Reproduction in Urbanised Coastal Waters: Shallow-Water Sea Anemones (Entacmaea quadricolor and Stichodactyla haddoni) Maintain High Genetic Diversity and Panmixia

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
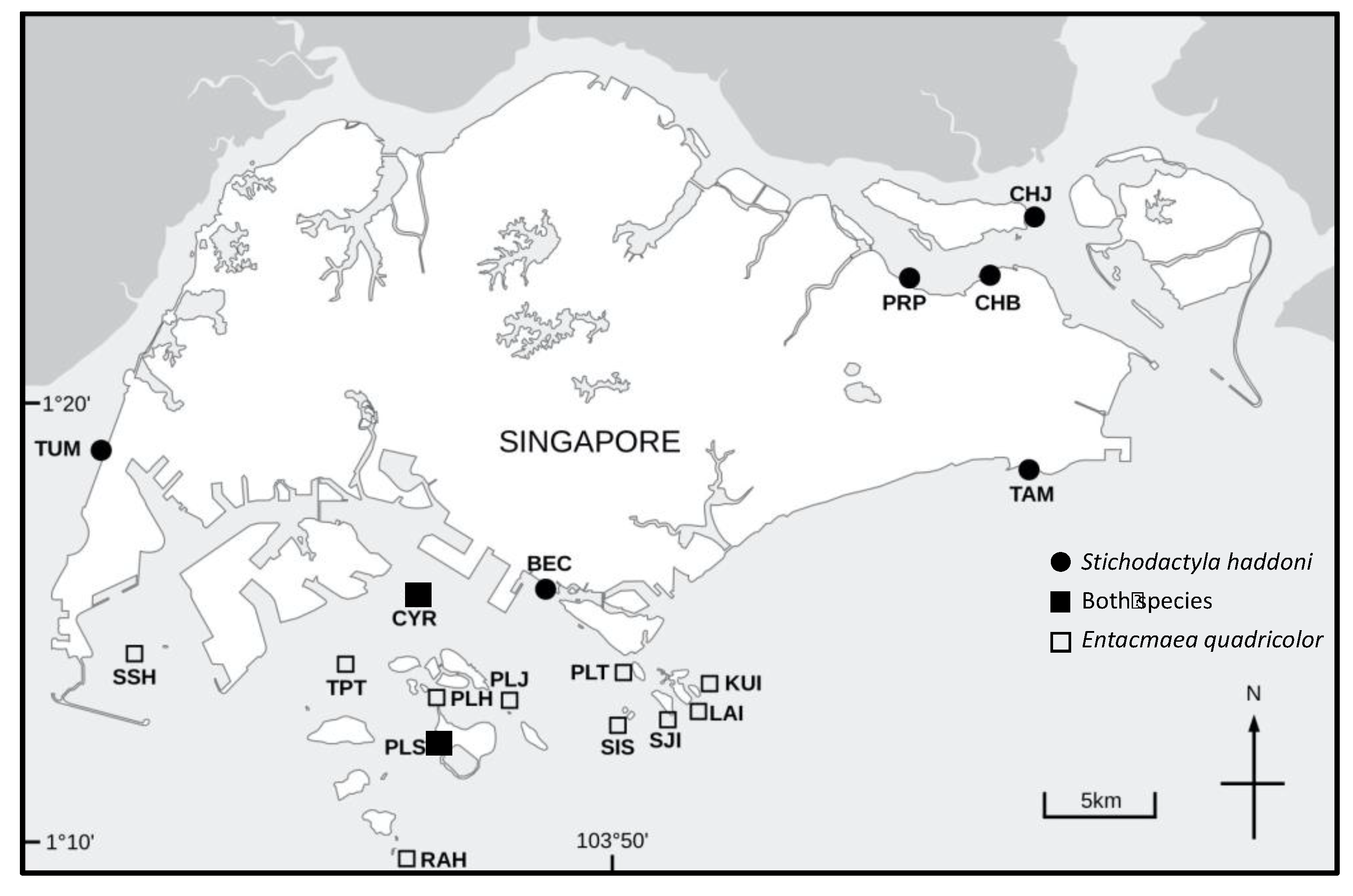
2.1. Sample Collection
2.2. DNA Extraction
2.3. DNA Barcoding for Species Confirmation
2.4. ddRADseq Library Preparation and Sequencing
2.5. Population Genomic Analyses
2.5.1. Single-Nucleotide Polymorphism Data
2.5.2. Population Genomic Structure and Diversity Analyses
3. Results
3.1. DNA Barcoding for Species Confirmation
3.2. Population Genomic Analyses
3.2.1. Quality Filtering of Data and SNP Calling
3.2.2. Population Genomic Structure
3.2.3. Diversity and Reproduction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Goodsell, P.J. Diversity in fragments of artificial and natural marine habitats. Mar. Ecol. Prog. Ser. 2009, 384, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Cowen, R.K.; Lwiza, K.M.; Sponaugle, S.; Paris, C.B.; Olson, D.B. Connectivity of marine populations: Open or closed? Science 2000, 287, 857–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, J.; Hare, J.A.; Sponaugle, S.U. Larval transport and dispersal in the coastal ocean and consequences for population connectivity. Oceanography 2007, 20, 22–39. [Google Scholar] [CrossRef]
- Scott, A.; Harrison, P.L. Broadcast spawning of two species of sea anemone Entacmaea quadricolor and Heteractis crispa that host anemonefish. Invertebr. Reprod. Dev. 2007, 50, 163–171. [Google Scholar] [CrossRef]
- Cowen, R.K.; Sponaugle, S. Larval dispersal and marine population connectivity. Ann. Rev. Mar. Sci. 2009, 1, 443–466. [Google Scholar] [CrossRef] [Green Version]
- Shanks, A.L. Pelagic larval duration and dispersal distance revisited. Biol. Bull. 2009, 216, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Fauvelot, C.; Bertozzi, F.; Costantini, F.; Airoldi, L.; Abbiati, M. Lower genetic diversity in the limpet Patella caerulea on urban coastal structures compared to natural rocky habitats. Mar. Biol. 2009, 156, 2313–2323. [Google Scholar] [CrossRef] [Green Version]
- Fauvelot, C.; Costantini, F.; Virgilio, M.; Abbiati, M. Do artificial structures alter marine invertebrate genetic makeup? Mar. Biol. 2012, 159, 2797–2807. [Google Scholar] [CrossRef]
- Marshall, D.J.; Monro, K.; Bode, M.; Keough, M.J.; Swearer, S. Phenotype–environment mismatches reduce connectivity in the sea. Ecol. Lett. 2010, 13, 128–140. [Google Scholar] [CrossRef]
- Hummel, H.; Patarnello, T. Genetics and pollution. In Genetics and Evolution of Aquatic Organisms; Chapman & Hall: London, UK, 1994; pp. 425–434. [Google Scholar]
- Guzmán, H.M.; Holst, I. Effects of chronic oil-sediment pollution on the reproduction of the Caribbean reef coral Siderastrea sidereal. Mar. Pollut. Bull. 1993, 26, 276–282. [Google Scholar] [CrossRef]
- van Oppen, M.J.; Gates, R.D. Conservation genetics and the resilience of reef-building corals. Mol. Ecol. 2006, 15, 3863–3883. [Google Scholar] [CrossRef]
