The Role of Hybridisation in the Making of the Species-Rich Arctic-Alpine Genus Saxifraga (Saxifragaceae)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Available Studies and the Extent of Hybridisation
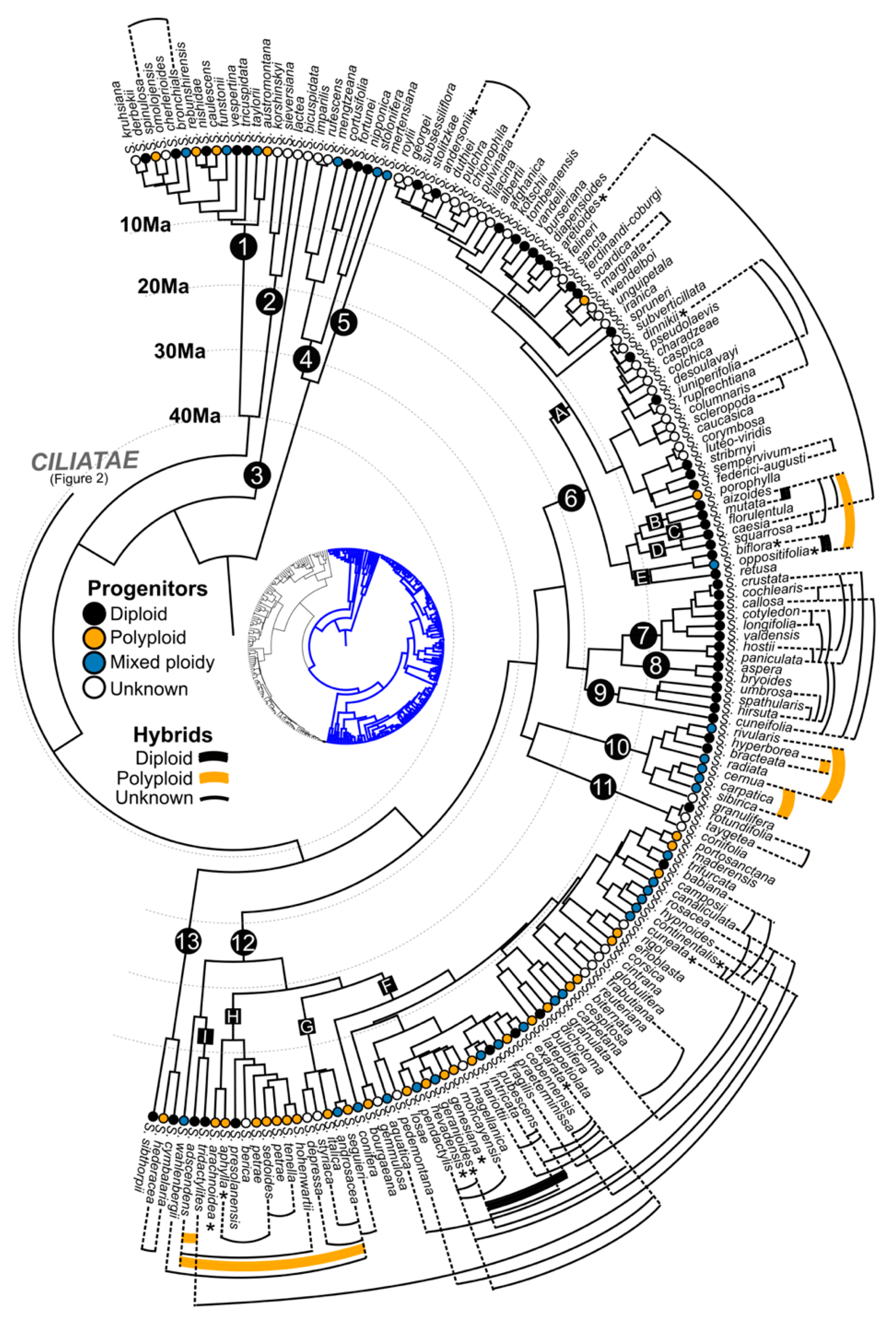
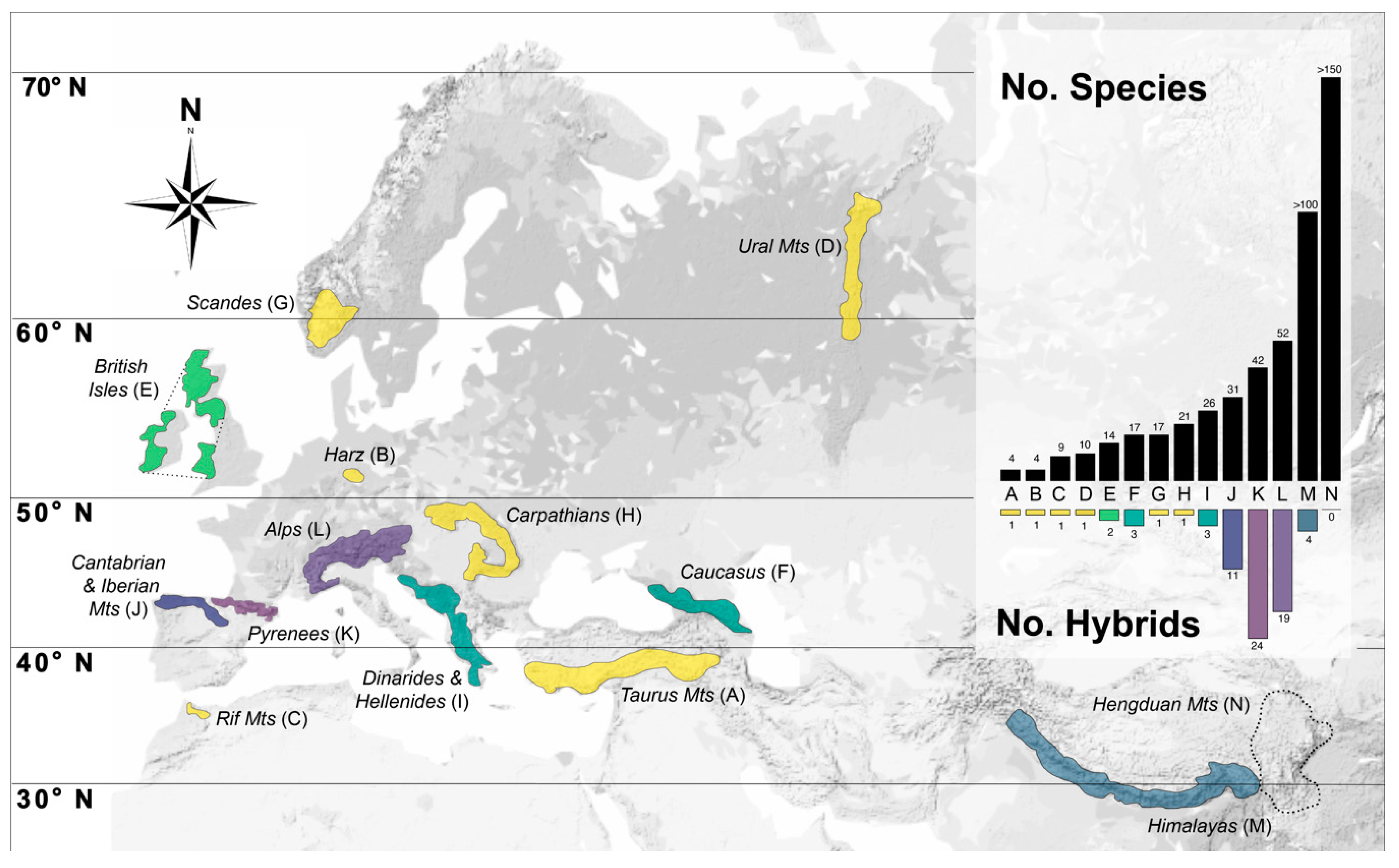
3. Taxonomic, Geographic and Temporal Distribution of Hybridisation in Saxifraga
4. Contemporary Hybrid Zones and Reproductive Isolation
5. Hybrid Speciation
6. Phylogenetic and Geographic Distribution of Ploidy Levels
7. Hybridisation and Polyploidisation as Drivers for Saxifraga Diversity
8. Perspectives in the Context of Mountain Biodiversity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barton, N.H.; Hewitt, G.M. Analysis of hybrid zones. Annual Rev. Ecol. Evol. Syst. 1985, 16, 113–148. [Google Scholar] [CrossRef]
- Arnold, M.L. Natural Hybridization and Evolution; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Wagner, W.H. Biosystematics and evolutionary noise. Taxon 1970, 19, 146–151. [Google Scholar]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA, 1950. [Google Scholar]
- Stebbins, G.L. The role of hybridization in evolution. Proc. Am. Philos. Soc. 1959, 103, 231–251. [Google Scholar]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seehausen, O. Hybridization and adaptive radiation. Trends Ecol. Evol. 2004, 19, 198–207. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. Speciation; Oxford University Press: Sunderland, MA, USA, 2004. [Google Scholar]
- Lowry, D.B.; Modliszewski, J.L.; Wright, K.M.; Wu, C.A.; Willis, J.H. The strength and genetic basis of reproductive isolating barriers in flowering plants. Philos. Trans. Ser. B 2008, 363, 3009–3021. [Google Scholar] [CrossRef] [Green Version]
