Paramecium Diversity and a New Member of the Paramecium aurelia Species Complex Described from Mexico


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling and Maintenance of Paramecium Strains
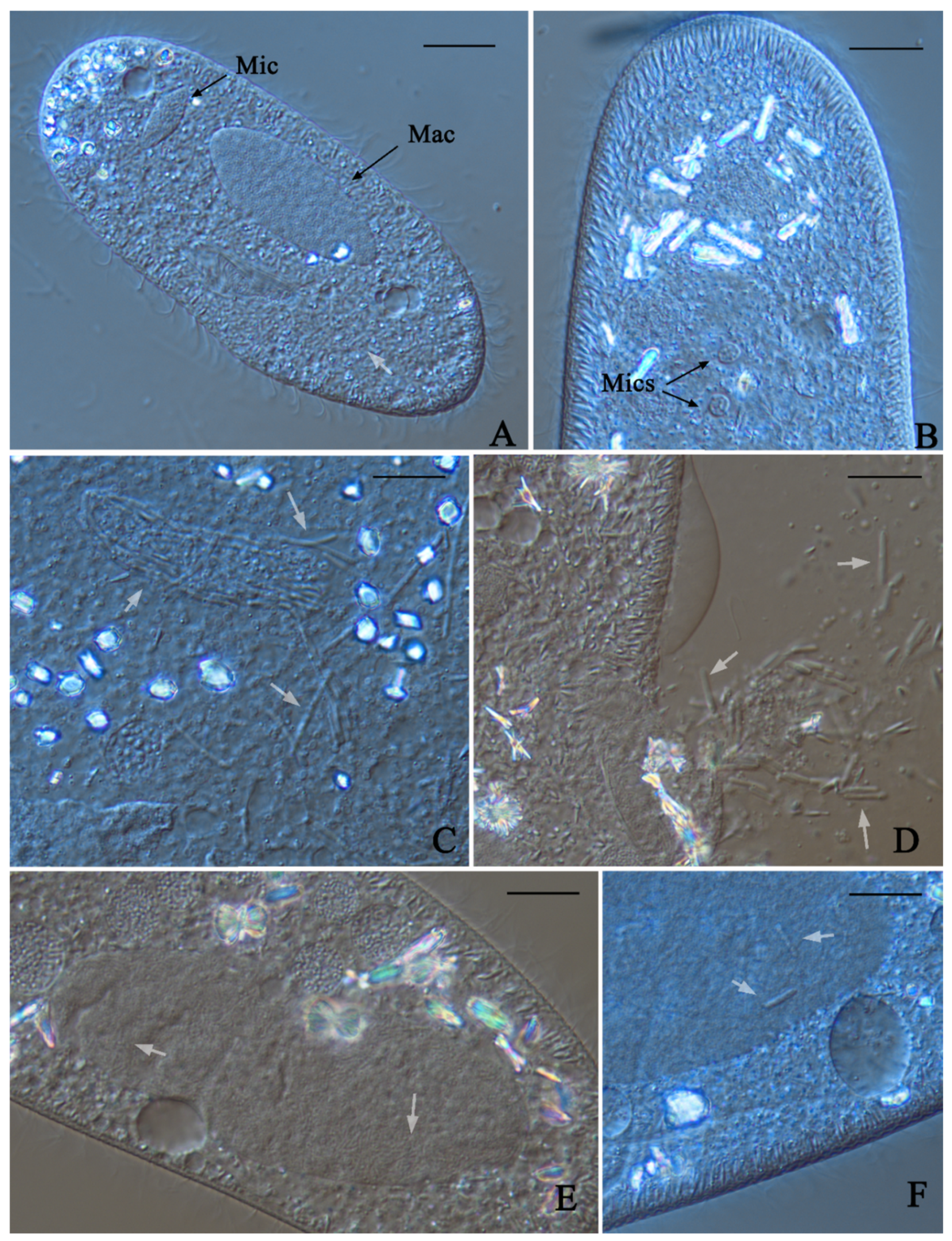
2.2. DIC Microscopy and Stainings
2.3. Molecular Identification of Paramecium Strains and Bacterial Symbionts
2.4. Molecular Phylogenetic Analysis
2.5. Mating Tests
3. Results
3.1. Diversity of Paramecium and Its Bacterial Symbionts Revealed by Extensive Sampling in Several Regions of Mexico
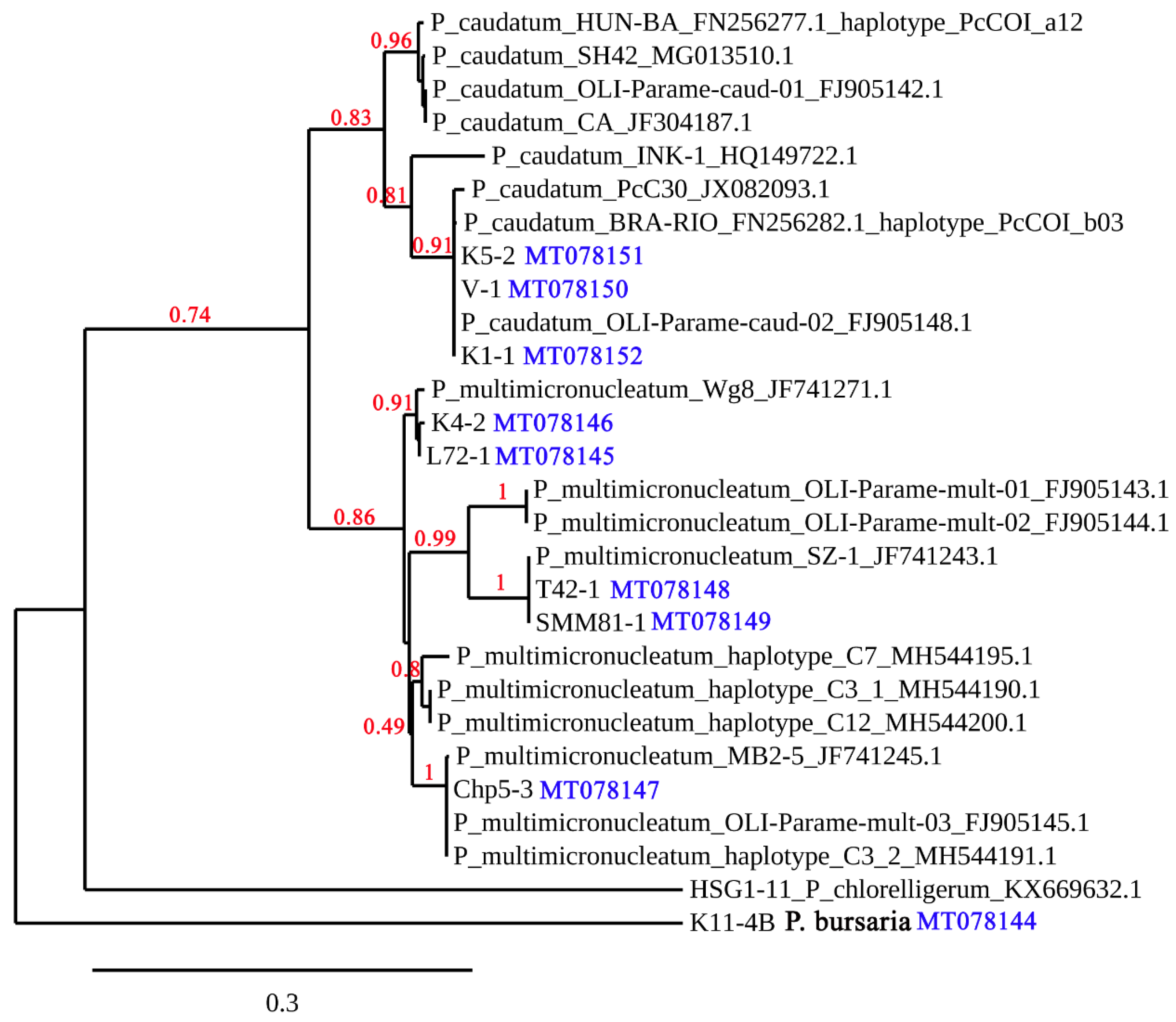
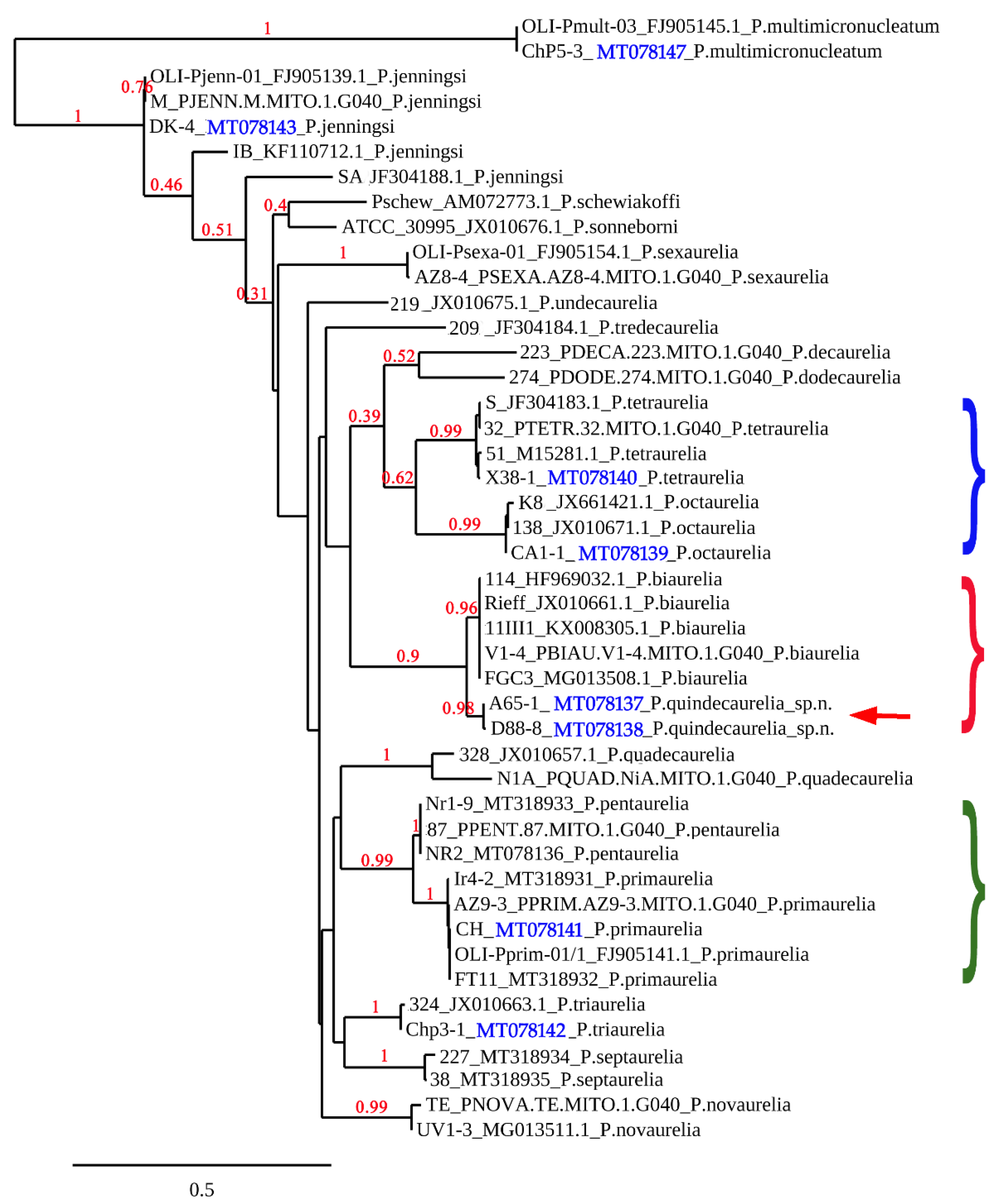
3.2. Phylogenetic Analysis of the Collected Strains
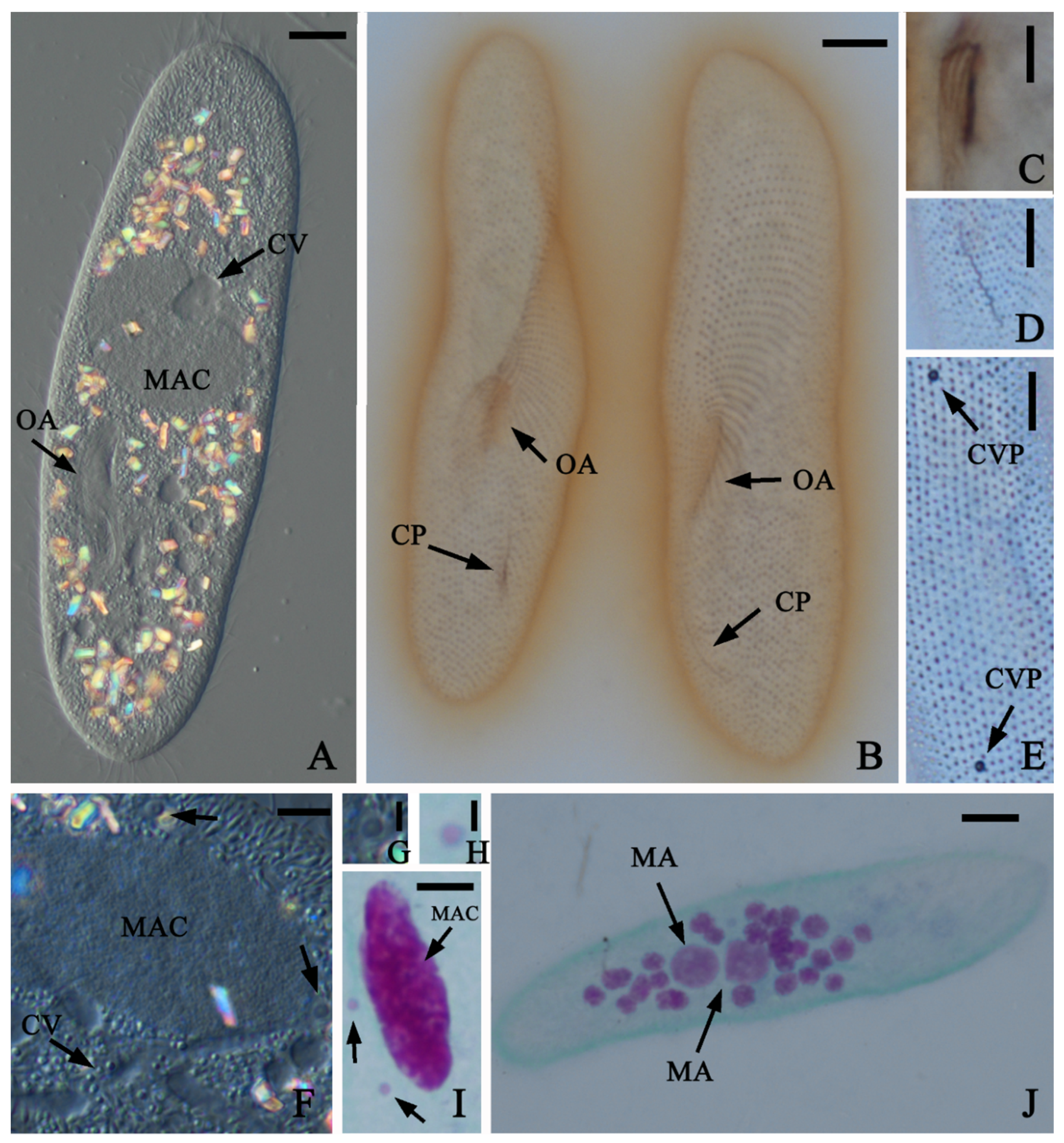
3.3. New Species of the Paramecium Aurelia Complex, P. Quindecaurelia n. sp.
4. Discussion
4.1. Overview of Paramecium Diversity Revealed in Mexico
4.2. Paramecium Quindecaurelia n. sp. and a Species Concept in Paramecium
4.3. Paramecium Quindecaurelia n. sp. Taxonomic Summary
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fokin, S.I. Paramecium genus: Biodiversity, some morphological features and the key to the main morphospecies discrimination. Protistology 2010, 6, 227–235. [Google Scholar]
- Kreutz, M.; Stoeck, T.; Foissner, W. Morphological and molecular characterization of Paramecium (Viridoparamecium nov. subgen.) chlorelligerum Kahl 1935 (Ciliophora). J. Eukaryot. Microbiol. 2012, 59, 548–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenek, S.; Berendonk, T.U.; Fokin, S.I. New Paramecium (Ciliophora, Oligohymenophorea) congeners shape our view on its biodiversity. Org. Divers. Evol. 2015, 15, 215–233. [Google Scholar] [CrossRef]
