Phylogenetic Assessment of Freshwater Mussels Castalia ambigua and C. inflata at an Ecotone in the Paraguay River Basin, Brazil Shows That Inflated and Compressed Shell Morphotypes Are the Same Species


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
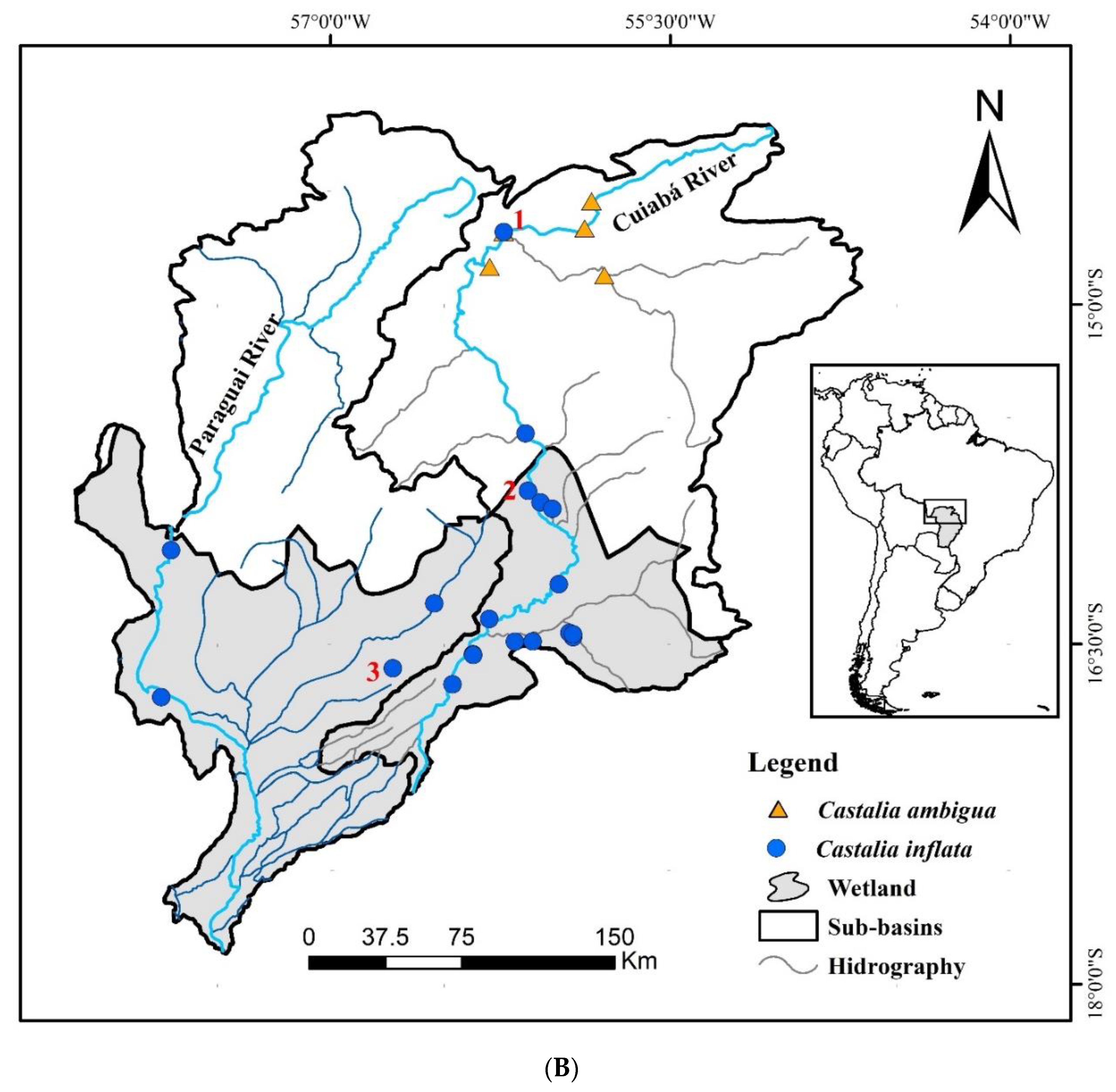
2.1. Study Area and Sample Collection
2.2. Biometrical Measurements
2.3. Molecular and Phylogenetic Analyses
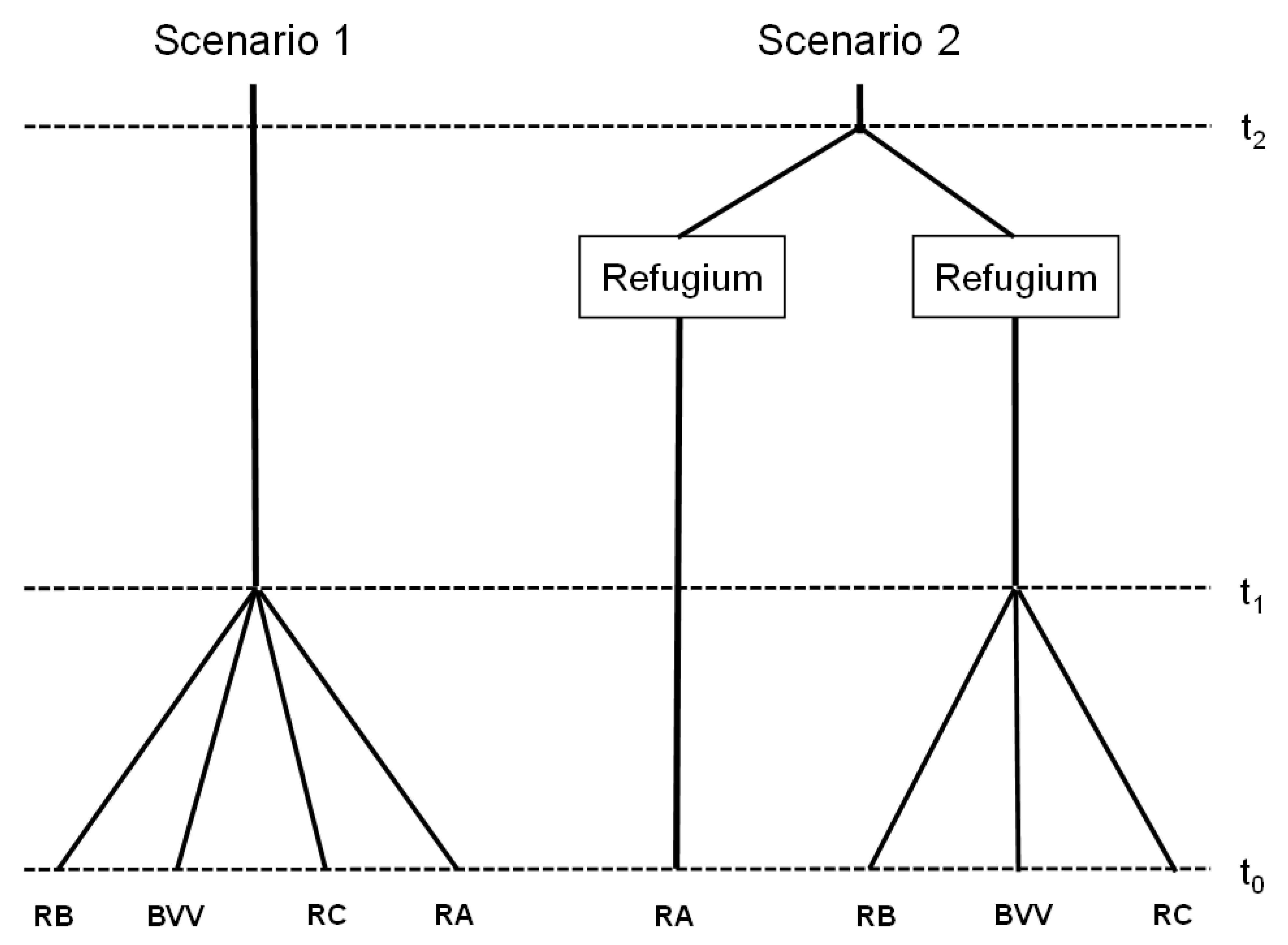
2.4. Inference of Historical Demography
2.5. Permits and Ethical Aspects
3. Results
3.1. Morphology and Distribution
3.2. Biometry
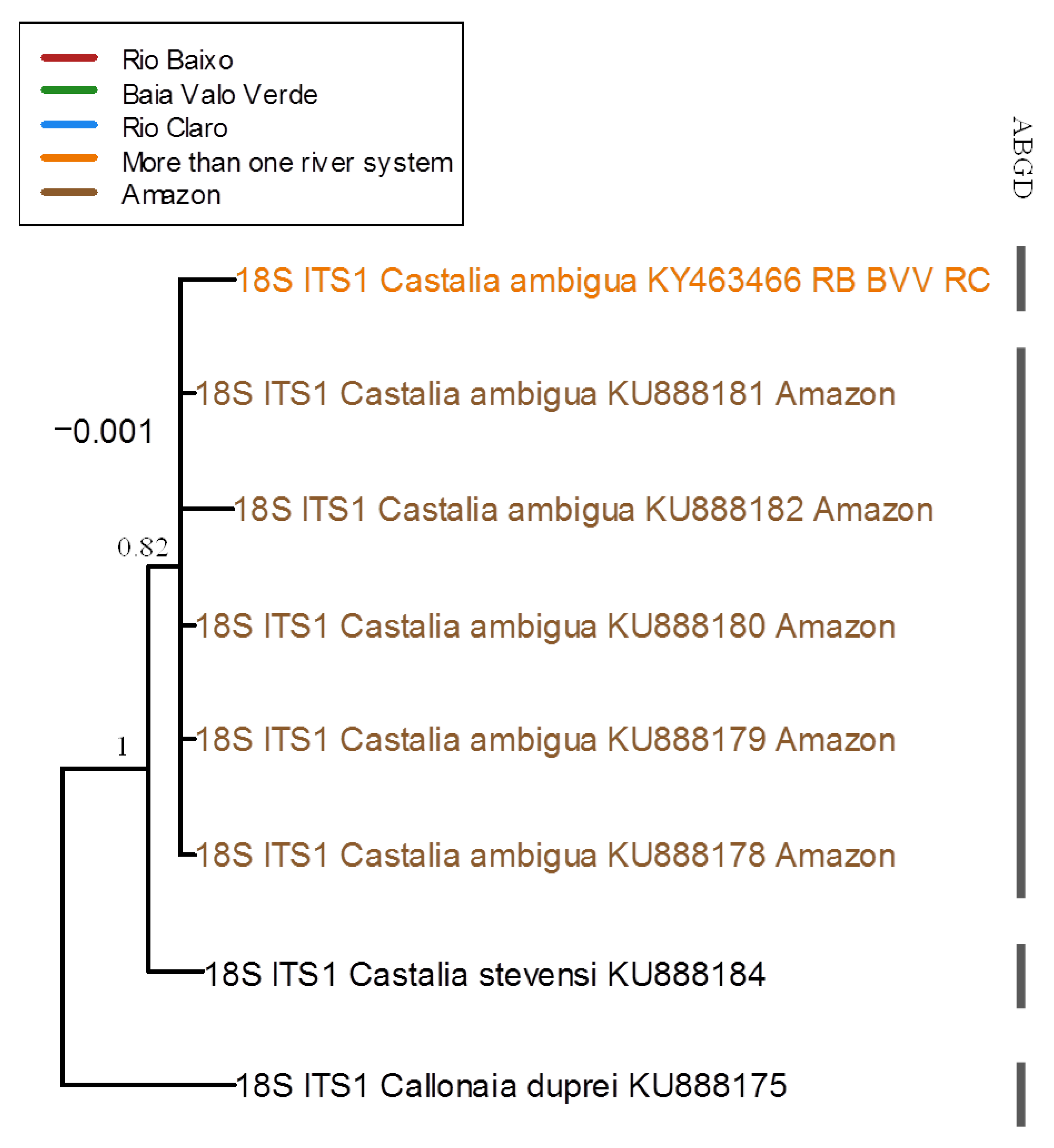
3.3. Molecular and Phylogenetic Analyses
3.4. Historical Demography
4. Discussion
4.1. Distribution, Morphology, and Taxonomy
4.2. Molecular and Phylogenetic Analyses
4.3. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mansur, M.C.D.; Pereira, D.; Bergonci, P.E.A.; Pimpão, D.M.; Barradas, J.R.S.; Sabaj, M.H. Morphological assessment of Rheodreissena (Bivalvia: Veneroida: Dreissenidae) with an updated diagnosis of the genus, descriptions of two new species, redescription of R. lopesi, and the first account of larval brooding in New World dreissenids. Proc. Acad. Nat. Sci. Phila. 2019, 166, 1–45. [Google Scholar] [CrossRef]
- Jaeckel, S.G.A. Die mollusken Sudamerikas. In Biogeography and Ecology in South America; Fittkau, E.J., Illiea, J., Klinge, H., Schwabe, G.H., Sioli, H., Eds.; Dr. W. Junk n. v. Publishers: The Hague, The Netherlands, 1969; Volume 2, pp. 794–827. [Google Scholar]
- Pereira, D.; Mansur, M.C.D.; Duarte, L.D.S.; de Oliveira, A.S.; Pimpao, D.M.; Callil, C.T.; Ituarte, C.; Parada, E.; Peredo, S.; Darrigan, G.; et al. Bivalve distribution in hydrographic regions in South America: Historical overview and conservation. Hydrobiologia 2014, 735, 15–44. [Google Scholar] [CrossRef]
- Torres, S.; Cao, L.; Gregoric, D.E.G.; de Lucía, M.; Brea, F.; Darrigran, G. Distribution of the Unionida (Bivalvia, Paleoheterodonta) from Argentina and its conservation in the southern Neotropical region. PLoS ONE 2018, 13, e0203616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonetto, A.A. Notas Sobre los Generos Castalina y Castalia en el Paraná Médio e Inferior; Direccion General de Recursos Naturales, Ministerio de Agricultura e Ganaderia: Santa Fé, Brazil, 1961. [Google Scholar]