- Nosil, P.; Vines, T.H.; Funk, D.J. Reproductive isolation caused by natural selection against immigrants from divergent habitats. Evolution 2005, 59, 705–719. [Google Scholar] [CrossRef]
- Sobel, J.M.; Chen, G.F.; Watt, L.R.; Schemske, D.W. The biology of speciation. Evolution 2010, 64, 295–315. [Google Scholar] [CrossRef]
- Mallet, J. Hybrid speciation. Nature 2007, 446, 279–283. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Hybrid origin of plant species. Annual Rev. Ecol. Evol. Syst. 1997, 28, 359–389. [Google Scholar] [CrossRef] [Green Version]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Payseur, B.A.; Rieseberg, L.H. A genomic perspective on hybridization and speciation. Mol. Ecol. 2016, 25, 2337–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto Feliner, G.; Casacuberta, J.; Wendel, J.F. Genomics of evolutionary novelty in hybrids and polyploids. Front. Genet. 2020, 11, 792. [Google Scholar] [CrossRef] [PubMed]
- Tkach, N.; Schneider, J.; Döring, E.; Wölk, A.; Hochbach, A.; Nissen, J.; Winterfeld, G.; Meyer, S.; Gabriel, J.; Hoffmann, M.H.; et al. Phylogenetic lineages and the role of hybridization as driving force of evolution in grass supertribe Poodae. Taxon 2020, 69, 234–277. [Google Scholar] [CrossRef] [Green Version]
- Lexer, C.; Welch, M.E.; Raymond, O.; Rieseberg, L.H. The origin of ecological divergence in Helianthus paradoxus (Asteraceae): Selection on transgressive characters in a novel hybrid habitat. Evolution 2003, 57, 1989–2000. [Google Scholar] [CrossRef]
- Gross, B.L.; Rieseberg, L.H. The ecological genetics of homoploid hybrid speciation. Heredity 2005, 96, 241–252. [Google Scholar] [CrossRef]