- Sonneborn, T.M. The Paramecium aurelia complex of 14 sibling species. Trans. Am. Microsc. Soc. 1975, 94, 155–178. [Google Scholar] [CrossRef]
- Greczek-Stachura, M.; Potekhin, A.; Przyboś, E.; Rautian, M.; Skoblo, I.; Tarcz, S. Identification of Paramecium bursaria syngens through molecular markers-comparative analysis of three loci in the nuclear and mitochondrial DNA. Protist 2012, 163, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Tasneem, F.; Shakoori, F.R.; Ilyas, M.; Shahzad, N.; Potekhin, A.; Shakoori, A.R. Genetic diversity of Paramecium species on the basis of multiple loci analysis and ITS secondary structure models. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wichterman, R. The Biology of Paramecium, 2nd ed.; Springer: New York, NY, USA, 1986; pp. 1–599. [Google Scholar]
- Strüder-Kypke, M.C.; Wright, A.-D.G.; Fokin, S.; Lynn, D.H. Phylogenetic relationships of the genus Paramecium inferred from small subunit rRNA gene sequences. Mol. Phylogenet. Evol. 2000, 14, 122–130. [Google Scholar] [CrossRef]
- Strüder-Kypke, M.C.; Lynn, D.H. Comparative analysis of the mitochondrial cytochrome c oxidase subunit I (COI) gene in ciliates (Alveolata, Ciliophora) and evaluation of its suitability as a biodiversity marker. Syst. Biodivers. 2010, 8, 131–148. [Google Scholar] [CrossRef]
