4.1. Survival and Habitat Selection
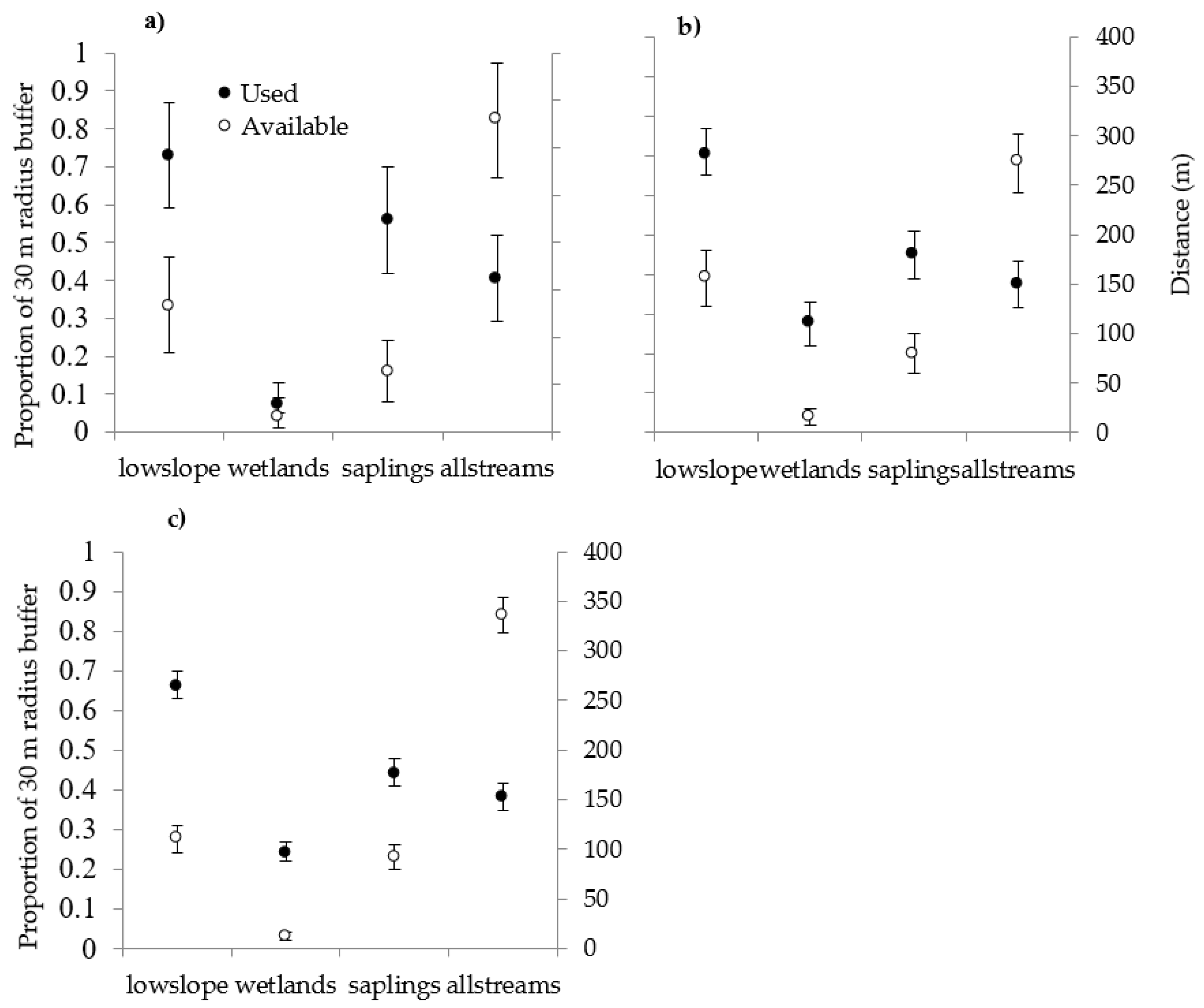
Rusty blackbirds in our study selected (at a 30 m radius scale) a combination of habitat conditions for nesting and post-fledging, including shallow wetlands and low slope with softwood/mixed-wood sapling stands. This was the first study to assess the importance of distance to 1st to 6th order streams and found that this feature was selected for both nesting and post-fledging. Availability of streams in addition to wetlands may increase foraging opportunities because streams and wetlands have different availability of prey types throughout the season [
31,
49,
50,
51,
52]. Early nestling development may be synchronized with the availability of small items typical of streams, such as Ephemeroptera, Plecoptera, and Trichoptera larvae, while dependent fledglings may require larger Odonata larvae, which are more abundant in wetlands (P. Wohner unpublished data). A diverse network of interconnected channels and impoundments, such as those created by beavers, may provide ideal foraging opportunities for rusty blackbirds [
52].