- Lowe, W.H.; Allendorf, F.W. What can genetics tell us about population connectivity? Mol. Ecol. 2010, 19, 3038–3051. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J.M.; Connolly, S.R.; Marshall, D.J.; Cohen, A.L. Projecting coral reef futures under global warming and ocean acidification. Science 2011, 333, 418–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevo, E.; Noy, R.; Lavie, B.; Beiles, A.; Muchtar, S. Genetic diversity and resistance to marine pollution. Biol. J. Linn. Soc. Lond. 1986, 29, 139–144. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. USA 2004, 101, 8998–9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, A.; Worm, B.; Reusch, T.B. Importance of genetic diversity in eelgrass Zostera marina for its resilience to global warming. Mar. Ecol. Prog. Ser. 2008, 355, 1–7. [Google Scholar] [CrossRef]
- Beger, M.; Selkoe, K.A.; Treml, E.; Barber, P.H.; Von Der Heyden, S.; Crandall, E.D.; Toonen, R.J.; Riginos, C. Evolving coral reef conservation with genetic information. Bull. Mar. Sci. 2014, 90, 159–185. [Google Scholar] [CrossRef] [Green Version]
- Mills, L.S.; Allendorf, F.W. The one-migrant-per-generation rule in conservation and management. Conserv. Biol. 1996, 10, 1509–1518. [Google Scholar] [CrossRef]
- Monkkonen, M.; Reunanen, P. On critical thresholds in landscape connectivity: A management perspective. Oikos 1999, 84, 302–305. [Google Scholar] [CrossRef]
- Drechsler, M.; Frank, K.; Hanski, I.; O’Hara, R.B.; Wissel, C. Ranking metapopulation extinction risk: From patterns in data to conservation management decisions. Ecol. Appl. 2003, 13, 990–998. [Google Scholar] [CrossRef]
- Dunn, D.F. The clownfish sea anemones: Stichodactylidae (Coelenterata: Actiniaria) and other sea anemones symbiotic with pomacentrid. Trans. Am. Philos. Soc. 1981, 71, 1–115. [Google Scholar] [CrossRef]
- Fautin, D.G.; Allen, G.R. Field Guide to Anemonefishes and Their Host Sea Anemones; Sea Challengers: Monterey, CA, USA; Western Australian Museum: Perth, Australia, 1992. [Google Scholar]
- Riemann-Zürneck, K. How sessile are sea anemones? A review of free-living forms in the Actiniaria Cnidaria: Anthozoa. Mar. Ecol. 1998, 19, 247–261. [Google Scholar] [CrossRef]
- Ayre, D.J. Localised adaptation of sea anemone clones: Evidence from transplantation over two spatial scales. J. Anim. Ecol. 1995, 64, 186–196. [Google Scholar] [CrossRef]
- Hoffmann, R.J. Variation in contributions of asexual reproduction to the genetic structure of populations of the sea anemone Metridium senile. Evolution 1986, 40, 357–365. [Google Scholar] [CrossRef]
- Ayre, D.J.; Read, J.; Wishart, J. Genetic subdivision within the eastern Australian population of the sea anemone Actinia tenebrosa. Mar. Biol. 1991, 109, 379–390. [Google Scholar] [CrossRef]
- Chia, F.S. Sea anemone reproduction: Patterns and adaptive radiations. In Coelenterate Ecology and Behavior; Springer: Boston, MA, USA, 1976; pp. 261–270. [Google Scholar]
- Bocharova, E.S.; Kozevich, I.A. Modes of reproduction in sea anemones (Cnidaria Anthozoa). Biol. Bull. 2011, 38, 849–860. [Google Scholar] [CrossRef]