- Vale, R.S.; Beasley, C.R.; Tagliaro, C.H.; Mansur, M.C.D. The glochidium and marsupium of Castalia ambigua ambigua Lamarck, 1819, from northern Brazil. Am. Malacol. Bull. 2005, 20, 43–48. [Google Scholar]
- Mansur, M.C.D. Lista dos Moluscos bivalves das famílias Hyriidae e Mycetopodidae para o estado do Rio Grande do Sul. Iheringia Série Zool. 1970, 39, 33–95. [Google Scholar]
- Graf, D.L.; Cummings, K.S. MUSSELp: The Mussel Project Website. 2015. Available online: http://mussel-project.uwsp.edu/ (accessed on 9 October 2018).
- D’Orbigny, A. Voyage dans L’Amerique Méridionale. Tome Cinquéme, 3a Partie: Molusques; P Bertrand: Paris, France, 1843. [Google Scholar]
- Ortmann, A.E. South American naiads: A contribution to the knowledge of the freshwater mussels of South America. Mem. Carnegie Mus. 1921, 8, 415–670. [Google Scholar]
- Bonetto, A.A. Las almejas sudamericanas de la Tribu Castaliini. Physis 1965, 25, 187–196. [Google Scholar]
- Rumi, A.; Gregoric, D.E.G.; Nunez, V.; Darrigan, G.A. Malacología Latinoamericana: Moluscos de agua dulce de Argentina. Rev. Biol. Trop. 2008, 56, 77–111. [Google Scholar] [CrossRef] [Green Version]
- Figueiras, A. La malacofauna dulceacuícola del Uruguay (Parte II). Commun. Soc. Malacol. Urug. 1965, 1, 223–270. [Google Scholar]
- Schade, F.H. Lista de los moluscos del Guaira (Villarrica—Paraguay) conocidos hasta el presente. Commun. Soc. Malacol. Urug. 1965, 1, 209–221. [Google Scholar]
- Simone, L.R.L. Land and Freshwater Molluscs of Brazil: An Illustrated Inventory of the Brazilian Malacofauna, Including Neighboring Regions of South America, Respect to the Terrestrial and Freshwater Ecosystems; FAPESP (Sao Paulo Research Foundation): Sao Paulo, Brazil, 2006. [Google Scholar]
- Graf, D.L.; Cummings, K.S. Review of the systematics and global diversity of freshwater mussel species (Bivalvia: Unionoida). J. Molluscan Stud. 2007, 73, 291–314. [Google Scholar] [CrossRef] [Green Version]