- Catania, F.; Würmser, F.; Potekhin, A.A.; Przyboś, E.; Lynch, M. Genetic diversity in the Paramecium aurelia species complex. Mol. Biol. Evol. 2009, 26, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Tarcz, S.; Rautian, M.; Potekhin, A.; Sawka, N.; Beliavskaya, A.; Kiselev, A.; Nekrasova, I.; Przyboś, E. Paramecium putrinum (Ciliophora, Protozoa): The first insight into the variation of two DNA fragments—Molecular support for the existence of cryptic species. Mol. Phylogenet. Evol. 2014, 73, 140–145. [Google Scholar] [CrossRef]
- Yi, Z.; Strüder-Kypke, M.; Hu, X.; Lin, X.; Song, W. Sampling strategies for improving tree accuracy and phylogenetic analyses: A case study in ciliate protists, with notes on the genus Paramecium. Mol. Phylogenet. Evol. 2014, 71, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Barth, D.; Krenek, S.; Fokin, S.I.; Berendonk, T.U. Intraspecific genetic variation in Paramecium revealed by mitochondrial cytochrome c oxidase I sequences. J. Eukaryot. Microbiol. 2006, 53, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gentekaki, E.; Yi, Z.; Lin, X. Genetic differentiation of the mitochondrial cytochrome oxidase C subunit I gene in genus Paramecium (Protista, Ciliophora). PLoS ONE 2013, 8, e77044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanzoni, O.; Fokin, S.I.; Lebedeva, N.; Migunova, A.; Petroni, G.; Potekhin, A. Rare freshwater ciliate Paramecium chlorelligerum Kahl, 1935 and its macronuclear symbiotic bacterium “Candidatus Holospora parva”. PLoS ONE 2016, 11, e0167928. [Google Scholar] [CrossRef]