- Hunt, A.; Ayre, D.J. Population structure in the sexually reproducing sea anemone Oulactis muscosa. Mar. Biol. 1989, 102, 537–544. [Google Scholar] [CrossRef]
- Russo, C.A.M.; Solé-Cava, A.M.; Thorpe, J.P. Population structure and genetic variation in two tropical sea anemones (Cnidaria Actinidae) with different reproductive strategies. Mar. Biol. 1994, 119, 267–276. [Google Scholar] [CrossRef]
- Gatins, R. Fine-Scale Population Structure of Two Anemones (Stichodactyla gigantea and Heteractis magnifica) in Kimbe Bay Papua New Guinea. Master’s Thesis, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia, 2014. [Google Scholar]
- Black, R.; Johnson, M.S. Asexual viviparity and population genetics of Actinia tenebrosa. Mar. Biol. 1979, 53, 27–31. [Google Scholar] [CrossRef]
- Waller, R.G. Deep-water Scleractinia (Cnidaria: Anthozoa): Current knowledge of reproductive processes. In Cold-Water Corals and Ecosystems; Springer: Berlin/Heidelberg, Germany, 2005; pp. 691–700. [Google Scholar]
- Sherman, C.D.; Ayre, D.J. Fine-scale adaptation in a clonal sea anemone. Evolution 2008, 62, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Veale, A.J.; Lavery, S.D. The population genetic structure of the waratah anemone (Actinia tenebrosa) around New Zealand. N. Z. J. Mar. Freshw. Res. 2012, 46, 523–536. [Google Scholar] [CrossRef]
- Jackson, J.B.C. Modes of dispersal of clonal benthic invertebrates: Consequences for species’ distributions and genetic structure of local populations. Bull. Mar. Sci. 1986, 39, 588–606. [Google Scholar]
- Fautin, D.G.; Allen, G.R. Anemonefishes and Their Host Sea Anemones: A Guide for Aquarists and Divers; Western Australian Museum: Perth, Australia, 1997. [Google Scholar]
- Scott, A.; Harrison, P.L. Synchronous spawning of host sea anemones. Coral Reefs 2005, 24, 208. [Google Scholar] [CrossRef]
- Scott, A.; Francisco, B. Observations on the feeding behaviour of resident anemonefish during host sea anemone spawning. Coral Reefs 2006, 25, 451. [Google Scholar] [CrossRef]
- Scott, A. Sea anemones. In Marine Ornamental Species Aquaculture; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 437–456. [Google Scholar]
- Sebens, K.P. Morphological variability during longitudinal fission of the intertidal sea anemone Anthopleura elegantissima (Brandt). Pac. Sci. 1983, 37, 121–132. [Google Scholar]
- Holbrook, S.J.; Schmitt, R.J. Growth reproduction and survival of a tropical sea anemone (Actiniaria): Benefits of hosting anemonefish. Coral Reefs 2005, 24, 67–73. [Google Scholar] [CrossRef]
- Edmands, S.; Potts, D.C. Population genetic structure in brooding sea anemones (Epiactis spp.) with contrasting reproductive modes. Mar. Biol. 1997, 127, 485–498. [Google Scholar] [CrossRef]
- Reitzel, A.M.; Herrera, S.; Layden, M.J.; Martindale, M.Q.; Shank, T.M. Going where traditional markers have not gone before: Utility of and promise for RAD sequencing in marine invertebrate phylogeography and population genomics. Mol. Ecol. 2013, 22, 2953–5970. [Google Scholar] [CrossRef] [Green Version]