- Yakimowski, S.B.; Rieseberg, L.H. The role of homoploid hybridization in evolution: A century of studies synthesizing genetics and ecology. Am. J. Bot. 2014, 101, 1247–1258. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazato, T.; Durphy, J.L.; Schwarzbach, A.E.; Donovan, L.A.; Lexer, C. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 2003, 301, 1211–1216. [Google Scholar] [CrossRef] [Green Version]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [Green Version]
- Abbott, R.J.; Hegarty, M.J.; Hiscock, S.J.; Brennan, A.C. Homoploid hybrid speciation. Taxon 2010, 59, 1375–1386. [Google Scholar]
- Schumer, M.; Rosenthal, G.G.; Andolfatto, P. How common is homoploid hybrid speciation? Evolution 2014, 68, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Nieto Feliner, G.; Álvarez, I.; Fuertes-Aguilar, J.; Heuertz, M.; Marques, I.; Moharrek, F.; Piñeiro, R.; Riina, R.; Rosselló, J.A.; Soltis, P.S.; et al. Is homoploid hybrid speciation that rare? An empiricist’s view. Heredity 2017, 118, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karrenberg, S.; Favre, A. Genetic and ecological differentiation in the hybridizing campions Silene dioica and S. latifolia. Evolution 2008, 62, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Lamont, B.B.; He, T.; Enright, N.J.; Krauss, S.L.; Miller, B.P. Anthropogenic disturbance promotes hybridization between Banksia species by altering their biology. J. Evol. Biol. 2003, 16, 551–557. [Google Scholar] [CrossRef]
- Cruzan, M.B.; Arnold, M.L. Ecological and genetic associations in an Iris hybrid zone. Evolution 1993, 47, 1432–1445. [Google Scholar] [CrossRef]
- Johnston, J.A.; Wesselingh, R.A.; Bouck, A.C.; Donovan, L.A.; Arnold, M.L. Intimately linked or hardly speaking? The relationship between genotype and environmental gradients in a Louisiana Iris hybrid population. Mol. Ecol. 2001, 10, 673–681. [Google Scholar] [CrossRef]
