A Review of Galaxias (Galaxiidae) Fossils from the Southern Hemisphere

 ,
,
Abstract
:
1. Introduction
“It was formerly believed that the same fresh-water species never existed on two continents distant from each other. But Dr. Günther has lately shown that the Galaxias attenuatus [sic; now Galaxias maculatus] inhabits Tasmania, New Zealand, the Falkland Islands, and the mainland of South America. This is a wonderful case, and probably indicates dispersal from an Antarctic centre during a former warm period. This case, however, is rendered in some degree less surprising by the species of this genus having the power of crossing by some unknown means considerable spaces of open ocean: thus there is one species common to New Zealand and to the Auckland Islands, though separated by a distance of about 230 miles. On the same continent fresh-water fish often range widely, and as if capriciously; for in two adjoining river-systems some of the species may be the same, and some wholly different.”([1], pp. 374–375)
2. History of Discovery of Galaxiidae Fossils in New Zealand

3. Material Studied, Fossil Localities and Ages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Fossil Record Number | Species | Preservation | Age | References |
|---|---|---|---|---|---|
| Foulden Maar | I43/f8503 | G. effusus | articulated larvae, juveniles and adults | early Miocene (23 Ma) | [14,16,30] |
| Bannockburn Inlet, Bannockburn Fm. | F41/f217 | Galaxias n. sp. ‘Bannockburn Inlet’ | isolated cranial bones, one partial specimen | early Miocene (19–16 Ma) | [14,17,31] |
| Manuherikia River, Bannockburn Fm. | H41/f089 H41/f090 H41/f096 | G. angustiventris G. brevicauda G. papilionis Galaxias sp. | otoliths; otoliths; otoliths; otoliths | early Miocene (19–16 Ma) | [18] |
| Manuherikia River and Vinegar Hill, Bannockburn Fm. | H41/f0112 H41/f0113 | G. bobmcdowalli G. parvirostris | otoliths;otoliths | early Miocene (19–16 Ma) | [18] |