- Lu, X.; Gentekaki, E.; Xu, Y.; Huang, L.; Li, Y.; Lu, X.; Zhao, Y.; Lin, X.; Yi, Z. Intra-population genetic diversity and its effects on outlining genetic diversity of ciliate populations: Using Paramecium multimicronucleatum as an example. Eur. J. Protistol. 2019, 67, 142–150. [Google Scholar] [CrossRef]
- Fokin, S.I.; Stoeck, T.; Schmidt, H.J. Rediscovery of Paramecium nephridiatum Gelei, 1925 and its characteristics. J. Eukaryot. Microbiol. 1999, 46, 416–426. [Google Scholar] [CrossRef]
- Fokin, S.I.; Stoeck, T.; Schmidt, H.J. Paramecium duboscqui Chatton et Brachon, 1933. Distribution, ecology and taxonomy. Europ. J. Protistol. 1999, 35, 161–167. [Google Scholar] [CrossRef]
- Sonneborn, T.M. Paramecium aurelia. In Handbook of Genetics; King, R., Ed.; Plenum: New York, NY, USA, 1974; Volume 2, pp. 469–594. [Google Scholar]
- Foissner, W. An update of basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. Int. J. Syst. Evol. Microbiol. 2014, 64, 271–292. [Google Scholar] [CrossRef] [Green Version]
- Vannini, C.; Rosati, G.; Verni, F.; Petroni, G. Identification of the bacterial endosymbionts of the marine ciliate Euplotes magnicirratus (Ciliophora, Hypotrichia) and proposal of ‘Candidatus Devosia euplotis’. Int. J. Syst. Evol. Microbiol. 2004, 54, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- ParameciumDB. Available online: https://paramecium.i2bc.paris-saclay.fr (accessed on 15 May 2020).
