Diversity and Distribution of the Dominant Ant Genus Anonychomyrma (Hymenoptera: Formicidae) in the Australian Wet Tropics


Abstract
:1. Introduction
- How many species of Anonychomyrma can be recognised in the AWT?
- How are the species distributed among subregions and along elevational and vertical gradients?
- What are the patterns of species richness and endemism?
- To what extent do species distributions correlate with contemporary climatic variables and reflect historical patterns of rainforest refugia?
2. Materials and Methods
2.1. Study Region
2.2. Ant Sampling
2.3. Gene Barcoding
2.4. Species Delimitation
2.5. Patterns of Abundance, Diversity, and Endemism
2.6. Species Distribution Modelling
3. Results
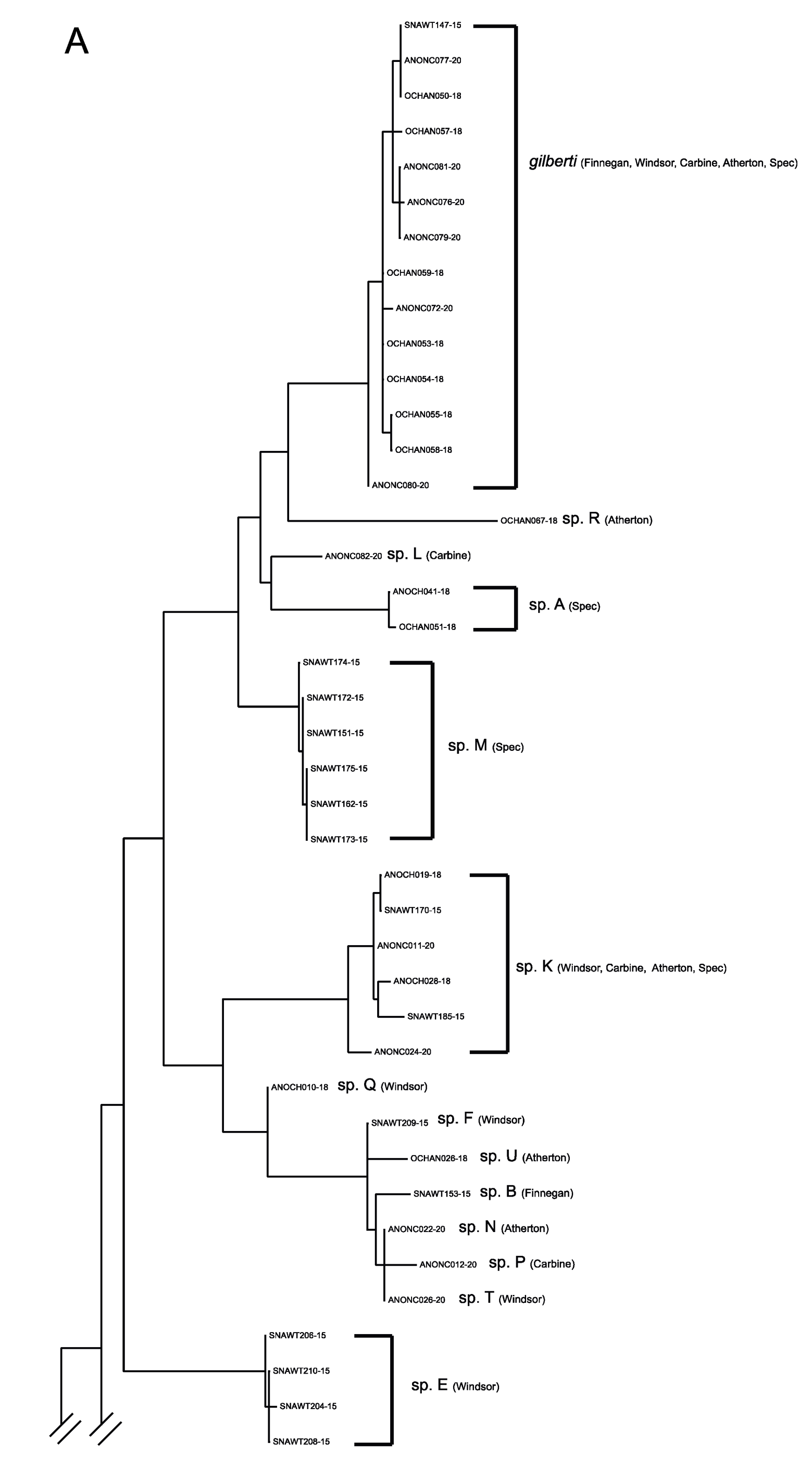
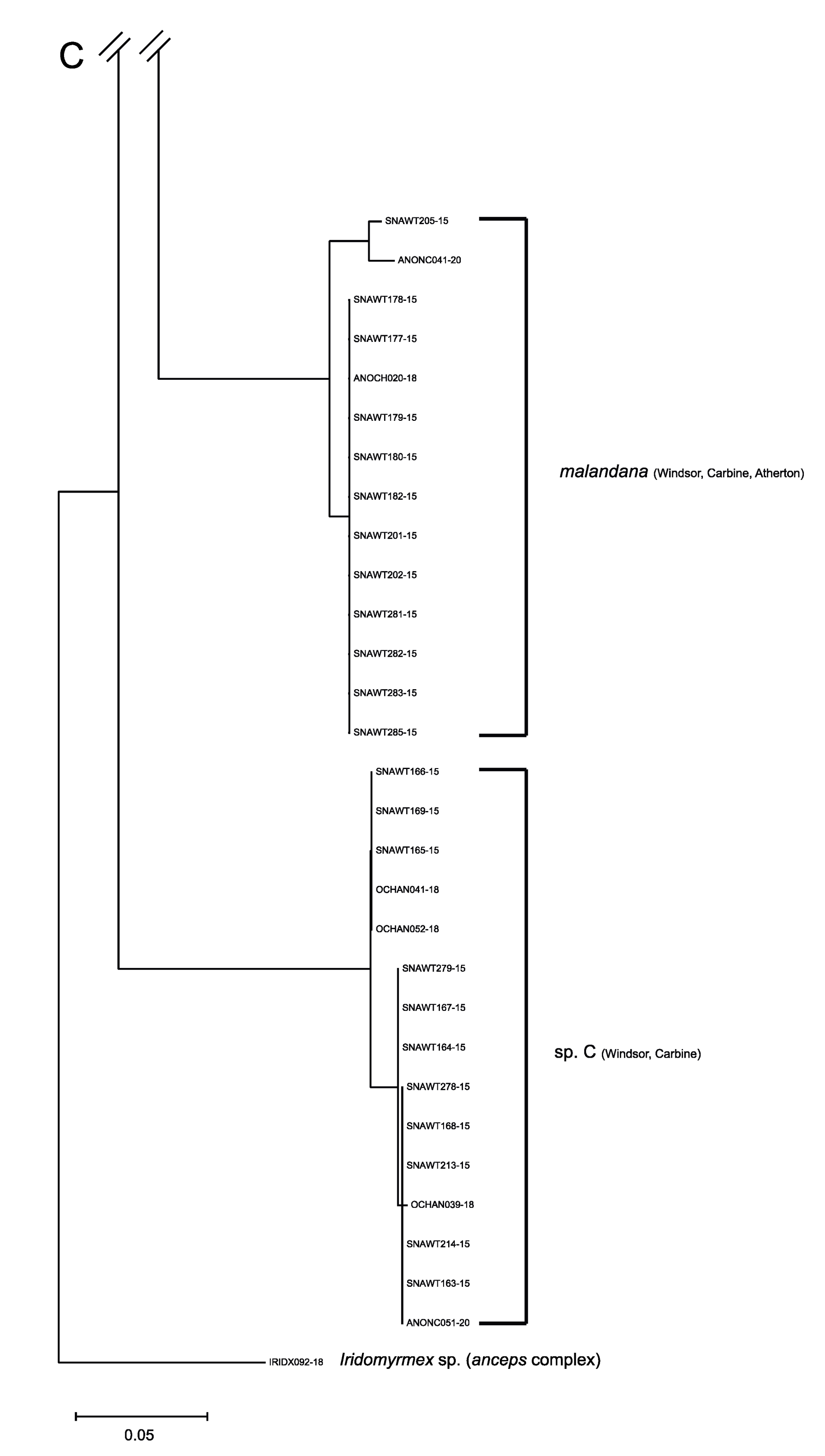
3.1. The Anonychomyrma Fauna
3.2. Patterns of Abundance, Diversity, and Endemism
3.3. Species Distribution Modelling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Dunn, R.R.; Agosti, D.; Andersen, A.N.; Arnan, X.; Bruhl, C.A.; Cerdá, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Shattuck, S.O. Review of the dolichoderine ant genus Iridomyrmex Mayr with descriptions of three new genera (Hymenoptera: Formicidae). Aust. J. Entomol. 1992, 31, 13–18. [Google Scholar] [CrossRef]
- AntWeb. Available online: https://www.antweb.org (accessed on 20 September 2020).