- Bogan, A.E. Mollusca-Bivalvia Checklist. Freshwater Animal Biodiversity Project. 2010. Available online: http://fada.biodiversity.be/CheckLists/Mollusca-Bivalvia.pdf (accessed on 9 October 2018).
- Haas, F. Superfamilia Unionacea. Das Tierreich 1969, 88, 1–663. [Google Scholar]
- Quintana, M.G. Catologo preliminar de la malacofauna del Paraguay. Rev. Mus. Argent. Cienc. Nat. Bernardino Rivadavia Cienc. Zool. 1982, 11, 61–158. [Google Scholar]
- Da Cruz Santos-Neto, G.; Beasley, C.R.; Schneider, H.; Pimpao, D.M.; Hoeh, W.R.; De Simone, L.R.L.; Tagliaro, C.H. Genetic relationships among freshwater mussel species from fifteen Amazonian rivers and inferences on the evolution of the Hyriidae (Mollusca: Bivalvia: Unionida). Mol. Phylogenet. Evol. 2016, 100, 148–159. [Google Scholar] [CrossRef]
- Ministerio de Agricultura y Ganaderia. Fauna Amenazada del Paraguay; Direccion de Parques Nacionales y Vida Silvestre, Fundacion Moises Bertoni: Asuncion, Paraguay, 1998. [Google Scholar]
- Colle, A.C.; Callil, C.T. Environmental influences on the composition and structure of the freshwater mussels in shallow lakes in the Cuiabá River floodplain. Braz. J. Biol. 2012, 72, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.C.L.; Callil, C.T.; Landeiro, V.L. Unraveling the effects of water–sediment conditions and spatial patterns on Unionida assemblages in seasonally connected floodplain lakes. Hydrobiologia 2020, 847, 2909–2922. [Google Scholar] [CrossRef]
- Junk, W.J.; Piedade, M.T.F.; Candotti, E. Água no Brasil: Excesso, escassez e problemas crescentes. Ciência Hoje 2014, 53, 52–53. [Google Scholar]
- Ortmann, A.E. Correlation of shape and station in fresh-water mussels (naiades). Proc. Am. Phil. Soc. 1920, 59, 268–312. [Google Scholar]
- Varis, O.; Tortajada, C.; Biswas, A.K. Management of Transboundary Rivers and Lakes; Springer: Berlin/Heidelberg, Germany, 2008; p. 271. [Google Scholar]
- Gotelli, N.; Ellison, A.M. Princípios de Estatística em Ecologia; Artmed: Porto Alegre, Brazil, 2010; 683p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 28 November 2018).