- Arnaiz, O.; Meyer, E.; Sperling, L. ParameciumDB 2019: Integrating genomic data across the genus for functional and evolutionary biology. Nucleic Acids Res. 2020, 48, D599–D605. [Google Scholar] [CrossRef]
- MUSCLE. Available online: http://www.ebi.ac.uk/Tools/msa/muscle/ (accessed on 15 May 2020).
- Phylogeny.fr Robust Phylogenetic Analysis for the Non-Specialist. Available online: http://www.phylogeny.fr (accessed on 15 May 2020).
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beisson, J.; Bétermier, M.; Bré, M.H.; Cohen, J.; Duharcourt, S.; Duret, L.; Kung, C.; Malinsky, S.; Meyer, E.; Preer, J.R., Jr.; et al. Maintaining clonal Paramecium tetraurelia cell lines of controlled age through daily reisolation. Cold Spring Harb. Protoc. 2010. [Google Scholar] [CrossRef] [PubMed]
- Sawka-Gądek, N.; Potekhin, A.; Singh, D.P.; Grevtseva, I.; Arnaiz, O.; Sperling, L.; Tarcz, S.; Nekrasova, I.; Meyer, E. Evolutionary plasticity of mating-type determination mechanisms in Paramecium aurelia sibling species. Genome Biol. Evol. Under review.
- Schrallhammer, M.; Castelli, M.; Petroni, G. Phylogenetic relationships among endosymbiotic R-body producer: Bacteria providing their host the killer trait. Syst. Appl. Microbiol. 2018, 41, 213–220. [Google Scholar] [CrossRef]
- Orias, E.; Singh, D.P.; Meyer, E. Genetics and epigenetics of mating type determination in Paramecium and Tetrahymena. Annu. Rev. Microbiol. 2017, 71, 133–156. [Google Scholar] [CrossRef]
- Tarcz, S.; Potekhin, A.; Rautian, M.; Przyboś, E. Variation in ribosomal and mitochondrial DNA sequences demonstrates the existence of intraspecific groups in Paramecium multimicronucleatum (Ciliophora, Oligohymenophorea). Mol. Phylogenet. Evol. 2012, 63, 500–509. [Google Scholar] [CrossRef]
- Fokin, S.I.; Chivilev, S.M. Paramecium. Morphometric analysis and taxonomy. Acta Protozool. 2000, 39, 1–14. [Google Scholar]
- Aladro-Lubel, M.A.; Martínez-Murillo, M.E.; Mayén-Estrada, R. Lista de los ciliados bentónicos salobres y marinos registrados en México. An. Inst. Biol. Univ. Nal. Autón. Méx. Ser. Zool. 1987, 58, 403–448. [Google Scholar]
- Aladro-Lubel, M.A.; Martínez-Murillo, M.E.; Mayén-Estrada, R. Manual de Ciliados Psamófilos Marinos y Salobres de México (Cuadernos 9); Instituto de Biología, Universidad Nacional Autónoma de México: México City, Mexico, 1990; pp. 1–172. [Google Scholar]
- Aladro-Lubel, M.A.; Mayén-Estrada, R.; Reyes-Santos, M. Listados Faunísticos de México XI. Registro Actualizado de Ciliados; Instituto de Biología, Universidad Nacional Autónoma de México: México City, Mexico, 2006; pp. 1–97. [Google Scholar]
- Ancona-Méndez, L.M. Dinámica de Población de Paramecium Caudatum en Diferentes Excretas. Bachelor’s Thesis, Universidad de Yucatán, Mérida, Mexico, 1979. [Google Scholar]
- Coats, D.; Clamp, J.C. Ciliated protists (Ciliophora) of the Gulf of Mexico. In Gulf of Mexico. Origins, Waters and Biota: Biodiversity; Felder, D.L., Camp, D.K., Eds.; Texas A&M Press: College Station, TX, USA, 2009; Volume 1, pp. 57–79. [Google Scholar]
- Cordero-Ramírez, J.D.; López-Rivera, R.; Calderón-Vázquez, C.L.; Figueroa-López, A.M.; Martínez-Álvarez, J.C.; Leyva-Madrigal, K.Y.; Cervantes-Gámez, R.G.; Maldonado-Mendoza, I.E. Microorganismos asociados a la rizosfera de jitomate en un agroecosistema del valle de Guasave, Sinaloa, México. Rev. Mex. Biodiv. 2012, 83, 712–730. [Google Scholar] [CrossRef]
- Gallegos-Neyra, E.; Sánchez-Rodríguez, M.R.; Calderón-Vega, A.; Lugo-Vázquez, A. Diversidad de protozoos. La diversidad biológica del Estado de México. Estudio de estado. In Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad; Ceballos, G., List, R., Garduño, G., López-Cano, R., Muñozcano-Quintanar, M.J., Collado, E., Eivin-San Román, J., Eds.; Secretaría de Medio Ambiente, Gobierno del Estado de México, Biblioteca Mexiquense del Bicentenario: México City, Mexico, 2009; pp. 83–88. [Google Scholar]
- Hernández-Anaya, M. Ciliados de Una Laguneta de los Médanos Cercanos al Puerto de Veracruz. Bachelor’s Thesis, Universidad Nacional Autónoma de México, México City, Mexico, 1981. [Google Scholar]