| Lauder, Bannockburn Fm. | G41/f9508 | G. tabidus | otoliths | early Miocene (19–16 Ma) | [18] |
| Hindon Maar | I44/f0392 | Galaxias n. sp. ‘Hindon’ | larvae, juveniles, adults | middle Miocene (14 Ma) | [29] |
| Frasers Gully, Kaikorai Leaf Beds | I44/f145 | G. kaikorai Whitley | articulated adults | middle Miocene (13–11 Ma) | [14,16,23] |
| Kilmog Hill, ‘Kaikorai Leaf Beds’ | I44/f015 | Galaxias n. sp. ‘Kilmog Hill’ | nearly entire fishes | middle Miocene (13–11 Ma) | [14,32] |
3.1. Foulden Maar (I43/f8503), Earliest Miocene, 23 Ma


3.2. Lake Manuherikia Localities
3.2.1. Bannockburn Inlet (F41/217), Early Miocene, 19–16 Ma
3.2.2. Manuherikia River, Near St Bathans (H41/f089, H41/f096, H41/f090), Early Miocene, 19–16 Ma
3.2.3. Vinegar Hill, Near St Bathans (H41/f112, H41/f113), Early Miocene, 19–16 Ma
3.2.4. Lauder, Central Otago (G41/f9508), Early Miocene, 19–16 Ma

3.3. Hindon Maar (I44/f0392) Middle Miocene, c. 14 Ma
3.4. Frasers Gully, Kaikorai Valley, Dunedin (I44/f0145) Middle Miocene, 13–11 Ma

3.5. Kilmog Hill (I44/f015) Middle Miocene, 13–11 Ma
4. Systematic Paleontology
4.1. Assignment of the Fossils to Galaxias
4.2. Macrofossils
4.2.1. Galaxias kaikorai Whitley 1956 (Figure 6A–E)

4.2.2. Galaxias effusus Lee, McDowall and Lindqvist 2007 (Figure 2 and Figure 3)
4.2.3. Galaxias n. sp. ‘Hindon’ (Figure 5)
4.2.4. Galaxias n. sp. ‘Kilmog Hill’ (Figure 7)
4.2.5. Galaxias n. sp. ‘Bannockburn Inlet’ (Figure 4)
4.3. Otolith-Based Species
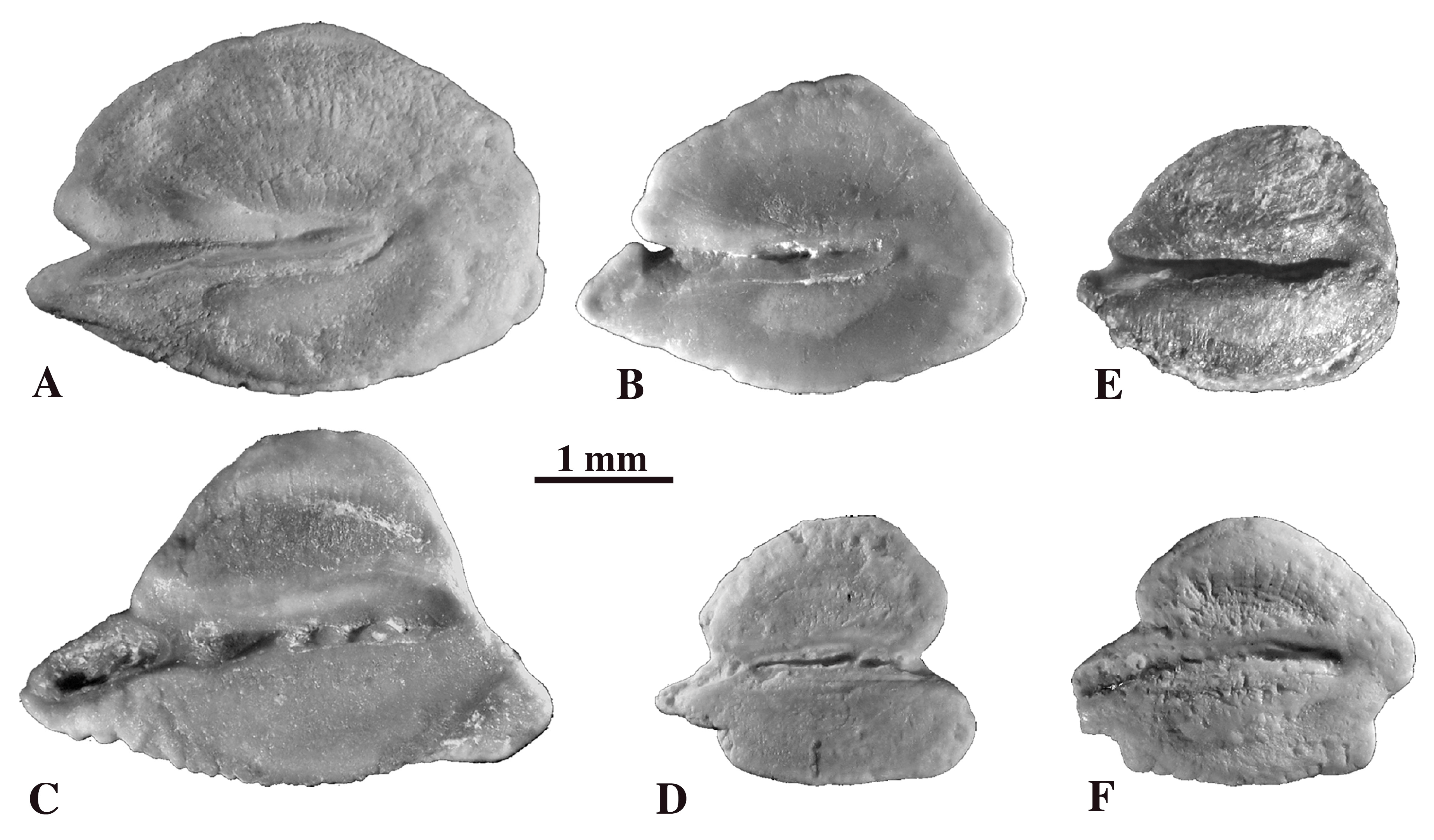
4.3.1. Galaxias angustiventris Schwarzhans 2012 (Figure 8A)

4.3.2. Galaxias brevicauda Schwarzhans 2012 (Figure 8B)
4.3.3. Galaxias bobmcdowalli Schwarzhans 2012 (Figure 8C)
4.3.4. Galaxias papilionis Schwarzhans 2012 (Figure 8D)
4.3.5. Galaxias parvirostris Schwarzhans 2012 (Figure 8E)
4.3.6. Galaxias tabidus Schwarzhans 2012 (Figure 8F)