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified interface to visualize statistical result of popular R packages. R J. 2016, 8, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Naimo, T.S.; Damschen, E.D.; Rada, R.G.; Monroe, E.M. Nonlethal evaluations of the physiological health of unionid mussels: Methods for biopsy and glycogen analysis. J. N. Am. Bentholog. Soc. 1998, 17, 121–128. [Google Scholar] [CrossRef]
- Bonetto, A.A. Algunos Factores Ecologicos Vinculados a la Distribucion Geografica de las Nayades em el Rio Paraná y sus Afluentes; Dirección General de Recursos Naturales, Ministerio de Agricultura e Ganaderia: Santa Fé, Brazil, 1962. [Google Scholar]
- Serrano, M.A.; Tietbothl, R.S.; Mansur, M.C.D. Sobre a ocorrência de moluscos bivalvia no Pantanal de Mato Grosso, Brasil. Biociências 1998, 6, 131–144. [Google Scholar]
- Castillo, A.R.; Brasil, L.G.; Mansur, M.C.D.; Querol, E.; Querol, M.V.M.; Olivera, E.V. Moluscos bivalves da localidade de São Marcos, bacia do Médio rio Uruguai, Uruguaiana, Brasil. Biotemas 2007, 20, 73–79. [Google Scholar]
- Wantzen, K.M.; Callil, C.T.; Butakka, C.M.M. Benthic invertebrates of the Pantanal and its tributaries. In The Pantanal: Ecology, Biodiversity and Sustainable Management of a Large Neotropical Seasonal Wetland; Junk, W.J., Wantzen, K.M., da Cunha, C.N., da Silva, C.J., Eds.; Pensoft: Sofia, Bulgaria, 2011; pp. 127–141. [Google Scholar]
- Beasley, C.R. The impact of exploitation on freshwater mussel (Bivalvia: Hyriidae) in the Tocantins River Brazil. Stud. Neotrop. Fauna Environ. 2001, 36, 159–165. [Google Scholar] [CrossRef]
- Pimpao, D.M.; Rocha, M.S.; Fettuccia, D.C. Freshwater mussels of Catalão, confluence of Solimões and Negro rivers, state of Amazonas, Brazil. Check List 2008, 4, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Pimpao, D.M.; Mansur, M.C.D. Chave pictórica para identificação dos bivalves do Rio Aripuanã, Amazonas, Brasil (Sphaeriidae, Hyriidae e Mycetopodidae). Biota Neotrop. 2009, 9, 1–8. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijnhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Campbell, D.C.; Serb, J.M.; Buhay, J.E.; Roe, K.J.; Minton, R.L.; Lydeard, C. Phylogeny of North American amblemines (Bivalvia, Unionoida): Prodigious polyphyly proves pervasive across genera. Invert. Biol. 2005, 124, 131–164. [Google Scholar] [CrossRef]
- Walker, J.M.; Curole, J.P.; Wade, D.E.; Chapman, E.G.; Bogan, A.E.; Watters, G.T.; Hoeh, W.R. Taxonomic distribution and phylogenetic utility of gender-associated mitochondrial genomes in the Unionoida (Bivalvia). Malacologia 2006, 48, 265–282. [Google Scholar]
- Varela, E.S.; Beasley, C.R.; Schneider, H.; Sampaio, I.; Marquez-Silva, N.D.S.; Tagliaro, C.H. Molecular phylogeny of mangrove oysters (Crassostrea) from Brazil. J. Molluscan Stud. 2007, 73, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S.; Martin, A.; Romano, S.; McMillian, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Privately published document compiled by S. Palumbi; Department of Zoology, University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Pleyte, K.A.; Duncan, S.D.; Phillips, R.B. Evolutionary relationships of the salmonid fish genus Salvelinus inferred from DNA sequences of the first internal transcribed spacer (ITS 1) of ribosomal DNA. Mol. Phylogenet. Evol. 1992, 1, 223–230. [Google Scholar] [CrossRef]
- Park, J.-K.; O’Foighil, D. Sphaeriid and corbiculid clams represent separate heterodont bivalve radiations into freshwater environments. Mol. Phylogenet. Evol. 2000, 14, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Colgan, D.J.; McLaughlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löytynoja, A.; Goldman, N. webPRANK: A phylogeny-aware multiple sequence aligner with interactive alignment browser. BMC Bioinform. 2010, 11, 579. [Google Scholar] [CrossRef] [Green Version]
- Borschenius, F. FastGap 1.2. 2009. Available online: http://www.aubot.dk/FastGap_home.htm (accessed on 4 March 2020).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rozas, J.; Librado, P.; Sanchez-Delbarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP; Version 5.10.00; Universita de Barcelona: Barcelona, Spain, 2009; Available online: http://www.ub.edu/dnasp/ (accessed on 6 March 2020).
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Meth. Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. Parallel Distrib. Process. Symp. Int. 2002, 2, 0184. [Google Scholar]
- Tanabe, A.S. Kakusan4 and Aminosan: Two programs for comparing nonpartitioned, proportional and separate models for combined molecular phylogenetic analyses of multilocus sequence data. Mol. Ecol. Resour. 2011, 11, 914–921. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer 1.6. 2014. Available online: http://beast.bio.ed.ac.k/tracer (accessed on 6 July 2020).
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. MCMC Trace Analysis Tool; Version 1.6.0; Institute of Evolutionary Biology, University of Edinburgh: Edinburg, Scotland, 2009; Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 6 July 2020).