- López-Ochoterena, E.; Roure-Cané, E. Lista taxonómica comentada de protozoarios de vida libre de México. Rev. Soc. Mex. Hist. Nat. 1970, 31, 23–68. [Google Scholar]
- Madrazo-Garibay, M.; López-Ochoterena, E. Segunda lista taxonómica comentada de protozoarios de vida libre de México. Rev. Lat-amer. Microbiol. 1982, 24, 281–295. [Google Scholar]
- Méndez-Sánchez, D. Estudio Taxonómico de Ciliados de Vida Libre (Alveolata: Ciliophora) en Cuerpos de Agua Dulce de Dos Regiones Biogeográficas de México. Master’s Thesis, Universidad Nacional Autónoma de México, México City, Mexico, 2017. [Google Scholar]
- Méndez-Sánchez, D.; Sánchez-Nava, P.; Mayén-Estrada, R. Free-living ciliates from a perturbed marsh in Central Mexico: Some notes about taxonomy and ecology. Protistology 2018, 12, 173–184. [Google Scholar] [CrossRef]
- Sigala-Regalado, I. Registro Protozoológico en Cinco Biotopos de la Cueva de Los Riscos, Querétaro, México. Bachelor’s Thesis, Universidad Nacional Autónoma de México, México City, Mexico, 2008. [Google Scholar]
- Sigala-Regalado, I. Diversidad de Protozoos Ciliados en Cinco Biotopos de Cuevas en México. Master’s Thesis, Universidad Nacional Autónoma de México, México City, Mexico, 2011. [Google Scholar]
- Przyboś, E.; Tarcz, S. Global molecular variation of Paramecium jenningsi complex (Ciliophora, Protista): A starting point for further, detailed biogeography surveys. Syst. Biodivers. 2019, 17, 527–539. [Google Scholar] [CrossRef]
- Allen, S.L.; Rushford, D.L.; Nerad, T.A.; Lau, E.T. Intraspecies variability in the esterases and acidphosphatases of Paramecium jenningsi and Paramecium multimicronucleatum: Assignment of unidentified paramecia, comparison with the Paramecium aurelia complex. J. Protozool. 1983, 30, 155–163. [Google Scholar] [CrossRef]
- Przyboś, E.; Tarcz, S. Paramecium jenningsi complex: Existence of three cryptic species confirmed by multi-locus analysis and strain crosses. Syst. Biodivers. 2016, 14, 140–154. [Google Scholar] [CrossRef]
- Przyboś, E.; Tarcz, S.; Dusi, E. New Paramecium quadecaurelia strains (P. aurelia spp. complex, Ciliophora) identified by molecular markers (rDNA and mtDNA). Eur. J. Protistol. 2013, 49, 477–486. [Google Scholar] [CrossRef]
- Aufderheide, K.J.; Daggett, P.-M.; Nerad, T.A. Paramecium sonneborni n. sp., a new member of the Paramecium aurelia species complex. J. Protozool. 1983, 30, 128–131. [Google Scholar] [CrossRef]
- Yusa, A. The morphology and morphogenesis of the buccal organelles in Paramecium with particular reference to their systematic significance. J. Protozool. 1957, 4, 128–142. [Google Scholar] [CrossRef]
- Dedukh, D.; Litvinchuk, J.; Svinin, A.; Litvinchuk, S.; Rosanov, J.; Krasikova, A. Variation in hybridogenetic hybrid emergence between populations of water frogs from the Pelophylax esculentus complex. PLoS ONE 2019, 14, e0224759. [Google Scholar] [CrossRef]
- Haggard, B.W. Interspecies crosses in Paramecium aurelia (syngen 4 by syngen 8). J. Protozool. 1974, 21, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Lynn, D.H.; Strüder-Kypke, M. Species of Tetrahymena identical by small subunit rRNA gene sequences are discriminated by mitochondrial cytochrome c oxidase I gene sequences. J. Eukaryot. Microbiol. 2006, 53, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Poekhin, A.; Przyboś, E.; Nekrasova, I.; Yashchenko, V.; Rautian, M. Species of the Paramecium aurelia complex in Russia: New stands and overall distribution. Folia Biol. 2010, 58, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarz, S.; Sawka-Gądek, N.; Przyboś, E. Worldwide sampling reveals low genetic variability in populations of the freshwater ciliate Paramecium biaurelia (P. aurelia species complex, Ciliophora, Protozoa). Org. Divers. Evol. 2018, 18, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Aury, J.-M.; Jaillon, O.; Duret, L.; Noël, B.; Jubin, C.; Porcel, B.M.; Segurens, B.; Daubin, V.; Anthouard, V.; Aiach, N.; et al. Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 2006, 444, 171–178. [Google Scholar] [CrossRef]