We found that while habitat selection was similar between nest sites and post-fledging locations at the 30 m scale, the stages had one important difference: wetland cover. Likewise, in a study that included the same drainages, rusty blackbirds did not select wetlands for nesting at the 5 m scale [
29]. At the 500 m scale, rusty blackbirds were reported more likely to select nesting habitat with increases in wetland cover in New Hampshire [
29]. However, the confidence interval for the wetland estimate ranged from −2 to 13, overlapping 1, and is therefore inconclusive [
29]. We concur that rusty blackbirds select nest sites independent of wetland cover at the 5, 30, or 500 m scale. However, wetlands are important to adults and fledglings at the 30 m scale. Concealment from predators may be more important than proximity to foraging areas during nesting and early post-fledging, while foraging opportunities quickly become a priority after fledging [
9].
It is likely that proximity to wetlands is important for nest-site selection at a larger scale than has been studied thus far, i.e., >500 m [
53]. Rusty blackbirds are highly mobile, traveling up to 2 km between wetlands and nesting areas, and have large home ranges (3.8–172.8 ha in Maine, equivalent to a circle with radius 35–741 m [
54]). It is possible that cues for selection of nesting and foraging habitat are decoupled at different spatial scales, and rusty blackbirds select wetlands at a 1–5 km scale for foraging, and nesting habitat at a much smaller scale (0.05–0.5 km [
53]). We propose that future nesting habitat selection studies that use multiple spatial scales including the 1–5 km scale would help to determine the density and size of wetlands important for rusty blackbirds at the home range scale and provide better predictive ability for rusty blackbirds [
55].
Finding the appropriate scale could have important consequences for research and management [
55]. For example, many surveys for nesting rusty blackbirds have been wetland-centric and typically only detect songbirds within 100–200 m of a point. In our study, 35 percent of nests were located >200 m from the nearest wetland. If rusty blackbird pairs forage at multiple wetlands and streams and nest in relatively distant uplands, wetland-based surveys likely underestimate occupancy and may result in biases [
56]. The low detectability documented in other studies that only surveyed wetlands could be a consequence of survey design [
28]. Other researchers have noted that point count-based surveys are not well suited for estimating the abundance or habitat use of breeding rusty blackbirds, even when broadcast calls are used [
19].
Our nest success estimates (0.48) are well within the range reported by other rusty blackbird researchers (0.21–0.75; [
27,
28,
29]) and are much higher than those reported for red-winged, yellow-headed, and Brewer’s blackbirds (0.30–0.39; [
30,
57]). Regenerating clear-cuts were suspected ecological traps for nesting in rusty blackbirds in Maine [
28]. However, we found evidence that regenerating clear-cuts support successful nesting by rusty blackbirds. Although we did not explicitly test stand origin, nest success did not decrease with increases in softwood sapling stands, 92% of which originated from even-aged forest management. This result is consistent with other research that found rusty blackbird nest success was independent of recent harvesting [
26]. We agree with other researchers that nest success is unlikely to be limiting rusty blackbird populations [
24,
26].
As in other New England rusty blackbird breeding studies, where nest selection was assessed at 5 and 500 m radius scales [
27,
29], rusty blackbirds in our study selected young (5–15 years old) or stunted softwood/mixed wood stands at the 30 m radius scale for nesting. Many of the New England nesting stands were created by even-aged management and are composed of regenerating softwoods. For example, 75% of nests in Maine [
28], 88% in Maine and New Hampshire [
29], and 92% in this study were located in young, even-aged softwood stands. While rusty blackbirds seem to have an affinity for regenerating clear-cuts at the 5, 30, and 500 m scales, there may be a threshold for increasing proportions of regenerating softwood stands at larger scales than 500 m. Blackpoll warblers (
Setophaga striata) were found to be positively associated with large proportions of clear-cut at the 115 m scale [
6]. However, at the 1250 m scale, they were positively associated with clear-cuts only when < 5% of cover was clear-cut, above which the relationship was negative [
6]. Thus, while even-aged forest management may be beneficial for rusty blackbirds at smaller scales, e.g., 5, 30, and 500 m radius, researchers should consider a scale larger than 500 m for future studies.