- Plowman, N.S.; Mottl, O.; Novotny, V.; Idigel, C.; Philip, F.J.; Rimandai, M.; Klimes, P. Nest microhabitats and tree size mediate shifts in ant community structure across elevation in tropical rainforest canopies. Ecography 2020, 43, 431–442. [Google Scholar] [CrossRef]
- Ward, P.S.; Brady, S.G.; Fisher, B.L.; Schultz, T.R. Phylogeny and biogeography of dolichoderine ants: Effects of data partitioning and relict taxa on historical inference. Syst. Biol. 2010, 59, 342–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, M.D.; Fox, B.J. Evidence for interspecific competition influencing ant species diversity in a regenerating heathland. In Ant-Plant Interactions in Australia; Buckley, R., Ed.; Springer: Dordrecht, The Netherlands, 1982; pp. 99–110. [Google Scholar]
- Andersen, A.N. Patterns of ant community organization in mesic southeastern Australia. Aust. J. Ecol. 1986, 11, 87–97. [Google Scholar] [CrossRef]
- Nowrouzi, S.; Andersen, A.N.; Macfadyen, S.; Staunton, K.M.; VanDerWal, J.; Robson, S.K.A. Ant diversity and distribution along elevation gradients in the Australian Wet Tropics: The importance of seasonal moisture stability. PLoS ONE 2016, 11, e0153420. [Google Scholar] [CrossRef]
- Blüthgen, N.; Stork, N.E. Ant mosaics in a tropical rainforest in Australia and elsewhere: A critical review. Austral Ecol. 2007, 32, 93–104. [Google Scholar] [CrossRef]
- Schneider, C.; Moritz, C. Rainforest refugia and Australia’s Wet Tropics. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Blüthgen, N.; Stork, N.; Fiedler, K. Bottom-up control and co-occurrence in complex communities: Honeydew and nectar determine a rainforest ant mosaic. Oikos 2004, 106, 344–358. [Google Scholar] [CrossRef]
- Williams, S.E.; Pearson, R.G.; Walsh, P.J. Distributions and biodiversity of the terrestrial vertebrates of Australia’s Wet Tropics: A review of current knowledge. Pac. Conserv. Biol. 1995, 2, 327–362. [Google Scholar] [CrossRef]
- Carnaval, A.C.; Moritz, C. Historical Climate Modelling Predicts Patterns of Current Biodiversity in the Brazilian Atlantic Forest. J. Biogeogr. 2008, 35, 1187–1201. [Google Scholar] [CrossRef]
- Davies, T.J.; Buckley, L.B.; Grenyer, R.; Gittleman, J.L. The influence of past and present climate on the biogeography of modern mammal diversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2526–2535. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, A. Pleistocene vegetation of the humid tropics of northeastern Queensland, Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 109, 399–412. [Google Scholar] [CrossRef]
- Hopkins, M.S.; Ash, J.; Graham, A.W.; Head, J.; Hewett, R. Charcoal evidence of the spatial extent of the Eucalyptus woodland expansions and rainforest contractions in North Queensland during the late Pleistocene. J. Biogeogr. 1993, 20, 357–372. [Google Scholar] [CrossRef]
- Williams, S.E.; Pearson, R.G. Historical rainforest contractions, localized extinctions and patterns of vertebrate endemism in the rainforests of Australia’s wet tropics. Proc. R. Soc. Lond. B Biol. Sci. 1997, 264, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Hugall, A.; Moritz, C.; Moussalli, A.; Stanisic, J. Reconciling paleodistribution models and comparative phylogeography in the Wet Tropics rainforest land snail Gnarosophia bellendenkerensis (Brazier 1875). Proc. Natl. Acad. Sci. USA 2002, 99, 6112–6117. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Graham, C.H.; Moritz, C.; Williams, S.E. Habitat history improves prediction of biodiversity in rainforest fauna. Proc. Natl. Acad. Sci. USA 2006, 103, 632–636. [Google Scholar] [CrossRef] [Green Version]
- Ponniah, M.; Hughes, J.M. The evolution of Queensland spiny mountain crayfish of the genus Euastacus. I. Testing vicariance and dispersal with interspecific mitochondrial DNA. Evolution 2004, 58, 1073–1085. [Google Scholar] [CrossRef]