- Bellis, E.S.; Howe, D.K.; Denver, D.R. Genome-wide polymorphism and signatures of selection in the symbiotic sea anemone Aiptasia. BMC Genom. 2016, 17, 160. [Google Scholar] [CrossRef] [Green Version]
- Shearer, T.L.; Van Oppen, M.J.H.; Romano, S.L.; Wörheide, G. Slow mitochondrial DNA sequence evolution in the Anthozoa (Cnidaria). Mol. Ecol. 2002, 11, 2475–2487. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Meier, R.; Todd, P.A.; Chou, L.M. Slow mitochondrial COI sequence evolution at the base of the metazoan tree and its implications for DNA barcoding. J. Mol. Evol. 2008, 66, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.R.; Good, J.M.; Miller, M.R.; Luikart, G.; Hohenlohe, P.A. Harnessing the power of RADseq for ecological and evolutionary genomics. Nat. Rev. Genet. 2016, 17, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.; Harrison, P.L. Gametogenic and reproductive cycles of the sea anemone Entacmaea quadricolor. Mar. Biol. 2009, 156, 1659. [Google Scholar] [CrossRef]
- Todd, P.A.; Chou, L.M. A tale of survival: Labrador Park, Singapore. Coral Reefs 2005, 24, 391. [Google Scholar] [CrossRef]
- Lai, S.; Loke, L.H.; Hilton, M.J.; Bouma, T.J.; Todd, P.A. The effects of urbanisation on coastal habitats and the potential for ecological engineering: A Singapore case study. Ocean. Coast. Manag. 2015, 103, 78–85. [Google Scholar] [CrossRef]
- Chia, L.S.; Khan, H.; Chou, L.M. The Coastal Environmental Profile of Singapore; ICLARM Technical Reports 21; International Center for Living Aquatic Resources Management: Manila, Philippines, 1988; pp. 1–92. [Google Scholar]
- Yong, K.Y.; Lee, S.L.; Karunaratne, G.P. Coastal reclamation in Singapore: A review. Urban coastal area management: The experience of Singapore. ICLARM Conf. Proc. 1991, 25, 59–67. [Google Scholar]
- Tan, K.S.; Acerbi, E.; Lauro, F.M. Marine habitats and biodiversity of Singapore’s coastal waters: A review. Reg. Stud. Mar. Sci. 2016, 8, 340–352. [Google Scholar] [CrossRef]
- Chou, L.M.; Huang, D.; Tan, K.S.; Toh, T.C.; Goh, B.P.; Tun, K. Chapter 24—Singapore. In World Seas: An Environmental Evaluation, 2nd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 539–558. [Google Scholar]
- Chou, L.M. Marine habitats in one of the world’s busiest harbours. In The Environment in Asia Pacific Harbours; Springer: Dordrecht, The Netherlands, 2006; pp. 377–391. [Google Scholar]
- Mashayekhi, S.; Columbus, J.T. Genetic diversity of Allium munzii (Amaryllidaceae), a rare southern California species and implication for its conservation. Biochem. Syst. Ecol. 2015, 59, 91–99. [Google Scholar] [CrossRef]
- Browne, N.K.; Precht, E.; Last, K.S.; Todd, P.A. Photo-physiological costs associated with acute sediment stress events in three near-shore turbid water corals. Mar. Ecol. Prog. Ser. 2014, 502, 129–143. [Google Scholar] [CrossRef] [Green Version]
- MPA Port Statistics. 2020. Available online: https://www.mpa.gov.sg/web/portal/home/maritime-singapore/port-statistics (accessed on 13 April 2020).