We observed low mortality rates for rusty blackbird fledglings in the first week after fledging (3%) compared to those reported for other songbirds, e.g., 21–81% for ovenbird [
2], 11% for worm-eating warbler [
8], and 90% for rose-breasted grosbeak (
Pheucticus ludovicianus) fledglings [
7]. All observed fledgling mortality took place in the first four days of a yellow warbler (
Setophaga petechi) study [
58]. Our high rusty blackbird fledgling survival contrasts with virtually every other songbird post-fledging study. We suggest that dense regenerating softwood stands approximately 5–15 years post-harvest, may afford < 1-week-old fledglings protection from potential predators while they are most vulnerable. Fledglings of songbirds that nest in mature hardwood stands with relatively open understories have sparse protective cover before moving to early successional habitats with dense vegetation [
1,
2,
6,
12]. Our overall fledgling survival during the 60-day post-fledging period was also relatively high (0.49) and is likely not an overwhelming factor contributing to population decline. However, we did not study survivorship after fledglings were completely independent from adults, nor during migration when young birds could experience substantial mortality [
9]. Populations of fledgling barn swallows (
Hirundo rustica erythrogaster) were found to be limited by the pre-migration phase [
9].
While few studies have estimated adult survival during either nesting or post-fledging, adult barn swallow survival over 60 days was 0.92, SE = 0.11 for males and females together [
59], compared to our estimates of 0.89 for females and 0.79 for males over the same time frame. Rusty blackbird male survival may truly be relatively low during the post-fledging period. However, the confidence intervals around our estimates are wide (0.54–1), likely due to small sample sizes. Transmitter harnesses may reduce survival, resulting in a survival estimate that is lower than actual survival in the general population. We regularly observed rusty blackbirds picking at their harnesses which could distract birds and expose them to higher predation. We have also recaptured blackbirds with transmitters embedded in their skin. Transmitter harnesses have been found to affect survival in sensitive species such as pileated woodpeckers (
Dryocopus pileatus; [
60]). Why harnesses would have a disproportionate effect on males over females is unknown but could be due to morphological differences between the sexes. High mortality of adult rusty blackbird males during the breeding season seems unusual, but if true, could itself be a key factor in population decline. Male survival during the breeding season warrants future study.
4.2. Recommendations
Nest success and adult and fledgling survival were not affected by any of the habitat variables we analyzed, which suggests that something other than breeding ground habitat may be limiting rusty blackbird populations. Thus, we expect that current forest management practices continue to create suitable landscapes for successful nesting and post-breeding survival. Our study adds support to a study in Nova Scotia, that found rusty blackbird habitat remains relatively abundant and well-distributed and is often located in wet lowlands which is a climate-resilient topographic landform [
61]. Targeted habitat management for rusty blackbirds is likely to be unnecessary in many areas due to the species’ use of regenerating softwood stands that are created by a variety of harvesting practices. We do recommend that harvest plans ensure the availability of at least one softwood stand 5 to 15 years post-harvest within 300 m of streams and shallow wetlands over time. In the Acadian forest, on ownerships where wildlife habitat is the focus of forest management, prioritizing harvests on poorly drained sites where trees grow more slowly could provide rusty blackbird nesting habitat for longer periods of time (≥40 years) [
62]).
To aid in finding high priority areas for research or conservation, we recommend overlaying the regression equation from our top models for nests and fledglings in the raster calculator in ARCMAP (i.e., for nests: −0.87–5.2 (distance to streams) + 0.01 (prop slope) + 0.02 (prop softwood/mixed-wood sapling stands). The nests and fledgling models had high goodness of fit, and together, could identify rusty blackbird habitat at the landscape level. We expect our regression equations will be applicable in the southeastern portion of the rusty blackbird’s range in Acadian Forest, i.e., New Brunswick, Nova Scotia, northern New England, and the Adirondacks.
Finally, we hope that rusty blackbird researchers will move from traditional survey protocols [
63,
64] like presence-absence surveys to protocols such as those used for western yellow-billed cuckoos (
Coccyzus americanus occidentalis), which also have large home ranges (16–91 ha) [
65,
66]. These surveys use broadcast calls every 100 m along transects in appropriate habitat [
67,
68]. (Spring surveys to locate pairs prior to nest searching for intensive research should avoid using broadcast calls, however). Future rusty blackbird research on the breeding ground should incorporate a landscape perspective and include multiple habitat types, including but not limited to dense young softwood stands, streams, seepages, and wetlands. Studying the species at a much larger scale than previously (e.g., >500m), may shed new light on wetland requirements during nesting.




{kind=link}
{kind=link}