystoseira sur côte des Albères. Répartition—Écologie—Morphogenèse. Ph.D. Thesis, Université Pierre et Marie Curie Paris VI, Paris, France, 1978. [Google Scholar]
- Guern, M. Embryologie de quelques espèces du genre Cystoseira Agardh 1821 (Fucales). Vie Milieu 1962, 13, 649–679. [Google Scholar]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The ups and downs of a canopy-forming seaweed over a span of more than one century. Sci. Rep. 2019, 9, 5250. [Google Scholar] [CrossRef] [PubMed]
- Agardh, J.G. Algae Maris Mediterranei et Adriatici, Observationes in Diagnosin specierum et Dispositionem Generum; Apud Fortin, Masson et Cie: Paris, France, 1842; p. 164. [Google Scholar]
- Raphélis, A. Liste des algues récoltées dans les environs de Cannes. Ann. Soc. Sci. Nat. Provence 1907, 1, 1–30. [Google Scholar]
- Thibaut, T.; Blanfumé, A.; Markovic, L.; Verlaque, M.; Boudouresque, C.-F.; Perret-Boudouresque, M.; Maćic, V.; Bottin, L. Unexpected abundance and long-term relative stability of the brown alga Cystoseira amentacea, hitherto regarded as a threatened species, in the north-western Mediterranean Sea. Mar. Pollut. Bull. 2014, 89, 305–323. [Google Scholar] [CrossRef]
- Verlaque, M.; Fritayre, P. Modifications des communautés algales méditerranéennes en présence de l’algue en-vahissante Caulerpa taxifolia (Vahl) C. Agardh. Oceanol. Acta 1994, 17, 659–672. [Google Scholar]
- Boudouresque, C.F.; Meinesz, A.; Ribera, M.A.; Ballesteros, E. Spread of the green alga Caulerpa taxifolia (Caulerpales, Chlorophyta) in the Mediterranean: Possible consequences of a major ecological event. Sci. Mar. 1995, 59 (Suppl. 1), 21–29. [Google Scholar]
- Piazzi, L.; Ceccherelli, G. Persistence of biological invasion effects: Recovery of macroalgal assemblages after removal of Caulerpa racemosa var. cylindracea. Estuar. Coast. Shelf Sci. 2006, 68, 455–461. [Google Scholar] [CrossRef]
- Hereu, B.; Capdevila, P.; Cebrian, E.; Díaz, D.; Garrabou, J.; Kerting, D.; Linares, C.; Navarro, L.; Pauner, O.; Teixido, N. Ecology and perturbations of Mediterranean deep-water algal communities: Linking population biology and community ecology for conservation. In Proceedings of the 5th Mediterranean Symposium on Marine Vegetation, Portorož, Slovenia, 27–28 October 2014; Langar, H., Bouafif, C., Ouerghi, A., Eds.; RAC/SPA publ.: Tunis, Tunisia, 2014; p. 20. [Google Scholar]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; p. 32. [Google Scholar] [CrossRef]
- ŽDuljević, A.; Peters, A.F.; Nikolić, V.; Antolić, B.; Despalatović, M.; Cvitković, I.; Isajlović, I.; Mihanović, H.; Matijević, S.; Shewring, D.M.; et al. The Mediterranean deep-water kelp Laminaria rodriguezii is an endangered species in the Adriatic Sea. Mar. Biol. 2016, 163, 69. [Google Scholar] [CrossRef] [Green Version]
- Ercegović, A. La Végétation des Algues sur les Fonds Péchereux de l’Adriatique. Ribarstveno-Bioloska Ekspedicija M/B Hvar Izvjesca 1948–1949. The M.V. Hvar Cruises-Researches into Fisheries Biology Reports 1948–1949; Institut za Oceanografiju i Ribarstvo: Split, Croatia, 1960; p. 32. [Google Scholar]
- Gamulin-Brida, H. Biocenoza muljevitog dna otvorenog srednjeg jadrana. (Biocoenose des fonds vaseux au large de l’Adriatique moyenne). Acta Adriat. 1965, 10, 1–27. [Google Scholar]
- Steen, F.; Aragay, J.; Zuljevic, A.; Verbruggen, H.; Mancuso, F.P.; Bunker, F.; Vitales, D.; Gómez Garreta, A.; De Clerck, O. Tracing the introduction history of the brown seaweed Dictyota cyanoloma (Phaeophyceae, Dictyotales) in Europe. Eur. J. Phycol. 2017, 52, 31–42. [Google Scholar] [CrossRef]
- Tronholm, A.; Steen, F.; Tyberghein, L.; Leliaert, F.; Verbruggen, H.; Siguan, M.A.R.; De Clerck, O. Species delimitation, taxonomy, and biogeography of Dictyota in Europe (Dictyotales, Phaeophyceae). J. Phycol. 2010, 46, 1301–1321. [Google Scholar] [CrossRef]