- Bhattachan, P.; Qiao, R.; Dong, B. Identification and population genetic comparison of three ascidian species based on mtDNA sequences. Ecol. Evol. 2020, 10, 3758–3768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.D.; van Rooyen, A.; Rašić, G.; Ierodiaconou, D.A.; Gorfine, H.K.; Day, R.; Wong, C.; Weeks, A.R. Contrasting patterns of population connectivity between regions in a commercially important mollusc Haliotis rubra: Integrating population genetics genomics and marine LiDAR data. Mol. Ecol. 2016, 25, 3845–3864. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.C.; Noreen, A.M.E.; Chou, L.M.; Todd, P.A. Genetic connectivity of the broadcast spawning reef coral Platygyra sinensis on impacted reefs, and the description of new microsatellite markers. Coral Reefs 2015, 34, 301–311. [Google Scholar] [CrossRef]
- Thomas, L.; Underwood, J.N.; Adam, A.A.; Richards, Z.T.; Dugal, L.; Miller, K.J.; Gilmour, J.P. Contrasting patterns of genetic connectivity in brooding and spawning corals across a remote atoll system in northwest Australia. Coral Reefs 2020, 39, 55–60. [Google Scholar] [CrossRef]
- Jones, G.P.; Planes, S.; Thorrold, S.R. Coral reef fish larvae settle close to home. Curr. Biol. 2005, 15, 1314–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, Y.C.; Chng, M.W.P.; Sew, W.W.G.; Rheindt, F.E.; Tun, K.P.P.; Meier, R. Beyond the Coral Triangle: High genetic diversity and near panmixia in Singapore’s populations of the broadcast spawning sea star Protoreaster nodosus. R. Soc. Open Sci. 2016, 3, 160253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas-Sánchez, C.A.; Rivera-Madrid, R.; Arias-González, J.E. Small-scale genetic connectivity of bicolor damselfish (Stegastes partitus) recruits in Mexican Caribbean reefs. Coral Reefs 2010, 29, 1023–1033. [Google Scholar] [CrossRef]
- Coolen, J.W.; Boon, A.R.; Crooijmans, R.; Van Pelt, H.; Kleissen, F.; Gerla, D.; Beermann, J.; Luttikhuizen, P.C. Marine stepping-stones: Connectivity of Mytilus edulis populations between offshore energy installations. Mol. Ecol. 2020, 29, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Jorde, P.E.; Knutsen, H.; Espeland, S.H.; Stenseth, N.C. Spatial scale of genetic structuring in coastal cod Gadus morhua and geographic extent of local populations. Mar. Ecol. Prog. Ser. 2007, 343, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, S.S.; Klanten, S.O.; Puritz, J.B.; Toonen, R.J.; Byrne, M. Very fine-scale population genetic structure of sympatric asterinid sea stars with benthic and pelagic larvae: Influence of mating system and dispersal potential. Biol. J. Linn. Soc. Lond. 2013, 108, 821–833. [Google Scholar] [CrossRef] [Green Version]
- Benestan, L.; Gosselin, T.; Perrier, C.; Sainte-Marie, B.; Rochette, R.; Bernatchez, L. RAD genotyping reveals fine-scale genetic structuring and provides powerful population assignment in a widely distributed marine species, the American lobster (Homarus americanus). Mol. Ecol. 2015, 24, 3299–3315. [Google Scholar] [CrossRef]
- Maas, D.L.; Prost, S.; Bi, K.; Smith, L.L.; Armstrong, E.E.; Aji, L.P.; Toha, A.H.A.; Gillespie, R.G.; Becking, L.E. Rapid divergence of mussel populations despite incomplete barriers to dispersal. Mol. Ecol. 2018, 27, 1556–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wort, E.J.; Chapman, M.A.; Hawkins, S.J.; Henshall, L.; Pita, A.; Rius, M.; Williams, S.T.; Fenberg, P.B. Contrasting genetic structure of sympatric congeneric gastropods: Do differences in habitat preference, abundance and distribution matter? J. Biogeogr. 2019, 46, 369–380. [Google Scholar] [CrossRef]
- Kelly, R.P.; Palumbi, S.R. Genetic structure among 50 species of the northeastern Pacific rocky intertidal community. PLoS ONE 2010, 5, e8594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, P.S.; Rand, D.M. Adaptive maintenance of genetic polymorphism in an intertidal barnacle: Habitat-and life-stage-specific survivorship of MPI genotypes. Evolution 2001, 55, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Fautin, D.G.; Tan, R.; Yap, N.W.L.; Hee, T.S.; Crowther, A.; Goodwill, R.; Sanpanich, K.; Tay, Y.C. Sea anemones (Cnidaria: Actiniaria) of Singapore: Shallow-water species known also from the Indian subcontinent. Raffles Bull. Zool. 2015, 31, 44–59. [Google Scholar]