- Moritz, C.; Hoskin, C.; MacKenzie, J.; Phillips, B.; Tonione, M.; Silva, N.; VanDerWal, J.; Williams, S.; Graham, C. Identification and dynamics of a cryptic suture zone in tropical rainforest. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 1235–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, C.S.; Hugall, A.F.; McDonald, K.R.; Jamieson, B.G.; Moritz, C. An ancient divide in a contiguous rainforest: Endemic earthworms in the Australian Wet Tropics. PLoS ONE 2015, 10, e0136943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussalli, A.; Moritz, C.; Williams, S.E.; Carnaval, A.C. Variable responses of skinks to a common history of rainforest fluctuation: Concordance between phylogeography and paleo-distribution models. Mol. Ecol. 2009, 18, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.E.; Middleton, J. Climatic seasonality, resource bottlenecks, and abundance of rainforest birds: Implications for global climate change. Divers. Distrib. 2008, 14, 69–77. [Google Scholar] [CrossRef]
- Frith, C.; Frith, D. Seasonality of insect abundance in an Australian upland rainforest. Aust. J. Ecol. 1985, 10, 237–248. [Google Scholar] [CrossRef]
- Blüthgen, N.; Fiedler, K. Competition for composition: Lessons from nectar-feeding ant communities. Ecology 2004, 85, 1479–1485. [Google Scholar] [CrossRef] [Green Version]
- Shoo, L.P.; Williams, S.E.; Hero, J.-M. Climate warming and the rainforest birds of the Australian Wet Tropics: Using abundance data as a sensitive predictor of change in total population size. Biol. Conserv. 2005, 125, 335–343. [Google Scholar] [CrossRef]
- McJannet, D.; Wallace, J.; Reddell, P. Precipitation interception in Australian tropical rainforests: II. Altitudinal gradients of cloud interception, stemflow, throughfall and interception. Hydrol. Process. 2007, 21, 1703–1718. [Google Scholar] [CrossRef]
- De Frenne, P.; Zellweger, F.; Rodriguez-Sanchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef]
- Leahy, L.; Scheffers, B.R.; Andersen, A.N.; Hirsch, B.T.; Williams, S.E. Vertical niche and elevation range size in tropical ants: Implications for climate resilience. Divers. Distrib. In press.
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stauffer, C.; Christian, E. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef]
- Williams, S.E.; Falconi, L.; Moritz, C.; Fenker Antunes, J. State of Wet Tropics Report 2015-2016. Ancient, Endemic, Rare and Threatened Vertebrates of the Wet Tropics; Wet Tropics Management Authority: Cairns, Australia, 2016. [Google Scholar]
- Williams, S.E.; VanDerWal, J.; Isaac, J.; Shoo, L.P.; Storlie, C.; Fox, S.; Bolitho, E.E.; Moritz, C.; Hoskin, C.J.; Williams, Y.M. Distributions, life-history specialization, and phylogeny of the rain forest vertebrates in the Australian Wet Tropics. Ecology 2010, 91, 2493-2493. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R. Biogeography of insects of New Guinea and Cape York Peninsula. In Bridge and Barrier: The Natural and Cultural History of Torres Strait; Walker, D., Ed.; Australian National University Press: Canberra, Australia, 1972; pp. 213–230. [Google Scholar]
- van Ingen, L.T.; Campos, R.I.; Andersen, A.N. Ant Community Structure along an Extended Rain Forest: Savanna Gradient in Tropical Australia. J. Trop. Ecol. 2008, 445–455. [Google Scholar] [CrossRef]
- Reichel, H.; Andersen, A.N. The rainforest ant fauna of Australia’s Northern Territory. Aust. J. Zool. 1996, 44, 81–95. [Google Scholar] [CrossRef]
- Brühl, C.A.; Gunsalam, G.; Linsenmair, K.E. Stratification of ants (Hymenoptera, Formicidae) in a primary rain forest in Sabah, Borneo. J. Trop. Ecol. 1998, 14, 285–297. [Google Scholar] [CrossRef]
- Longino, J.T.; Coddington, J.; Colwell, R.K. The ant fauna of a tropical rain forest: Estimating species richness three different ways. Ecology 2002, 83, 689–702. [Google Scholar] [CrossRef]
- Andersen, A.N. The Ants of Northern Australia: A Guide to the Monsoonal Fauna; CSIRO Publishing: Clayton, Australia, 2000. [Google Scholar]
- Staunton, K.M.; Robson, S.K.; Burwell, C.J.; Reside, A.E.; Williams, S.E. Projected distributions and diversity of flightless ground beetles within the Australian Wet Tropics and their environmental correlates. PLoS ONE 2014, 9, e88635. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N.; Hoffmann, B.D.; Oberprieler, S.K. Integrated morphological, CO1 and distributional analysis confirms many species in the Iridomyrmex anceps (Roger) complex of ants. Zool. Syst. 2020, 45, 219–230. [Google Scholar]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree: Molecular Evolution, Phylogenetics and Epidemiology. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 May 2020).