4.3.7. Galaxias sp.
4.4. Coprolites
5. Discussion
5.1. Diversity of Miocene Galaxiids
5.2. Paleolake Ecosystems and Diadromy
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Origin of Species by Means of Natural Selection; 6th (reprint) ed.; J.M. Dent & Sons Ltd.: London, UK, 1872. [Google Scholar]
- McDowall, R.M. Generalized tracks and dispersal in biogeography. Syst. Zool. 1978, 27, 88–104. [Google Scholar] [CrossRef]
- Allibone, R.M.; Wallis, G.P. Genetic variation and diadromy in some native New Zealand galaxiids (Teleostei: Galaxiidae). Biol. J. Linn. Soc. 1993, 50, 19–33. [Google Scholar] [CrossRef]
- Allibone, R.M.; Crowl, T.A.; Holmes, J.M.; King, T.M.; McDowall, R.M.; Townsend, C.R.; Wallis, G.P. Isozyme analysis of Galaxias species (Teleostei: Galaxiidae) from the Taieri River, South Island, New Zealand: A species complex revealed. Biol. J. Linn. Soc. 1996, 57, 107–127. [Google Scholar] [CrossRef] [Green Version]
- Burridge, C.P.; McDowall, R.M.; Craw, D.; Wilson, M.V.H.; Waters, J.M. Marine dispersal as a pre-requisite for Gondwanan vicariance among elements of the galaxiid fish fauna. J. Biogeogr. 2012, 39, 306–321. [Google Scholar] [CrossRef]
- Waters, J.M.; Dijkstra, L.H.; Wallis, G.P. Biogeography of a southern hemisphere freshwater fish: How important is marine dispersal? Mol. Ecol. 2000, 9, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.M.; Lopez, J.A.; Wallis, G.P. Molecular phylogenetics and biogeography of galaxiid fishes (Osteichthyes: Galaxiidae): Dispersal, vicariance, and the position of Lepidogalaxias salamandroides. Syst. Biol. 2000, 49, 777–795. [Google Scholar] [CrossRef] [Green Version]
- Burridge, C.P.; Craw, D.; Waters, J.M. River capture, range expansion, and cladogenesis: The genetic signature of freshwater vicariance. Evolution 2006, 60, 1038–1049. [Google Scholar] [CrossRef]
- Waters, J.M.; Craw, D.; Youngson, J.H.; Wallis, G.P. Genes meet geology: Fish phylogeographic pattern reflects ancient, rather than modern, drainage connections. Evolution 2001, 55, 1844–1851. [Google Scholar] [CrossRef]
- Waters, J.M.; Rowe, D.L.; Burridge, C.P.; Wallis, G.P. Gene trees versus species trees: Reassessing life-history evolution in a freshwater fish radiation. Syst. Biol. 2010, 59, 504–517. [Google Scholar] [CrossRef] [Green Version]
- Slater, G.J.; Harmon, L.J. Unifying fossils and phylogenies for comparative analyses of diversification and trait evolution. Methods Ecol. Evol. 2013, 4, 699–702. [Google Scholar] [CrossRef]
- Wallis, G.P.; Trewick, S.A. New Zealand phylogeography: Evolution on a small continent. Mol. Ecol. 2009, 18, 3548–3580. [Google Scholar] [CrossRef] [PubMed]
- Wallis, G.P.; Jorge, F. Going under down under? Lineage ages argue for extensive survival of the Oligocene marine transgression on Zealandia. Mol. Ecol. 2018, 27, 4368–4396. [Google Scholar] [CrossRef]
- Lee, D.E.; McDowall, R.M.; Lindqvist, J.K. Galaxias fossils from Miocene lake deposits, Otago, New Zealand: The earliest records of the Southern Hemisphere family Galaxiidae (Teleostei). J. Roy. Soc. N. Z. 2007, 37, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Lindqvist, J.K.; Mildenhall, D.C.; Bannister, J.M.; Kaulfuss, U. Paleobotany, palynology and sedimentology of Late Cretaceous-Miocene sequences in Otago and Southland. In Field Trip Guides, Geosciences 09 Conference, Oamaru, New Zealand; Turnbull, I.M., Ed.; Geological Society of New Zealand Miscellaneous Publication: Wellington, New Zealand, 2009; Volume 128B, pp. FT12−1–FT12−39. [Google Scholar]