- Rull Lluch, J.R.; Ballesteros, E.; Barceló, M.C.; Gómez Garreta, A.; Ribera Siguan, M.A. Dictyota ciliolata Sonder ex Kützing (Phaeophyceae, Dictyotales) in the Mediterranean Sea. Cryptogamie Algol. 2007, 28, 89–97. [Google Scholar]
- Afonso-Carrillo, J. Lista actualizada de las Algas Marinas de las Islas Canarias, 2014; Elaborada para la Sociedad Española de Ficología (SEF): Las Palmas, Spain, 2014; p. 64. [Google Scholar]
- Bárbara, I.; De Clerk, O.; García-Redondo, V.; Peña, V.; García-Fernández, A.; Peteiro, C.; Sánchez, N. Nuevas citas y adiciones corológicas para la flora bentónica marina del Atlántico Ibérico. Acta Bot. Malac. 2015, 40, 191–198. [Google Scholar] [CrossRef]
- Bárbara, I.; Peña, V.; García-Redondo, V.; Díaz-Tapia, P.; García-Fernández, A.; Lugilde, J.; Piñeíro-Corbeira, C. Fragmentos taxonómicos, corológicos, nomenclaturales y fitocenológicos. 237. Nuevas citas y registros corológicos para la flora bentónica marina del Noroeste ibérico. Acta Bot. Malac. 2016, 41, 247–254. [Google Scholar] [CrossRef]
- Bárbara, I.; García-Redondo, V.; Díaz Tapia, P.; García-Fernández, A.; Piñeiro-Corbeira, C.; Peña, V.; Lugilde, J.; Cremades, J. Adiciones y correcciones a la flora bentónica marina del Atlántico ibérico norte. Acta Bot. Malac. 2019, 44, 1–10. [Google Scholar] [CrossRef]
- Gallardo, T.; Bárbara, I.; Afonso-Carrillo, J.; Bermejo, R.; Altamirano, M.; Gómez Garreta, A.; Barceló Martí, M.C.; Rull Lluch, J.; Ballesteros, E.; De la Rosa, J. Nueva lista crítica de las algas bentónicas marinas de España. A new checklist of benthic marine algae of Spain. Algas. Boletín Informativo Sociedad Española Ficología 2016, 51, 7–52. [Google Scholar]
- Bunker, F.; Brodie, J.A.; Maggs, C.A.; Bunker, A.R. Seaweeds of Britain and Ireland, 2nd ed.; Wild Nature Press: Plymouth, UK, 2017; pp. 1–312. [Google Scholar]
- Joher, S.; Ballesteros, E.; Cebrian, E.; Sánchez, N.; Rodríguez-Prieto, C. Deep-water macroalgal-dominated coastal detritic assemblages on the continental shelf off Mallorca and Menorca (Balearic Islands, Western Mediterranean). Bot. Mar. 2012, 55, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Joher, S.; Ballesteros, E.; Rodríguez-Prieto, C. Contribution to the study of deep coastal detritic bottoms: The algal communities of the continental shelf off the Balearic Islands, Western Mediterranean. Med. Mar. Sci. 2015, 16, 573–590. [Google Scholar] [CrossRef] [Green Version]
- Joher, S.; Ballesteros, E.; Rodríguez-Prieto, C. Macroalgal-dominated coastal detritic communities from the Western Mediterranean and the Northeastern Atlantic. Med. Mar. Sci. 2016, 17, 476–495. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Kim, M.S. Female reproductive structures define the novel genus, Nesoia (Halymeniaceae, Rhodophyta). Eur. J. Phycol. 2019, 54, 66–77. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, C.; De Clerck, O.; Huisman, J.M.; Lin, S.-M. Characterization of Nesoia latifolia (Halymeniaceae, Rhodophyta) from Europe with emphasis on cystocarp development and description of Nesoia mediterranea sp. nov. Phycologia 2019, 58, 393–404. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, C.; Afonso-Carrillo, J.; De Clerck, O.; Huisman, J.M.; Lin, S.-M. Systematic revision of the foliose Halymeniaceae (Halymeniales, Rhodophyta) from Europe, with the description of Halymenia ballesterosii sp. nov. from the Mediterranean Sea and Nesoia hommersandii from the Canary Islands. Eur. J. Phycol. 2020, 53, 520–536. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, C.; De Clerck, O.; Huisman, J.M.; Lin, S.-M. Systematics of the red algal genus Halymenia (Halymeniaceae, Rhodophyta): Characterization of the generitype H. floresii and description of Neofolia rosea gen. et sp. nov. Eur. J. Phycol. 2018, 53, 520–536. [Google Scholar] [CrossRef]
- Sáenz-Arroyo, A.; Roberts, C.M.; Torre, J.; Cariño-Olvera, M. Using fishers’ anecdotes, naturalists’ observations and grey literature to reassess marine species at risk: The case of the Gulf grouper in the Gulf of California. Mexico. Fish Fish. 2005, 6, 121–133. [Google Scholar] [CrossRef]