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, automatic barcode gap discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Paradis, E.; Klaus, S.; Schwartz, R. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Meth. Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Ravigne, V.; Estoup, A. Inference on population history and model checking using DNA sequence and microsatellite data with the software DIYABC (v1.0). BMC Bioinform. 2010, 11, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Santos, F.; Beaumont, M.A.; Robert, C.P.; Marin, J.M.; Baldong, D.J.; Guillemaud, T.; Estoup, A. Inferring population history with DIYABC: A user-friendly approach to approximate Bayesian computation. Bioinformatics 2008, 24, 2713–2719. [Google Scholar] [CrossRef] [Green Version]
- Bonetto, A.A. La Superfamília Unionacea en la cuenca Amazonica. In Atlas Simpósio Biota Amazônica 3: Limnologia; Lent, H., Ed.; CNPq: Rio de Janeiro, Brazil, 1967; pp. 63–82. [Google Scholar]
- Mansur, M.C.D.; Callil, C.T.; Aguiar, E.P.; Pedroso, L.M.G. Inventário dos Bivalves Límnicos do rio Cuiabá (MT, Brasil) Como Subsídio para o Reconhecimento da Integridade Ecológica; Relatório Final; Edital Fundação de Amparo a Pesquisa do Estado de Mato Grosso: Cuiaba, MT, Brazil, 2007. [Google Scholar]
- Rodriguez, F.J.L.; Oliver, J.L.; Marin, A.; Medina, J.R. The general stochastic model of nucleotide substitution. J. Theor. Biol. 1990, 142, 485–501. [Google Scholar] [CrossRef]
- Graf, D.L.; Ó Foighil, D. Molecular phylogenetic analysis of 28S rDNA supports a Gondwanan origin for Australasian Hyriidae (Mollusca: Bivalvia: Unionoida). Vie Milieu 2000, 50, 245–254. [Google Scholar]
- Callil, C.T.; Junk, W.J. Concentração e incorporação de mercúrio por moluscos bivalves Anodotites trapesialis (Lamarck, 1819) e Castalia ambigua (Lamarck, 1819) do Pantanal de Poconé—MT, Brasil. Biociências 1999, 72, 3–28. [Google Scholar]
- Mansur, M.C.D.; Valer, R.M. Moluscos bivalves do Rio Uraricoera e Rio Branco, Roraima, Brasil. Amazoniana 1992, 12, 85–100. [Google Scholar]
- Da Mata, L.S.; Tagliaro, C.H.; Simeone, D.; Robert, C. Shell shape variation in Amazonian freshwater mussels (Unionida: Hyriidae: Hyriini) Beasley. J. Molluscan Stud. 2019, 85, 212–223. [Google Scholar] [CrossRef]
- Haag, W.R. North American Freshwater Mussels: Natural History, Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Whelan, N.V.; Geneva, A.J.; Graf, D.L. Molecular phylogenetic analysis of tropical freshwater mussels (Mollusca: Bivalvia: Unionoida) resolves the position of Coelatura and supports a monophyletic Unionidae. Mol. Phylogenet. Evol. 2011, 61, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Zieritz, A.; Hoffman, J.I.; Amos, W.; Aldridge, D.C. Phenotypic plasticity and genetic isolation-by-distance in the freshwater mussel Unio pictorum. Evol. Ecol. 2010, 24, 923–938. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Makhrov, A.A.; Gofarov, M.Y.; Aksenova, O.V.; Aspholm, P.E.; Bespalaya, Y.V.; Kabakov, M.B.; Kolosova, Y.S.; Kondakov, A.V.; Ofenböck, T.; et al. Climate warming as a possible trigger of keystone mussel population decline in oligotrophic rivers at the continental scale. Sci. Rep. 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Hayes, D.M.; Harris, J.L.; Christian, A.D. Phylogenetic and morphometric analyses reveal ecophenotypic plasticity in freshwater mussels Obovaria jacksoniana and Villosa arkansasensis (Bivalvia: Unionidae). Ecol. Evol. 2013, 3, 2670–2683. [Google Scholar] [CrossRef] [PubMed]
- Prie, V.; Puillandre, N. Molecular phylogeny, taxonomy, and distribution of French Unio species. Hydrobiologia 2014, 735, 95–110. [Google Scholar] [CrossRef]
- Lundberg, J.G.; Marshall, L.G.; Guerrero, B.H.; Malabarba, M.C.S.L.; Wesselingh, F. The stage for Neotropical fish diversification: A history of tropical South American rivers. In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M., Lucena, C.A.S., Eds.; Edipucrs: Porto Alegre, Brazil, 1998; pp. 13–48. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake or River | Latitude | Longitude | Castalia inflata d’Orbigny, 1835 | Castalia ambigua Lamarck, 1819 | Occurrence | |
|---|---|---|---|---|---|---|
| 1 | Cuiabazinho River | 14°32′35.48″ S | 55°50′53.49″ W | - | x | Highlands |
| 2 | Cuiabazinho River | 14°39′46.63″ S | 55°52′45.30″ W | - | x | Highlands |
| 3 | APM Manso | 14°52′16.89″ S | 55°47′31.27″ W | - | x | Highlands |
| 4 | Baixo Stream | 14°40′46.39″ S | 56°14′9.72″ W | x | x | Highlands |
| 5 | Cuiabá River | 14°49′59.78″ S | 56°17′53.34″ W | - | x | Highlands |
| 6 | Cuiabá River | 15°34′08.25″ S | 56°08′23.02″ W | x | - | Lowlands |
| 7 | Valo Verde oxbow lake | 15°49′20.36″ S | 56° 7′43.59″ W | x | - | Lowlands |
| 8 | Cuiabá River | 15°52′26.66″ S | 56° 4′25.09″ W | x | Lowlands | |
| 9 | Poço oxbow lake | 15°54′06.73″ S | 56°01′17.20″ W | x | - | Lowlands |
| 10 | Paraguay River | 16°05′08.60″ S | 57°42′18.65″ W | x | - | Lowlands |
| 11 | Corixo das Conchas oxbow lake | 16°28′6.075″ S | 55°55′49.837″ W | x | - | Lowlands |
| 12 | Coqueiro oxbow lake | 16°23′19.11″ S | 56°18′00.57″ W | x | - | Lowlands |