- Richardson, D.L.; Harriott, V.J.; Harrison, P.L. Distribution and abundance of giant sea anemones (Actiniaria) in subtropical eastern Australian waters. Mar. Freshw. Res. 1997, 48, 59–66. [Google Scholar] [CrossRef]
- Scott, A.; Harasti, D.; Davis, T.; Smith, S.D. Southernmost records of the host sea anemone Stichodactyla haddoni and associated commensal shrimps in a climate change hotspot. Mar. Biol. 2014, 45, 145–146. [Google Scholar] [CrossRef]
- Titus, B.M.; Benedict, C.; Laroche, R.; Gusmão, L.C.; Van Deusen, V.; Chiodo, T.; Meyer, C.P.; Berumen, M.L.; Bartholomew, A.; Yanagi, K.; et al. Phylogenetic relationships among the clownfish-hosting sea anemones. Mol. Phylogenet. Evol. 2019, 139, 106526. [Google Scholar] [CrossRef]
- Shuman, C.S.; Hodgson, G.; Ambrose, R.F. Population impacts of collecting sea anemones and anemonefish for the marine aquarium trade in the Philippines. Coral Reefs 2005, 24, 564–573. [Google Scholar] [CrossRef]
- Nakajima, Y.; Nishikawa, A.; Iguchi, A.; Sakai, K. Gene flow and genetic diversity of a broadcast-spawning coral in northern peripheral populations. PLoS ONE 2010, 5, e11149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.; Doyle, J. CTAB DNA extraction in plants. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Stoletzki, N.; Schierwater, B. Genetic and color morph differentiation in the Caribbean sea anemone Condylactis gigantea. Mar. Biol. 2005, 147, 747–754. [Google Scholar] [CrossRef]
- Meier, R.; Wong, W.; Srivathsan, A.; Foo, M. $1 DNA barcodes for reconstructing complex phenomes and finding rare species in specimen-rich samples. Cladistics 2016, 32, 100–110. [Google Scholar] [CrossRef]
- Wang, W.Y.; Srivathsan, A.; Foo, M.; Yamane, S.K.; Meier, R. Sorting specimen-rich invertebrate samples with cost-effective NGS barcodes: Validating a reverse workflow for specimen processing. Mol. Ecol. Res. 2018, 18, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Murali, K.; Khoo, B.C.; Lou, J.; Kumar, K. Circulation modelling in the Strait of Singapore. J. Coast. Res. 2005, 21, 960–972. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Recknagel, H.; Jacobs, A.; Herzyk, P.; Elmer, K.R. Double-digest RAD sequencing using Ion Proton semiconductor platform (ddRADseq-ion) with nonmodel organisms. Mol. Ecol. Res. 2015, 15, 1316–1329. [Google Scholar] [CrossRef]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. A Quality Control Tool for High Throughput Sequence Data; FastQC: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 11 June 2020).
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris, J.R.; Stevens, J.R.; Catchen, J.M. Lost in parameter space: A road map for stacks. Methods Ecol. Evol. 2017, 8, 1360–1373. [Google Scholar] [CrossRef]
- Bongaerts, P.; Riginos, C.; Brunner, R.; Englebert, N.; Smith, S.R.; Hoegh-Guldberg, O. Deep reefs are not universal refuges: Reseeding potential varies among coral species. Sci. Adv. 2017, 3, e1602373. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Gabor, T.M.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Foll, M. BayeScan v2. 1 user manual. Ecology 2012, 20, 1450–1462. [Google Scholar]
- Lischer, H.E.L.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef] [Green Version]
- Meirmans, P.G.; Van Tienderen, P.H. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Chhatre, V.E.; Emerson, K.J. StrAuto: Automation and parallelization of STRUCTURE analysis. BMC Bioinform. 2017, 18, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Ecol. 2004, 4, 137–138. [Google Scholar]
- Willing, E.M.; Dreyer, C.; Van Oosterhout, C. Estimates of genetic differentiation measured by FST do not necessarily require large sample sizes when using many SNP markers. PLoS ONE 2012, 7, e42649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, A.M.; Jørgensen, A.M.; Coolen, J.W.; Jones, D.O.; Svendsen, J.C.; Brabant, R.; Rumes, B.; Degraer, S. The ecology of infrastructure decommissioning in the North Sea: What we need to know and how to achieve it. ICES J. Mar. Sci. 2020, 77, 1109–1126. [Google Scholar] [CrossRef]