- McGrath, C.L.; Gout, J.F.; Johri, P.; Doak, T.G.; Lynch, M. Differential retention and divergent resolution of duplicate genes following whole-genome duplication. Genome Res. 2014, 24, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Jori, P.; Krenek, S.; Marinov, G.K.; Doak, T.G.; Berendonk, T.U.; Lynch, M. Population genomics of Paramecium species. Mol. Biol. Evol. 2017, 34, 1194–1216. [Google Scholar] [CrossRef]
- Steck, T.; Przyboś, E.; Kusch, J.; Schmidt, H.J. Intra-species differentiation and level of inbreeding of different sibling species of the Paramecium aurelia complex. Acta Protozool. 2000, 39, 15–22. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Species | Sibling Species/Intraspecific Group | Strain Index | Waterbody | Origin | State | Coordinates | Bacterial Symbionts |
|---|---|---|---|---|---|---|---|
| The Paramecium aurelia species complex | P. primaurelia | CH | Water supply pond | Temozon, near Cenote Hubiku | Yucatán | 20°49′05″ N/88°10′25″ W | ND 1 |
| P. triaurelia | Chp3-1 | Lake | Mexico City, Chapultepec lake | Ciudad de México | 19°25′23″ N/99°11′07″ W | ND | |
| P. tetraurelia | X38 | Lake | Mexico City, Xochimilco lake | Ciudad de México | 19°16′46″ N/99°06′09″ W | Cytoplasmic, R-body-producing | |
| P. octaurelia | CA1 | Cenote | Cenote Azul, Bacalar | Quintana Roo | 18°38′51″ N/88°24′45″ W | Cytoplasmic | |
| P. quindecaurelia n. sp. | A65 | Pond | Amealco | Querétaro | 20°11′22″ N/100°08′28″ W | ND | |
| D88 | Drain | Mexico City, Los Dinamos | Ciudad de México | 19°16′02″ N/99°17′31″ W | ND | ||
| P. jenningsi | Syngen 3 | DK | Roof garden | Mexico City | Ciudad de México | 19°25′36″N/99°09′35″W | ND |
| P. caudatum | NA 2 | K1-1 | Pond | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | ND |
| K5-2 | Pond | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | ND | ||
| V-1 | Stream | Santuario Bosque de Niebla | Veracruz | 19°30′47″ N/96°56′49″ W | ND | ||
| P. multimicronucleatum | Clade I | Chp5-3 | Lake | Mexico City, Chapultepec lake | Ciudad de México | 19°25′23″ N/99°11′07″ W | Intranuclear, in macronucleus 3 |
| Chp3-4 | Lake | Mexico City, Chapultepec lake | Ciudad de México | 19°25′23″ N/99°11′07″ W | Intranuclear, in macronucleus | ||
| E59 | Lake | Endho lake | Hidalgo | 20°08′25″ N/99°21′41″ W | ND | ||
| Clade II | K4-2 | Pond | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | ND | |
| Chp10-2 | Lake | Mexico City, Chapultepec lake | Ciudad de México | 19°25′23″ N/99°11′07″ W | ND | ||
| R49 | Lake | Requena lake | Hidalgo | 19°56′31″ N/99°19′54″ W | ND | ||
| R51 | Lake | Requena lake | Hidalgo | 19°56′31″ N/99°19′54″ W | Intranuclear, in macronucleus | ||
| R53 | Lake | Requena lake | Hidalgo | 19°56′31″ N/99°19′54″ W | ND | ||
| R58 | Lake | Requena lake | Hidalgo | 19°56′31″ N/99°19′54″ W | ND | ||
| L72 | Wetlands | Lerma | Estado de México | 19°15′35″ N/99°29′29″ W | ND | ||
| SMM80-11 | Lake | San Miguel Almaya | Estado de México | 19°12′53″ N/99°26′18″ W | Cytoplasmic | ||
| SK6 | Wetlands | Laguna Chunyaxché, Sian Ka’an | Quintana Roo | 20°04′15″ N/87°34′24″ W | ND | ||
| LB2 | Lagoon | Bacalar | Quintana Roo | 18°41′46″ N/88°22′34″ W | ND | ||
| Clade III | T42 | Canal | Mexico City, Tláhuac | Ciudad de México | 19°15′59″ N/99°00′31″ W | Cytoplasmic | |
| SMM81 | Lake | San Miguel Almaya | Estado de México | 19°12′53″ N/99°26′18″ W | ND | ||
| P. putrinum | NA | K6 | Lake | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | Cytoplasmic 4 |
| K8 | Lake | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | Cytoplasmic | ||
| K11-3 | Stream | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | Cytoplasmic | ||
| P. bursaria | Syngen R3 | K11-4 | Stream | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | ND |
| K15-1 | Lake | Mexico City, Cantera Oriente | Ciudad de México | 19°19′05″ N/99°10′22″ W | ND | ||
| A66 | Lake | Amealco | Querétaro | 20°11′22″ N/100°08′28″ W | ND |
| A65-1 P. quindecaurelia (this study) | D88-8 P. quindecaurelia (this study) | 114 P. biaurelia (HF969032.1) | V1-4 P. biaurelia (PBIAU.V1-4.MITO.1.G040) | 11III1 P. biaurelia (KX008305.1) | FGC3 P. biaurelia (MG013508.1) | |
|---|---|---|---|---|---|---|