| 13 | Jacaré (Ninhal) oxbow lake | 16°14′08.89″ S | 55°59′37.10″ W | x | - | Lowlands |
| 14 | Bento Gomes River | 16°19′13.49″ S | 56°32′36.21″ W | x | - | Lowlands |
| 15 | Pombas oxbow lake | 16°26′54.07″ S | 55°56′45.30″ W | x | - | Lowlands |
| 16 | Conchas oxbow lake | 16°27′18.80″ S | 55°55′46.40″ W | x | - | Lowlands |
| 17 | Serragem oxbow lake | 16°29′14.80″ S | 56°06′31.30″ W | x | - | Lowlands |
| 18 | Barco Velho oxbow lake | 16°29′18.00″ S | 56°11′13.15″ W | x | - | Lowlands |
| 19 | Cuiabá River | 16°32′36.45″ S | 56°22′20.90″ W | x | - | Lowlands |
| 20 | Oxbow lakes | 16°32′49.20″ S | 56°18′12.00″ W | x | - | Lowlands |
| 21 | Claro Stream | 16°36′21.84″ S | 56°43′31.67″ W | x | - | Lowlands |
| 22 | Santa Rosa oxbow lake | 16°40′33.20″ S | 56°27′45.30″ W | x | - | Lowlands |
| 23 | Descalvado oxbow lake | 16°44′00.29″ S | 57°44′55.67″ W | x | - | Lowlands |
| Gene, Primer | Primer Name | Primer Sequence (5′ to 3′) | Reference |
|---|---|---|---|
| COI F | LCO1490 | GGT CAA CAA ATC ATA AAG ATA TTG G | [38] |
| COI F | COIF | GTT CCA CAA ATC ATA AGG ATA TTG G | [39] |
| COI R | HCO700dy2 | TCA GGG TGA CCA AAA AAY CA | [40] |
| 16S F | 16SL1987 | GCC TCG CCT GTT TAC CAA AAA C | [41] |
| 16S R | 16Sbr-H | CCG GTC TGA ACT CAG ATC ACG | [42] |
| 18S ITS F | ITS1A | AAA AAG CTT TTG TAC ACA CCG CCC GTC | [43] |
| 18S ITS R | ITS1B | AGC TTG CTG CGT TCT TCA TCG A | [43] |
| 28S F | D23F | GAG AGT TCA AGA GTA CGT G | [44] |
| 28S R | D4RB | TGT TAG ACT CCT TGG TCC GTG T | [44] |
| H3 F | H3F | ATG GCT CGT ACC AAG CAG ACV GC | [45] |
| H3 R | H3R | ATA TCC TTR GGC ATR ATR GTG AC | [45] |
| River Population | Parameter | Prior Distributions | Posterior Distributions | ||||
|---|---|---|---|---|---|---|---|
| Min. | Max. | Distribution | Median | 95% Credible Interval | |||
| Baixo Stream | Ne | 10 | 50,000 | Uniform | 7660 | 1880 | 31,200 |
| Valo Verde Lake | Ne | 10 | 50,000 | Uniform | 4020 | 1130 | 15,900 |
| Claro Stream | Ne | 10 | 50,000 | Uniform | 28,800 | 10,400 | 47,100 |
| Amazon River | Ne | 10 | 50,000 | Uniform | 32,600 | 13,500 | 48,100 |
| Ancestral | Ne | 10 | 100,000 | Uniform | 77,800 | 37,700 | 98,000 |
| T1 (generations) | 10 | 20,000 | Uniform | 1290 | 366 | 6080 | |
| T2 (generations) | 1000 | 100,000 | Uniform | 7240 | 2830 | 9750 | |
| Mutation rate (µ) | 1 × 10−8 | 1 × 10−6 | Uniform | 6.64 × 10−7 | 3.64 × 10−7 | 9.5 × 10−7 | |
| 25 | 28 | 31 | 34 | 40 | 47 | 49 | 51 | 52 | 55 | 61 | 64 | 67 | 70 | 73 | 85 | 118 | 139 | 148 | 154 | 163 | 169 | 172 | |
| Cast COI 01 | G | G | A | A | G | C | T | G | G | A | C | A | A | G | G | A | G | G | G | T | G | A | G |
| Cast COI 02 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 03 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 04 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 05 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 06 | . | . | . | . | . | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 07 | . | . | . | . | . | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 08 | . | . | . | . | . | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 09 | . | . | . | . | . | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 10 | . | . | . | . | . | . | . | A | . | . | . | . | G | A | . | . | . | . | . | A | . | . | . |
| Cast COI 11 | . | . | . | . | A | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 12 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 13 | . | . | . | . | . | . | . | . | . | . | . | . | G | A | . | . | . | . | . | . | . | . | . |
| Cast COI 14 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | A |
| Cast COI 15 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 16 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Cast COI 17 | . | . | . | . | . | . | . | . | A | . | . | . | G | . | . | . | . | . | . | . | . | . | . |
| Castalia ambigua 1 | A | . | G | G | . | . | . | . | . | G | G | . | G | . | . | . | . | . | . | . | A | G | . |
| Castalia ambigua 2 | A | . | G | G | . | . | . | . | . | G | G | . | G | . | . | . | . | . | . | . | A | G | . |
| Castalia ambigua 3 | A | . | G | G | . | . | . | . | . | G | G | . | G | . | . | . | . | . | . | . | A | G | . |
| Castalia ambigua 4 | A | . | G | G | . | . | . | . | . | G | G | . | G | . | . | . | . | . | . | . | A | G | . |
| Castalia ambigua 5 | . | . | G | . | . | . | . | . | . | G | G | G | G | . | . | G | . | A | . | . | A | G | . |
| Castalia ambigua 6 | . | . | G | . | . | . | . | . | . | G | G | G | G | . | . | G | . | A | . | . | A | G | . |
| Castalia ambigua 7 | . | T | . | G | . | T | A | . | . | G | G | G | . | A | A | . | A | A | A | . | . | G | . |
| Castalia ambigua 8 | . | T | . | G | . | T | A | . | . | G | G | G | . | A | A | . | A | A | A | . | . | G | . |
| 181 | 187 | 192 | 193 | 199 | 211 | 229 | 253 | 259 | 260 | 262 | 265 | 268 | 278 | 289 | 295 | 310 | 316 | 320 | 328 | 335 | 337 | 361 | |
| Cast COI 01 | A | G | C | T | T | T | G | G | A | T | A | A | G | G | A | G | G | T | T | T | G | T | G |
| Cast COI 02 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 03 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 04 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 05 | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . |
| Cast COI 06 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 07 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 08 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 09 | G | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 10 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 11 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 12 | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 13 | . | A | . | . | . | . | . | . | . | . | . | G | . | A | . | . | . | . | . | . | . | . | . |
| Cast COI 14 | . | A | . | . | . | . | . | . | . | C | . | G | . | . | . | . | . | . | . | C | . | . | . |
| Cast COI 15 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 16 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast COI 17 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Castalia ambigua 1 | . | . | . | . | . | . | A | . | . | . | G | . | A | . | T | A | . | C | . | . | T | . | A |
| Castalia ambigua 2 | . | . | . | . | . | . | A | . | . | . | G | . | A | . | T | A | . | C | . | . | T | . | A |
| Castalia ambigua 3 | . | . | . | . | . | . | A | . | . | . | G | . | A | . | T | A | . | C | . | . | T | C | A |
| Castalia ambigua 4 | . | . | . | . | . | . | A | . | . | . | G | . | A | . | T | A | . | C | . | . | T | C | A |
| Castalia ambigua 5 | . | . | . | . | C | . | . | . | . | . | . | . | A | . | T | A | . | A | C | . | . | C | A |
| Castalia ambigua 6 | . | . | . | . | C | . | . | . | . | . | . | . | A | . | T | A | . | A | C | . | . | C | A |
| Castalia ambigua 7 | . | . | . | C | . | A | . | A | G | . | G | . | . | . | T | . | A | . | . | . | . | . | . |
| Castalia ambigua 8 | . | . | . | C | . | A | . | A | G | . | G | . | . | . | T | . | A | . | . | . | . | . | . |
| 376 | 379 | 388 | 397 | 406 | 424 | 430 | 442 | 451 | 457 | 463 | 467 | 491 | 535 | C. ambigua | C. inflata | C. ambigua-like | Rio Baixo | Baia Valo Verde | Rio Claro | Rio Amazonas | Accession Number | ||
| Cast COI 01 | G | T | A | A | G | C | T | T | G | G | A | A | G | T | 1 | 1 | KY474356 | ||||||
| Cast COI 02 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 23 | 5 | 18 | KY474357 | |||||
| Cast COI 03 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 8 | 1 | 3 | 5 | KY474358 | |||
| Cast COI 04 | . | . | . | . | . | . | . | C | . | . | . | . | A | . | 1 | 1 | KY474359 | ||||||
| Cast COI 05 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 6 | 6 | 4 | 16 | KY474360 | ||||
| Cast COI 06 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 4 | 1 | 5 | KY474361 | |||||
| Cast COI 07 | . | . | . | . | . | . | . | C | . | . | . | . | . | G | 2 | 1 | 3 | KY474362 | |||||
| Cast COI 08 | . | . | . | . | . | . | . | C | . | A | . | . | . | . | 1 | 1 | KY474363 | ||||||
| Cast COI 09 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474364 | ||||||
| Cast COI 10 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474365 | ||||||
| Cast COI 11 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 2 | 2 | 4 | KY474366 | |||||
| Cast COI 12 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474367 | ||||||
| Cast COI 13 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474368 | ||||||
| Cast COI 14 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | KY474369 | ||||||
| Cast COI 15 | A | . | . | G | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474370 | ||||||
| Cast COI 16 | A | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474371 | ||||||
| Cast COI 17 | . | . | . | . | . | . | . | C | . | . | . | . | . | . | 1 | 1 | KY474372 | ||||||
| Castalia ambigua 1 | A | C | . | G | . | T | C | . | . | . | . | . | - | - | 1 | 1 | KU888236 | ||||||
| Castalia ambigua 2 | A | C | . | G | . | T | C | . | . | . | . | . | - | - | 1 | 1 | KU888237 | ||||||
| Castalia ambigua 3 | A | C | . | . | . | T | C | . | . | . | . | . | - | - | 1 | 1 | KU888238 | ||||||
| Castalia ambigua 4 | A | C | . | . | . | T | C | . | . | . | . | . | - | - | 1 | 1 | KU888239 | ||||||
| Castalia ambigua 5 | A | C | G | . | . | T | G | . | . | T | . | G | - | - | 1 | 1 | KU888240 | ||||||
| Castalia ambigua 6 | A | C | G | . | . | T | G | . | . | T | . | G | - | - | 1 | 1 | KU888241 | ||||||
| Castalia ambigua 7 | A | . | . | . | A | T | . | . | A | T | G | . | - | - | 1 | 1 | KU888242 | ||||||
| Castalia ambigua 8 | A | . | . | . | A | T | . | . | A | T | G | . | - | - | 1 | 1 | KU888243 | ||||||
| Species | Collection Area | n | Number of Haplotypes | Average Number of Nucleotide Differences (k) | Haplotype Diversity (h) | Nucleotide Diversity (π) |
|---|---|---|---|---|---|---|
| C. ambigua | Baixo Stream | 19 | 9 | 3.81 | 0.865 | 0.007 |
| C. inflata | Baixo Stream | 7 | 2 | 1.71 | 0.286 | 0.003 |
| C. ambigua-like | Baixo Stream | 8 | 4 | 4.18 | 0.750 | 0.008 |
| C. inflata | Valo Verde Lake | 10 | 4 | 0.93 | 0.711 | 0.002 |
| C. inflata | Claro Stream | 27 | 6 | 1.09 | 0.536 | 0.002 |
| C. ambigua | Amazon River | 8 | 4 | 18.29 | 0.857 | 0.040 |
| 35 | 36 | 83 | 87 | 92 | 98 | 111 | 119 | 124 | 125 | 138 | 149 | 176 | 184 | 186 | 191 | 197 | 200 | 206 | 211 | 214 | 217 | 246 | 258 | 262 | 263 | |