- Tan, W.T.; Loke, L.H.; Yeo, D.C.; Tan, S.K.; Todd, P.A. Do Singapore’s seawalls host non-native marine molluscs? Aquat. Invasions 2018, 13, 365–378. [Google Scholar] [CrossRef]
- Loke, L.H.; Bouma, T.J.; Todd, P.A. The effects of manipulating microhabitat size and variability on tropical seawall biodiversity: Field and flume experiments. J. Exp. Mar. Biol. Ecol. 2017, 492, 113–120. [Google Scholar] [CrossRef]
- Ng, C.S.L.; Chen, D.; Chou, L.M. Hard coral assemblages on seawalls in Singapore. Contrib. Mar. Sci. 2012, 2012, 75–79. [Google Scholar]
- Wilson, J.R.; Harrison, P.L. Spawning patterns of scleractinian corals at the Solitary Islands, a high latitude coral community in eastern Australia. Mar. Ecol. Prog. Ser. 2003, 260, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Guest, J.R.; Baird, A.H.; Goh, B.P.L.; Chou, L.M. Multispecific synchronous coral spawning in Singapore. Coral Reefs 2002, 21, 422–423. [Google Scholar] [CrossRef]
- Guest, J.R.; Baird, A.H.; Goh, B.P.L.; Chou, L.M. Seasonal reproduction in equatorial reef corals. Invertebr. Reprod. Dev. 2005, 48, 207–218. [Google Scholar] [CrossRef]
- Tay, Y.C.; Todd, P.A.; Rosshaug, P.S.; Chou, L.M. Simulating the transport of broadcast coral larvae among the Southern Islands of Singapore. Aquat. Biol. 2012, 15, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Pang, W.C. Modelling tidal and monsoon driven currents in the Singapore Strait. Singap. Marit. Port J. 2003, 2003, 151–162. [Google Scholar]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Poll. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef] [PubMed]
- Chow, G.S.E.; Chan, Y.K.S.; Jain, S.S.; Huang, D. Light limitation selects for depth generalists in urbanised reef coral communities. Mar. Environ. Res. 2019, 147, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Goossens, B.; Chikhi, L.; Ancrenaz, M.; Lackman-Ancrenaz, I.; Andau, P.; Bruford, M.W. Genetic signature of anthropogenic population collapse in orang-utans. PLoS Biol. 2006, 4, e25. [Google Scholar] [CrossRef] [Green Version]
- Poquita-Du, R.C.; Quek, Z.B.R.; Jain, S.S.; Schmidt-Roach, S.; Tun, K.; Heery, E.C.; Chou, L.M.; Todd, P.A.; Huang, D. Last species standing: Loss of Pocilloporidae corals associated with coastal urbanization in a tropical city state. Mar. Biodivers. 2019, 49, 1727–1741. [Google Scholar] [CrossRef]
- Ewers, C.; Wares, J.P. Examining an outlier: Molecular diversity in the Cirripedia. Integr. Comp. Biol. 2012, 52, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Chen, Z.; Wei, Z.; Bu, R.; Wu, Z. DNA barcoding revises a misidentification on mossy frog: New record and distribution extension of Theloderma corticale Boulenger, 1903 (Amphibia: Anura: Rhacophoridae). Mitochondrial DNA Part A 2018, 29, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.F.; Dahl, M.; André, C. Lophelia pertusa corals from the Ionian and Barents seas share identical nuclear ITS2 and near-identical mitochondrial genome sequences. BMC Res. Notes 2013, 6, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohna, T.A.; Kochzius, M. Obstacles to molecular species identification in sea anemones (Hexacorallia: Actiniaria) with COI a COI intron and ITS II. Mar. Biodivers. 2016, 46, 291–297. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | CHJ | PRP | TUM | PLS | TAM | BEC | CHB | CYR | ||||
| CHJ | -- | 0.006 | −0.001 | 0.004 | 0.006 | 0.002 | 0 | 0.001 | ||||
| PRP | 0.11 | -- | 0.006 | 0.013 | −0.003 | −0.003 | 0.003 | 0.007 | ||||
| TUM | 0.56 | 0.043 | -- | 0.009 | −0.007 | −0.004 | 0.005 | 0.003 | ||||
| PLS | 0.243 | 0.02 | 0.023 | -- | 0.002 | 0.005 | 0.012 | 0.006 | ||||
| TAM | 0.163 | 0.678 | 0.955 | 0.394 | -- | −0.016 | −0.002 | −0.003 | ||||
| BEC | 0.362 | 0.805 | 0.872 | 0.172 | 1 | -- | −0.002 | 0.002 | ||||