- Smith, M.A.; Fisher, B.L.; Hebert, P.D. DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: The ants of Madagascar. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Wild, A.L. Evolution of the Neotropical ant genus Linepithema. Syst. Entomol. 2009, 34, 49–62. [Google Scholar] [CrossRef]
- Schär, S.; Talavera, G.; Espadaler, X.; Rana, J.D.; Andersen Andersen, A.; Cover, S.P.; Vila, R. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. J. Biogeogr. 2018, 45, 1917–1928. [Google Scholar] [CrossRef]
- Wagner, H.C.; Gamisch, A.; Arthofer, W.; Moder, K.; Steiner, F.M.; Schlick-Steiner, B.C. Evolution of morphological crypsis in the Tetramorium caespitum ant species complex (Hymenoptera: Formicidae). Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander Jr, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- van Proosdij, A.S.; Sosef, M.S.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.; Li, J.; Peterson, A.T.; Graham, C.; Guisan, A.; Group, N.P.S.D.W. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Papeş, M.; Gaubert, P. Modelling ecological niches from low numbers of occurrences: Assessment of the conservation status of poorly known viverrids (Mammalia, Carnivora) across two continents. Divers. Distrib. 2007, 13, 890–902. [Google Scholar] [CrossRef]
- Storlie, C.; Phillips, B.; VanDerWal, J.; Williams, S. Improved spatial estimates of climate predict patchier species distributions. Divers. Distrib. 2013, 19, 1106–1113. [Google Scholar] [CrossRef]
- McMahon, J.; Hutchinson, M.; Nix, H.; Ord, K. ANUCLIM User’s Guide. Version 1; Centre for Resource and Environmental Studies, Australian National University: Canberra, Australia, 1995. [Google Scholar]
- VanDerWal, J.; Shoo, L.P.; Johnson, C.N.; Williams, S.E. Abundance and the Environmental Niche: Environmental Suitability Estimated from Niche Models Predicts the Upper Limit of Local Abundance. Am. Nat. 2009, 174, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Nowrouzi, S.; Bush, A.; Harwood, T.; Staunton, K.M.; Robson, S.K.; Andersen, A.N. Incorporating habitat suitability into community projections: Ant responses to climate change in the Australian Wet Tropics. Divers. Distrib. 2019, 25, 1273–1288. [Google Scholar] [CrossRef]
- Brandt, L.A.; Benscoter, A.M.; Harvey, R.; Speroterra, C.; Bucklin, D.; Romañach, S.S.; Watling, J.I.; Mazzotti, F.J. Comparison of climate envelope models developed using expert-selected variables versus statistical selection. Ecol. Model. 2017, 345, 10–20. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitao, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Naimi, B.; Hamm, N.A.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef] [Green Version]
- Longino, J.T.; Branstetter, M.G. The truncated bell: An enigmatic but pervasive elevational diversity pattern in Middle American ants. Ecography 2019, 42, 272–283. [Google Scholar] [CrossRef]
- Longino, J.T.; Branstetter, M.G.; Colwell, R.K. How ants drop out: Ant abundance on tropical mountains. PLoS ONE 2014, 9, e104030. [Google Scholar] [CrossRef] [Green Version]
- Burwell, C.J.; Nakamura, A. Can changes in ant diversity along elevational gradients in tropical and subtropical Australian rainforests be used to detect a signal of past lowland biotic attrition? Austral Ecol. 2015, 41, 209–218. [Google Scholar] [CrossRef]