| Cast 16S 1 | A | A | C | T | C | A | C | C | C | C | G | C | G | T | A | T | T | T | A | C | T | A | T | T | C | C |
| Cast 16S 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 3 | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 4 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 5 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 6 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | T | . |
| Cast 16S 7 | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 8 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cast 16S 9 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | . | . |
| C. ambigua 1 | C | T | . | . | . | . | . | T | . | . | . | T | . | C | . | . | C | . | C | . | C | . | . | . | T | . |
| C. ambigua 2 | T | . | . | . | . | . | . | T | . | . | . | T | . | C | . | . | C | . | C | . | C | . | . | . | T | . |
| C. ambigua 3 | . | . | . | . | . | . | . | T | . | . | . | T | . | C | . | . | C | C | C | . | C | . | . | . | T | . |
| C. ambigua 4 | . | . | . | . | . | . | . | T | . | T | . | T | . | C | . | . | C | . | C | . | C | . | . | . | T | . |
| C. ambigua 5 | T | . | . | . | . | . | . | T | . | . | . | T | . | . | . | . | C | . | . | T | . | . | . | . | T | . |
| C. ambigua 6 | C | . | . | . | . | . | . | T | . | . | . | T | . | . | . | . | C | . | . | T | . | . | . | . | T | . |
| C. ambigua 7 | T | T | . | C | A | G | T | . | T | . | A | T | . | C | . | . | C | . | . | . | C | G | C | C | A | T |
| 265 | 269 | 273 | 276 | 280 | 291 | 292 | 336 | 390 | 400 | 413 | 428 | C. ambigua | C. inflata | C. ambigua-like | Rio Baixo | Baia Valo Verde | Rio Claro | Rio Amazonas | Accession number | |||||||
| Cast 16S 1 | A | G | T | C | T | T | G | C | C | T | T | G | 7 | 6 | 1 | KY463457 | ||||||||||
| Cast 16S 2 | . | . | . | . | . | . | . | . | . | C | . | . | 18 | 30 | 7 | 32 | 3 | 20 | KY463458 | |||||||
| Cast 16S 3 | . | . | . | . | . | . | . | T | T | C | . | . | 1 | 1 | KY463459 | |||||||||||
| Cast 16S 4 | . | . | . | . | . | . | . | . | . | C | . | T | 1 | 1 | KY463460 | |||||||||||
| Cast 16S 5 | . | . | . | . | . | . | . | . | . | C | . | . | 1 | 1 | KY463461 | |||||||||||
| Cast 16S 6 | . | . | . | . | . | . | . | . | . | C | C | . | 2 | 2 | KY463462 | |||||||||||
| Cast 16S 7 | . | . | . | . | . | . | . | T | . | C | . | . | 1 | 1 | KY463463 | |||||||||||
| Cast 16S 8 | . | . | . | . | . | . | A | . | . | C | . | . | 1 | 1 | KY463464 | |||||||||||
| Cast 16S 9 | . | . | . | . | . | . | . | . | . | C | . | . | 1 | 1 | KY463465 | |||||||||||
| C. ambigua 1 | C | . | . | A | C | C | . | A | - | - | - | - | 1 | 1 | KU888207 | |||||||||||
| C. ambigua 2 | C | . | . | A | C | C | . | A | - | - | - | - | 1 | 1 | KU888208 | |||||||||||
| C. ambigua 3 | C | . | . | A | C | C | . | T | - | - | - | - | 1 | 1 | KU888209 | |||||||||||
| C. ambigua 4 | C | . | . | A | C | C | . | T | - | - | - | - | 1 | 1 | KU888210 | |||||||||||
| C. ambigua 5 | C | . | C | A | C | . | . | A | - | - | - | - | 1 | 1 | KU888211 | |||||||||||
| C. ambigua 6 | C | . | C | A | C | . | . | A | - | - | - | - | 1 | 1 | KU888212 | |||||||||||
| C. ambigua 7 | C | A | C | T | C | C | . | A | - | - | - | - | 1 | 1 | KU888213 | |||||||||||
| Species | Collection Area | n | Number of Haplotypes | Average Number of Nucleotide Differences (k) | Haplotype Diversity (h) | Nucleotide Diversity (π) |
|---|---|---|---|---|---|---|
| C. ambigua | Baixo Stream | 19 | 2 | 0.105 | 0.105 | 0.0002 |
| C. inflata | Baixo Stream | 7 | 1 | 0.000 | 0.000 | 0.0000 |
| C. ambigua-like | Baixo Stream | 8 | 2 | 0.250 | 0.250 | 0.0006 |
| C. inflata | Valo Verde Lake | 10 | 3 | 1.133 | 0.600 | 0.0026 |
| C. inflata | Claro Stream | 26 | 6 | 0.680 | 0.412 | 0.0015 |
| C. ambigua | Amazon River | 7 | 7 | 9.333 | 1.000 | 0.0308 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivera-Hyde, M.; Hallerman, E.; Santos, R.; Jones, J.; Varnerin, B.; da Cruz Santos Neto, G.; Mansur, M.C.; Moraleco, P.; Callil, C. Phylogenetic Assessment of Freshwater Mussels Castalia ambigua and C. inflata at an Ecotone in the Paraguay River Basin, Brazil Shows That Inflated and Compressed Shell Morphotypes Are the Same Species. Diversity 2020, 12, 481. https://doi.org/10.3390/d12120481
Olivera-Hyde M, Hallerman E, Santos R, Jones J, Varnerin B, da Cruz Santos Neto G, Mansur MC, Moraleco P, Callil C. Phylogenetic Assessment of Freshwater Mussels Castalia ambigua and C. inflata at an Ecotone in the Paraguay River Basin, Brazil Shows That Inflated and Compressed Shell Morphotypes Are the Same Species. Diversity. 2020; 12(12):481. https://doi.org/10.3390/d12120481
Chicago/Turabian StyleOlivera-Hyde, Miluska, Eric Hallerman, Rogério Santos, Jess Jones, Brianne Varnerin, Guilherme da Cruz Santos Neto, Maria Cristina Mansur, Priscilla Moraleco, and Claudia Callil. 2020. "Phylogenetic Assessment of Freshwater Mussels Castalia ambigua and C. inflata at an Ecotone in the Paraguay River Basin, Brazil Shows That Inflated and Compressed Shell Morphotypes Are the Same Species" Diversity 12, no. 12: 481. https://doi.org/10.3390/d12120481