| CHB | 0.534 | 0.243 | 0.03 | 0.003* | 0.731 | 0.698 | -- | 0.002 | ||||
| CYR | 0.377 | 0.046 | 0.153 | 0.058 | 0.77 | 0.291 | 0.137 | -- | ||||
| (b) | PLH | SIS | PLJ | TPT | KUI | RAL | SJI | PLS | SSH | LAI | PLT | CYR |
| PLH | -- | 0 | 0.004 | −0.002 | 0 | −0.001 | −0.004 | −0.003 | 0.002 | 0.004 | 0.002 | 0 |
| SIS | 0.529 | -- | 0.003 | 0.002 | −0.002 | −0.004 | 0.006 | 0.003 | 0.007 | 0.003 | 0.003 | 0.001 |
| PLJ | 0.059 | 0.216 | -- | 0.003 | −0.001 | −0.002 | 0.001 | 0.002 | 0.005 | 0.006 | 0.006 | 0.001 |
| TPT | 0.84 | 0.204 | 0.16 | -- | −0.002 | −0.005 | −0.007 | −0.003 | −0.002 | −0.006 | 0 | −0.005 |
| KUI | 0.443 | 0.711 | 0.546 | 0.699 | -- | −0.008 | −0.002 | −0.019 | 0.002 | 0.001 | −0.007 | −0.005 |
| RAL | 0.59 | 0.675 | 0.697 | 0.79 | 0.887 | -- | −0.008 | −0.012 | 0.002 | −0.009 | 0.002 | −0.002 |
| SJI | 0.796 | 0.14 | 0.472 | 0.92 | 0.567 | 0.92 | -- | −0.006 | −0.001 | −0.002 | 0.003 | 0 |
| PLS | 0.765 | 0.26 | 0.325 | 0.76 | 1 | 0.922 | 0.86 | -- | −0.005 | −0.002 | −0.005 | −0.005 |
| SSH | 0.304 | 0.044 | 0.082 | 0.745 | 0.316 | 0.388 | 0.524 | 0.873 | -- | −0.002 | 0.004 | 0.001 |
| LAI | 0.149 | 0.268 | 0.073 | 0.898 | 0.409 | 0.889 | 0.711 | 0.681 | 0.65 | -- | 0.002 | −0.006 |
| PLT | 0.28 | 0.239 | 0.047 | 0.455 | 0.984 | 0.412 | 0.256 | 0.903 | 0.144 | 0.277 | -- | −0.003 |
| CYR | 0.534 | 0.401 | 0.323 | 0.935 | 0.923 | 0.627 | 0.492 | 0.838 | 0.469 | 0.95 | 0.763 | -- |
| Population | n | Num | Eff_num | Ho | Hs | Ht | H’t | Gis | |
|---|---|---|---|---|---|---|---|---|---|
| S. haddoni | CHJ | 8 | 1.692 | 1.4 | 0.241 | 0.272 | 0.272 | --- | 0.113 |
| PRP | 9 | 1.752 | 1.414 | 0.241 | 0.276 | 0.276 | --- | 0.127 | |
| TUM | 7 | 1.851 | 1.429 | 0.24 | 0.276 | 0.276 | --- | 0.131 | |
| PLS | 10 | 1.811 | 1.426 | 0.232 | 0.278 | 0.278 | --- | 0.165 | |
| TAM | 7 | 1.687 | 1.393 | 0.226 | 0.271 | 0.271 | --- | 0.167 | |
| BEC | 6 | 1.745 | 1.412 | 0.234 | 0.277 | 0.277 | --- | 0.153 | |
| CHB | 17 | 1.887 | 1.422 | 0.229 | 0.268 | 0.268 | --- | 0.145 | |
| CYR | 8 | 1.838 | 1.424 | 0.237 | 0.274 | 0.274 | --- | 0.135 | |
| Overall | 81 | 1.989 | 1.388 | 0.235 | 0.274 | 0.274 | 0.274 | 0.142 | |
| E. quadricolor | PLH | 12 | 1.806 | 1.364 | 0.198 | 0.238 | 0.238 | --- | 0.17 |
| SIS | 13 | 1.78 | 1.367 | 0.204 | 0.242 | 0.242 | --- | 0.158 | |
| PLJ | 8 | 1.704 | 1.351 | 0.183 | 0.236 | 0.236 | --- | 0.225 | |
| TPT | 17 | 1.842 | 1.363 | 0.173 | 0.237 | 0.237 | --- | 0.268 | |
| KUI | 9 | 1.736 | 1.36 | 0.207 | 0.241 | 0.241 | --- | 0.14 | |
| RAL | 3 | 1.504 | 1.331 | 0.199 | 0.245 | 0.245 | --- | 0.186 | |
| SJI | 4 | 1.562 | 1.329 | 0.19 | 0.234 | 0.234 | --- | 0.189 | |
| PLS | 8 | 1.598 | 1.333 | 0.175 | 0.233 | 0.233 | --- | 0.25 | |
| SSH | 8 | 1.684 | 1.347 | 0.179 | 0.235 | 0.235 | --- | 0.241 | |
| LAI | 5 | 1.645 | 1.36 | 0.21 | 0.247 | 0.247 | --- | 0.15 | |
| PLT | 8 | 1.695 | 1.35 | 0.195 | 0.237 | 0.237 | --- | 0.174 | |
| CYR | 7 | 1.697 | 1.352 | 0.196 | 0.238 | 0.238 | --- | 0.176 | |
| Overall | 99 | 2 | 1.2 | 0.192 | 0.239 | 0.238 | 0.238 | 0.194 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, W.W.R.; Tay, Y.C.; Ang, H.P.; Tun, K.; Chou, L.M.; Huang, D.; Meier, R. Reproduction in Urbanised Coastal Waters: Shallow-Water Sea Anemones (Entacmaea quadricolor and Stichodactyla haddoni) Maintain High Genetic Diversity and Panmixia. Diversity 2020, 12, 467. https://doi.org/10.3390/d12120467
Chan WWR, Tay YC, Ang HP, Tun K, Chou LM, Huang D, Meier R. Reproduction in Urbanised Coastal Waters: Shallow-Water Sea Anemones (Entacmaea quadricolor and Stichodactyla haddoni) Maintain High Genetic Diversity and Panmixia. Diversity. 2020; 12(12):467. https://doi.org/10.3390/d12120467
Chicago/Turabian StyleChan, Wan Wen Rochelle, Ywee Chieh Tay, Hui Ping Ang, Karenne Tun, Loke Ming Chou, Danwei Huang, and Rudolf Meier. 2020. "Reproduction in Urbanised Coastal Waters: Shallow-Water Sea Anemones (Entacmaea quadricolor and Stichodactyla haddoni) Maintain High Genetic Diversity and Panmixia" Diversity 12, no. 12: 467. https://doi.org/10.3390/d12120467