- Prebus, M. Insights into the evolution, biogeography and natural history of the acorn ants, genus Temnothorax Mayr (hymenoptera: Formicidae). BMC Evol. Biol. 2017, 17, 250. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Blackburn, T.M.; Lawton, J.H. Interspecific abundance-range size relationships: An appraisal of mechanisms. J. Anim. Ecol. 1997, 66, 579–601. [Google Scholar] [CrossRef] [Green Version]
- Storlie, C.; Merino-Viteri, A.; Phillips, B.; VanDerWal, J.; Welbergen, J.; Williams, S. Stepping inside the niche: Microclimate data are critical for accurate assessment of species’ vulnerability to climate change. Biol. Lett. 2014, 10, 20140576. [Google Scholar] [CrossRef] [Green Version]
- Scheffers, B.R.; Phillips, B.L.; Laurance, W.F.; Sodhi, N.S.; Diesmos, A.; Williams, S.E. Increasing arboreality with altitude: A novel biogeographic dimension. Proc. R. Soc. B 2013, 280, 20131581. [Google Scholar] [CrossRef]
- Kaspari, M.; Clay, N.A.; Lucas, J.; Yanoviak, S.P.; Kay, A. Thermal adaptation generates a diversity of thermal limits in a rainforest ant community. Glob. Chang. Biol. 2015, 21, 1092–1102. [Google Scholar] [CrossRef]
- Janzen, D.H.; Schoener, T.W. Differences in insect abundance and diversity between wetter and drier sites during a tropical dry season. Ecology 1968, 49, 96–110. [Google Scholar] [CrossRef]
- Lowman, M.D. Seasonal variation in insect abundance among three Australian rain forests, with particular reference to phytophagous types. Aust. J. Ecol. 1982, 7, 353–361. [Google Scholar] [CrossRef]
- Forster, P.I.; Hyland, B.P.M. Two new species of Eucryphia Cav.(Cunoniaceae) from Queensland. Austrobaileya 1997, 4, 589–596. [Google Scholar]
- Aristophanous, M. Understanding Patterns of Endemic Dung Beetle (Coleoptera: Scarabaeidae: Scarabaeinae) Biodiversity in the Australian Wet Tropics Rainforest: Implications of Climate Change; James Cook University: Townsville, Australia, 2014. [Google Scholar]
- Yeates, D.; Bouchard, P.; Monteith, G. Patterns and levels of endemism in the Australian Wet Tropics rainforest: Evidence from flightless insects. Invertebr. Syst. 2002, 16, 605–619. [Google Scholar] [CrossRef]
- Wilson, R.D.; Trueman, J.W.; Williams, S.E.; Yeates, D.K. Altitudinally restricted communities of Schizophoran flies in Queensland’s Wet Tropics: Vulnerability to climate change. Biodivers. Conserv. 2007, 16, 3163–3177. [Google Scholar] [CrossRef]
- Levins, R.; Pressick, M.L.; Heatwole, H. Coexistence Patterns in Insular Ants: In which it is shown that ants travel a lot from island to island, and what they do when they get there. Am. Sci. 1973, 61, 463–472. [Google Scholar]
- Pianka, E. Latitudinal Gradients in Species Diversity: A Review of Concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Williams, S.E. New approaches to understanding late Quaternary climate fluctuations and refugial dynamics in Australian wet tropical rain forests. J. Biogeogr. 2009, 36, 291–301. [Google Scholar] [CrossRef]
- Williams, S.E.; Bolitho, E.E.; Fox, S. Climate change in Australian tropical rainforests: An impending environmental catastrophe. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1887–1892. [Google Scholar] [CrossRef]