| A65-1 P. quindecaurelia (this study) | 100 | 99.3 (755/760) | 94.6 (671/709) | 95.0 (722/760) | 94.9 (721/760) | 94.8 (651/687) |
| D88-8 P. quindecaurelia (this study) | 99.3 (755/760) | 100 | 94.5 (670/709) | 94.6 (719/760) | 94.5 (718/760) | 94.6 (650/687) |
| 114 P. biaurelia (HF969032.1) | 94.6 (671/709) | 94.5 (670/709) | 100 | 99.9 (708/709) | 99.7 (707/709) | 99.9 (686/687) |
| V1-4 P. biaurelia (PBIAU.V1-4.MITO.1.G040) | 95.0 (722/760) | 94.6 (719/760) | 99.9 (708/709) | 100 | 99.9 (756/757) | 100% (687/687) |
| 11III1 P. biaurelia (KX008305.1) | 94.9 (721/760) | 94.5 (718/760) | 99.7 (707/709) | 99.9 (756/757) | 100 | 99.9 (686/687) |
| FGC3 P. biaurelia (MG013508.1) | 94.8 (651/687) | 94.6 (650/687) | 99.9 (686/687) | 100% (687/687) | 99.9 (686/687) | 100 |
| 87/5 P. pentaurelia (PPENT.87.MITO.1.G040) | NR-2/5 P. pentaurelia (This Study) | Nr1-9/5 P. pentaurelia (This Study) | AZ9-3/1 P. primaurelia (PPRIM.AZ9-3.MITO.1.G040) | OLI-prim01 P. primaurelia FJ905141.1 | IR4-2 P. primaurelia (This Study) | FT11 P. primaurelia (This Study) | CH P. primaurelia (This Study) | |
|---|---|---|---|---|---|---|---|---|
| 87/5 P. pentaurelia (PPENT.87.MITO.1.G040) | 100 | 99.9 (652/653) | 100 (767/767) | 94.3 (712/755) | 94.2 (711/755) | 94.4 (724/767) | 94.3 (723/767) | 94.1 (722/767) |
| NR-2/5 P. pentaurelia (this study) | 99.9 (652/653) | 100 | 99.9 (652/653) | 94.3 (616/653) | 94.2 (615/653) | 94.2 (615/653) | 94.2 (615/653) | 94.0 (614/653) |
| Nr1-9/5 P. pentaurelia (this study) | 100 (767/767) | 99.9 (652/653) | 100 | 94.3 (712/755) | 94.2 (711/755) | 94.4 (724/767) | 94.3 (723/767) | 94.1 (722/767) |
| AZ9-3/1 P. primaurelia (PPRIM.AZ9-3.MITO.1.G040) | 94.3 (712/755) | 94.3. (616/653) | 94.3 (712/755) | 100 | 99.9 (759/760) | 99.5 (751/755) | 99.9 (754/755) | 99.8 (753/755) |
| OLI-prim01 P. primaurelia FJ905141.1 | 94.2 (711/755) | 94.2 (615/653) | 94.2 (711/755) | 99.9 (759/760) | 100 | 99.4 (750/755) | 100 (755/755) | 99.9 (754/755) |
| IR4-2 P. primaurelia (this study) | 94.4 (724/767) | 94.2 (615/653) | 94.4 (724/767) | 99.5 (751/755) | 99.4 (750/755) | 100 | 99.4 (762/767) | 99.2 (761/767) |
| FT11 P. primaurelia (this study) | 94.3 (723/767) | 94.2 (615/653) | 94.3 (723/767) | 99.9 (754/755) | 100 (755/755) | 99.4 (762/767) | 100 | 99.9 (766/767) |
| CH P. primaurelia (this study) | 94.1 (722/767) | 94.0 (614/653) | 94.1 (722/767) | 99.8 (753/755) | 99.9 (754/755) | 99.2 (761/767) | 99.9 (766/767) | 100 |
| Species/Mexican States | CdM | Chi | EdM | Gro | Hgo | Mor | Oax | Pue | Qro | Q.R. | Sin | Ver | Yuc | CA * | SA * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. aurelia | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| P. primaurelia | X | X | X | ||||||||||||
| P. biaurelia | X | ||||||||||||||
| P. triaurelia | X | ||||||||||||||
| P. tetraurelia | X | X | X | X | |||||||||||
| P. sexaurelia | X | X | |||||||||||||
| P. octaurelia | X | X | |||||||||||||
| P. tredecaurelia | X * | X | |||||||||||||
| P. quadecaurelia | X | ||||||||||||||
| P. quindecaurelia | X | X | |||||||||||||
| P. bursaria | X | X | X | X | X | X | |||||||||
| P. calkinsi | X | X | |||||||||||||
| P. caudatum | X | X | X | X | X | X | X | X | X | X | X | ||||
| P. duboscqui | X | ||||||||||||||
| P. jenningsi | X | X | |||||||||||||
| P. multimicronucleatum | X | X | X | X | X | X | X | X | X | ||||||
| P. polycaryum | X | ||||||||||||||
| P. putrinum | X | X | X |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potekhin, A.; Mayén-Estrada, R. Paramecium Diversity and a New Member of the Paramecium aurelia Species Complex Described from Mexico. Diversity 2020, 12, 197. https://doi.org/10.3390/d12050197
Potekhin A, Mayén-Estrada R. Paramecium Diversity and a New Member of the Paramecium aurelia Species Complex Described from Mexico. Diversity. 2020; 12(5):197. https://doi.org/10.3390/d12050197
Chicago/Turabian StylePotekhin, Alexey, and Rosaura Mayén-Estrada. 2020. "Paramecium Diversity and a New Member of the Paramecium aurelia Species Complex Described from Mexico" Diversity 12, no. 5: 197. https://doi.org/10.3390/d12050197