- McDowall, R.M. Notes on some Galaxias fossils from the Pliocene of New Zealand. J. Roy. Soc. N. Z. 1976, 6, 17–22. [Google Scholar] [CrossRef]
- McDowall, R.M.; Pole, M. A large galaxiid fossil (Teleostei) from the Miocene of Central Otago, New Zealand. J. Roy. Soc. N. Z. 1997, 27, 193–198. [Google Scholar] [CrossRef]
- Schwarzhans, W.; Scofield, R.P.; Tennyson, A.J.D.; Worthy, J.P.; Worthy, T.H. Fish remains, mostly otoliths, from the non-marine early Miocene of Otago, New Zealand. Acta Palaeontol. Pol. 2012, 57, 319–350. [Google Scholar] [CrossRef] [Green Version]
- McDowall, R.M. New Zealand freshwater fishes. A natural history and guide; Heinemann Reed: Auckland, New Zealand, 1990. [Google Scholar]
- McDowall, R.M. The Reed Field Guide to New Zealand Freshwater Fishes; Reed Publishing: Auckland, New Zealand, 2000. [Google Scholar]
- Mortimer, N.; Campbell, H. Zealandia: Our Continent Revealed; Penguin and GNS: Auckland, New Zealand; Lower Hutt, New Zealand, 2014. [Google Scholar]
- Gibbs, G. Ghosts of Gondwana; revised ed.; Potton & Burton: Nelson, New Zealand, 2016. [Google Scholar]
- Oliver, W.R.B. The Tertiary flora of the Kaikorai Valley, Otago, New Zealand. Trans. Proc. Roy. Soc. N. Z. 1937, 66, 284–304. [Google Scholar]
- Stokell, G. The systematic arrangement of the New Zealand Galaxiidae. Part I. Generic and sub-generic classification. Trans. Roy. Soc. N. Z. 1945, 75, 124–137. [Google Scholar]
- Campbell, J.D. Casuarinaceae, Fagaceae, and other plant megafossils from Kaikorai Leaf Beds (Miocene) Kaikorai Valley, Dunedin, New Zealand. N. Z. J. Bot. 1985, 23, 311–320. [Google Scholar] [CrossRef]
- Whitley, G.P. New fishes from Australia and New Zealand. Proc. Roy. Zool. Soc. N. S. W. 1956, 1954–55, 34–38. [Google Scholar]
- Travis, C. Geology of the Slip Hill area east of Middlemarch. Master’s Thesis, University of Otago, Dunedin, New Zealand, 1965. [Google Scholar]
- Kaulfuss, U. Geology and Paleontology of Foulden Maar, Otago, New Zealand. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 2013. [Google Scholar]
- Kaulfuss, U.; Lee, D.E.; Wartho, J.-A.; Bowie, E.; Lindqvist, J.K.; Conran, J.G.; Bannister, J.M.; Mildenhall, D.C.; Kennedy, E.M.; Gorman, A.R. Geology and palaeontology of the Hindon Maar Complex: A Miocene terrestrial fossil Lagerstätte in southern New Zealand. Palaeogeogr. Palaeocl. 2018, 500, 52–68. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Kaulfuss, U.; Conran, J.G.; Bannister, J.M.; Lindqvist, J.K. Biodiversity and palaeoecology of Foulden Maar: An Early Miocene Konservat-Lagerstätte deposit in southern New Zealand. Alcheringa 2016, 40, 525–541. [Google Scholar] [CrossRef]
- McDowall, R.M. Relationships of the galaxioid fishes with further discussion of salmoniform relationships. Copeia 1969, 4, 796–824. [Google Scholar] [CrossRef]
- Harvey, B.E. Dunedin non-marine deposits. Bachelor’s Thesis, University of Otago, Dunedin, New Zealand, 1978. [Google Scholar]
- Lindqvist, J.K.; Lee, D.E. High-frequency paleoclimate signals from Foulden Maar, Waipiata Volcanic Field, southern New Zealand: An Early Miocene varved lacustrine diatomite deposit. Sediment Geol. 2009, 222, 98–110. [Google Scholar] [CrossRef]
- Mildenhall, D.C.; Kennedy, E.M.; Lee, D.E.; Kaulfuss, U.; Bannister, J.M.; Fox, B.; Conran, J.G. Palynology of the early Miocene Foulden Maar, Otago, New Zealand: Diversity following destruction. Rev. Palaeobot. Palynol. 2014, 204, 27–42. [Google Scholar] [CrossRef]
- Kaulfuss, U. Crater stratigraphy and the post-eruptive evolution of Foulden Maar, southern New Zealand. N. Z. J. Geol. Geophys. 2017, 60, 410–432. [Google Scholar] [CrossRef]