- Williams, S.E.; Shoo, L.P.; Isaac, J.L.; Hoffmann, A.A.; Langham, G. Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biol. 2008, 6, e235. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Edwards, D.P.; Macdonald, S.L.; Senior, R.A.; Andriamahohatra, L.R.; Roslan, N.; Rogers, A.M.; Haugaasen, T.; Wright, P.; Williams, S.E. Extreme thermal heterogeneity in structurally complex tropical rain forests. Biotropica 2017, 49, 35–44. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Shoo, L.; Phillips, B.; Macdonald, S.L.; Anderson, A.; VanDerWal, J.; Storlie, C.; Gourret, A.; Williams, S.E. Vertical (arboreality) and horizontal (dispersal) movement increase the resilience of vertebrates to climatic instability. Glob. Ecol. Biogeogr. 2017, 26, 787–798. [Google Scholar] [CrossRef]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.M.; Jess, M.; Williams, S.E. Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BMC Position | North | North | North | South | South | |
|---|---|---|---|---|---|---|
| Region | Finnegan | Windsor | Carbine | Atherton | Spec | |
| (200–800) | (900–1300) | (100–1200) | (100–1000) | (350–1000) | ||
| Species | n | |||||
| gilberti | 96 | 200–800 | 900–1300 | 100–1200 | 100–1000 | 350–1000 |
| sp. G | 50 | 200–800 | 900–1300 | 100–1200 | 400–1000 | 350–800 |
| sp. B | 1 | 200 | ||||
| sp. C | 22 | 1100–1300 | 1000–1200 | |||
| malandana | 13 | 1100–1300 | 1200 | 1000 | ||
| sp. E | 8 | 900–1100 | ||||
| sp. K | 7 | 900–1100 | 1000 | 800 | 800 | |
| sp. H | 6 | 1300 | 1200 | |||
| sp. F | 1 | 1100 | ||||
| sp. Q | 1 | 900 | ||||
| sp. T | 1 | 1100 | ||||
| sp. L | 1 | 1200 | ||||
| sp. P | 1 | 1000 | ||||
| sp. J | 4 | 200–1000 | ||||
| sp. O | 4 | 400–800 | ||||
| sp. I | 2 | 800–1000 | ||||
| sp. N | 1 | 1000 | ||||
| sp. R | 1 | 600 | ||||
| sp. S | 1 | 1000 | ||||
| sp. U | 1 | 800 | ||||
| sp. M | 11 | 800–1000 | ||||
| sp. A | 10 | 350–1000 |
| Species | n | AUC | Variable | Permutation Importance |
|---|---|---|---|---|
| gilberti | 96 | 0.798 | Max T hottest month | 31.2 |
| Rain wettest month | 21.8 | |||
| Rain seasonality | 22.6 | |||
| sp. G | 50 | 0.754 | Max T hottest month | 64.4 |
| Rain seasonality | 12.3 | |||
| Rain wettest month | 11.4 | |||
| sp. C | 22 | 0.971 | Max T hottest month | 95.7 |
| Isothermality | 3 | |||
| Rain wettest month | 0.7 | |||
| malandana | 13 | 0.958 | Max T hottest month | 82.6 |
| Rain wettest month | 14.7 | |||
| Rain seasonality | 2.4 | |||
| sp. M | 11 | 0.992 | Rain seasonality | 48.9 |
| Max T hottest month | 39.4 | |||
| Rain wettest month | 10.5 | |||
| sp. E | 10 | 0.948 | Max T hottest month | 72 |
| Rain wettest month | 13.5 | |||
| Isothermality | 11.9 | |||
| sp. A | 8 | 0.971 | Rain seasonality | 73.4 |
| Max T hottest month | 23.7 | |||
| Temp seasonality | 2.8 | |||
| sp. K | 7 | 0.892 | Max T hottest month | 95.9 |
| Rain wettest month | 2.1 | |||
| Isothermality | 1.3 | |||
| sp. H | 6 | 0.99 | Max T hottest month | 98.7 |
| Rain seasonality | 1.1 | |||
| Isothermality | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leahy, L.; Scheffers, B.R.; Williams, S.E.; Andersen, A.N. Diversity and Distribution of the Dominant Ant Genus Anonychomyrma (Hymenoptera: Formicidae) in the Australian Wet Tropics. Diversity 2020, 12, 474. https://doi.org/10.3390/d12120474
Leahy L, Scheffers BR, Williams SE, Andersen AN. Diversity and Distribution of the Dominant Ant Genus Anonychomyrma (Hymenoptera: Formicidae) in the Australian Wet Tropics. Diversity. 2020; 12(12):474. https://doi.org/10.3390/d12120474
Chicago/Turabian StyleLeahy, Lily, Brett R. Scheffers, Stephen E. Williams, and Alan N. Andersen. 2020. "Diversity and Distribution of the Dominant Ant Genus Anonychomyrma (Hymenoptera: Formicidae) in the Australian Wet Tropics" Diversity 12, no. 12: 474. https://doi.org/10.3390/d12120474