- Husa, V.; Steen, H.; Sjøtun, K. Historical changes in macroalgal communities in Hardangerfjord (Norway). Mar. Biol. Res. 2014, 10, 226–240. [Google Scholar] [CrossRef]
- Graham, C.H.; Ferrier, S.; Huettman, F.; Moritz, C.; Peterson, A.T. New developments in museum-based informatics and applications in biodiversity analysis. Trends Ecol. Evol. 2004, 19, 497–503. [Google Scholar] [CrossRef]
- Riera, R.; Sangil, C.; Sansón, M. Long-term herbarium data reveal the decline of a temperate-water algae at its southern range. Estuar. Coast. Shelf Sci. 2015, 165, 159–165. [Google Scholar] [CrossRef]
- Provan, J.; Booth, D.; Todd, N.P.; Beatty, G.E.; Maggs, C.A. Tracking biological invasions in space and time: Elucidating the invasive history of the green alga Codium fragile using old DNA. Diversity Distrib. 2008, 14, 343–354. [Google Scholar] [CrossRef]
- Schaffer, H.B.; Fisher, R.N.; Davidson, C. The role of natural history collections in documenting species declines. Trends Ecol. Evol. 1998, 13, 27–30. [Google Scholar] [CrossRef]
- Tingley, M.W.; Beissinger, S.R. Detecting range shifts from historical species occurrences: New perspectives on old data. Trends Ecol. Evol. 2009, 24, 625–633. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Acronym | Institution | City | Number of Specimens of Marine Macroalgae |
|---|---|---|---|
| PC | Muséum National d’Histoire Naturelle | Paris, France | 500,000 * |
| C | University of Copenhagen | Copenhagen, Denmark | 260,000 |
| BM | Natural History Museum | Kensington, United Kingdom | 205,000 |
| W | Naturhistorisches Museum Wien | Wien, Austria | 200,000 * |
| L | Naturalis Biodiversity Center | Leiden, The Netherlands | 148,000 |
| LD | Lund University | Lund, Sweden | 102,000 |
| GENT | Ghent University | Ghent, Belgium | 42,653 |
| HCOM | Centre d’Océanologie de Marseille - University of Aix-Marseille II | Marseilles, France | 40,000 |
| SANT | Universidad de Santiago de Compostela | Santiago de Compostela, Spain | 30,218 |
| FI | Natural history Museum | Firenze, Italy | 30,000 |
| TCD | Trinity College | Dublin, Ireland | 25,000 |
| RO | Sapienza Università di Roma | Rome, Italy | 23,000 |
| HGI | Universitat de Girona | Girona, Spain | 20,600 |
| GB | Gothenburg University | Gothenburg, Sweden | 20,000 |
| MCVE | Museo di Storia Naturale di Venezia | Venice, Italy | 18,500 |
| PAD | Università degli Studi di Padova | Padova, Italy | 15,000 |
| MS | Università di Messina | Messina, Italy | 8000 |
| BCN | Universidad de Barcelona | Barcelona, Spain | 7500 |
| CAT | Università di Catania | Catania, Italy | 6193 |
| MGC | Universidad de Málaga | Malaga, Spain | 5496 |
| TSB | Università de Triestre | Trieste, Italy | 5169–5426 |
| MA | Real Jardín Botánico de Madrid | Madrid, Spain | 4105 |
| ZA and ZAHO | University of Zagreb | Zagreb, Croatia | 4000 |
| PAL | Orto Botanico dell’Università degli Studi di Palermo | Palermo, Italy | 3856 |
| --- | Herbarium J.J. Rodríguez y Femenías, Ateneu de Menorca | Menorca, Spain | 2775 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, A.M.; Armeli Minicante, S.; Rodríguez-Prieto, C. Phycological Herbaria as a Useful Tool to Monitor Long-Term Changes of Macroalgae Diversity: Some Case Studies from the Mediterranean Sea. Diversity 2020, 12, 309. https://doi.org/10.3390/d12080309
Mannino AM, Armeli Minicante S, Rodríguez-Prieto C. Phycological Herbaria as a Useful Tool to Monitor Long-Term Changes of Macroalgae Diversity: Some Case Studies from the Mediterranean Sea. Diversity. 2020; 12(8):309. https://doi.org/10.3390/d12080309
Chicago/Turabian StyleMannino, Anna Maria, Simona Armeli Minicante, and Conxi Rodríguez-Prieto. 2020. "Phycological Herbaria as a Useful Tool to Monitor Long-Term Changes of Macroalgae Diversity: Some Case Studies from the Mediterranean Sea" Diversity 12, no. 8: 309. https://doi.org/10.3390/d12080309