- Harper, M.A.; Van De Vijver, B.; Kaulfuss, U.; Lee, D.E. Resolving the confusion between two fossil freshwater diatoms from Otago, New Zealand: Encyonema jordanii and Encyonema jordaniforme (Cymbellaceae, Bacillariophyta). Phytotaxa 2019, 394, 231–243. [Google Scholar] [CrossRef]
- Chapman, A.; Morgan, D.L.; Gill, H.S. Description of the larval development of Galaxias maculatus in landlocked lentic and lotic systems in Western Australia. N. Z. J. Mar. Fresh. 2009, 43, 563–569. [Google Scholar] [CrossRef]
- Douglas, B.J. Lignite resources of Central Otago (Manuherikia Group of Central Otago, New Zealand: Stratigraphy, Depositional Environments, Lignite Resource Assessment and Exploration Models); New Zealand Energy Research and Development Committee Publication P104; University of Auckland: Auckland, New Zealand, 1986. [Google Scholar]
- Qu, Q.; Haitina, T.; Zhu, M.; Ahlberg, P.E. New genomic and fossil data illuminate the origin of enamel. Nature 2015, 526, 108–112. [Google Scholar] [CrossRef]
- Grange, L.I. Diatomite: Principal New Zealand occurrences and uses. New Zeal. J. Sci. Tech. 1930, 12, 94–99. [Google Scholar]
- Cervellini, P.M.; Battini, M.A.; Cussac, V.E. Ontogenetic shifts in the diet of Galaxias maculatus (Galaxiidae) and Odontesthes microlepidotus (Atherinidae). Environ. Biol. Fish. 1993, 36, 283–290. [Google Scholar] [CrossRef]
- Waters, J.M.; Craw, D.; Burridge, C.P.; Kennedy, M.; King, T.M.; Wallis, G.P. Within-river genetic connectivity patterns reflect contrasting geomorphology. J. Biogeogr. 2015, 42, 2452–2460. [Google Scholar] [CrossRef]
- Waters, J.M.; Wallis, G.P.; Burridge, C.P.; Craw, D. Geology shapes biogeography: Quaternary river-capture explains New Zealand’s biologically ‘composite’ Taieri River. Quat. Sci. Rev. 2015, 120, 47–56. [Google Scholar] [CrossRef]
- Prebble, J.G.; Reichgelt, T.; Mildenhall, D.C.; Greenwood, D.R.; Raine, J.I.; Kennedy, E.M.; Seebeck, H.C. Terrestrial climate evolution in the Southwest Pacific over the past 30 million years. Earth Planet Sci. Lett. 2017, 459, 136–144. [Google Scholar] [CrossRef]
- McDowall, R.M.; Kennedy, E.M.; Lindqvist, J.K.; Lee, D.E.; Alloway, M.R.; Gregory, M.R. Probable Gobiomorphus fossils from the Miocene and Pleistocene of New Zealand. J. Roy. Soc. N. Z. 2006, 36, 97–109. [Google Scholar] [CrossRef]
- McDowall, R.M.; Lee, D.E. Probable perciform fish scales from a Miocene freshwater lake deposit, Central Otago, New Zealand. J. Roy. Soc. N. Z. 2005, 35, 339–344. [Google Scholar] [CrossRef]
- Nolf, D. The Diversity of Fish Otoliths, Past and Present; Operational Directorate “Earth and History of Life” of the Royal Belgian Institute of Natural Sciences: Brussels, Belgium, 2013. [Google Scholar]
- Seehausen, O.; Wagner, C.E. Speciation in freshwater fishes. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 621–651. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaulfuss, U.; Lee, D.E.; Robinson, J.H.; Wallis, G.P.; Schwarzhans, W.W. A Review of Galaxias (Galaxiidae) Fossils from the Southern Hemisphere. Diversity 2020, 12, 208. https://doi.org/10.3390/d12050208
Kaulfuss U, Lee DE, Robinson JH, Wallis GP, Schwarzhans WW. A Review of Galaxias (Galaxiidae) Fossils from the Southern Hemisphere. Diversity. 2020; 12(5):208. https://doi.org/10.3390/d12050208
Chicago/Turabian StyleKaulfuss, Uwe, Daphne E. Lee, Jeffrey H. Robinson, Graham P. Wallis, and Werner W. Schwarzhans. 2020. "A Review of Galaxias (Galaxiidae) Fossils from the Southern Hemisphere" Diversity 12, no. 5: 208. https://doi.org/10.3390/d12050208