- Brochmann, C.; Brysting, A.K.; Alsos, I.G.; Grundt, H.H.; Scheen, A.-C.; Elven, R. Polyploidy in arctic plants. Biol. J. Linn. Soc. 2004, 82, 521–536. [Google Scholar] [CrossRef] [Green Version]
- Stebbins, G.L. Polyploidy and the distribution of the arctic-alpine flora: New evidence and a new approach. Bot. Helv. 1984, 94, 1–13. [Google Scholar] [CrossRef]
- Rice, A.; Šmarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Löve, Á.; Löve, D. Arctic polyploidy. Proc. Genet. Soc. Canada 1957, 2, 23–27. [Google Scholar]
- Tkach, N.V.; Hoffmann, M.H.; Röser, M.; Korobkov, A.A.; von Hagen, K.B. Parallel evolutionary patterns in multiple lineages of arctic Artemisia L. (Asteraceae). Evolution 2008, 62, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Seehausen, O. Conditions when hybridization might predispose populations for adaptive radiation. J. Evol. Biol. 2013, 26, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.B.; Soltis, D.E.; Li, Z.; Marx, H.E.; Barker, M.S.; Tank, D.C.; Soltis, P.S. Impact of whole-genome duplication events on diversification rates in angiosperms. Am. J. Bot. 2018, 105, 348–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, P.R.; Grant, B.R. Hybridization increases population variation during adaptive radiation. Proc. Natl. Acad. Sci. USA 2019, 116, 23216–23224. [Google Scholar] [CrossRef]
- Körner, C. Coldest places on earth with angiosperm plant life. Alpine Bot. 2011, 121, 11–22. [Google Scholar] [CrossRef]
- Pan, J.; Gornall, R.J.; Ohba, H. Saxifraga L. In Flora of China, vol. 8, Brassicaceae through Saxifragaceae; Wu, C., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2001; pp. 280–344. [Google Scholar]
- Brouillet, L.; Elvander, P.E. Saxifraga L. In Flora of North America North of Mexico, vol. 8, Magnoliophyta: Paeoniaceae to Ericaceae; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA; Oxford, UK, 2009; pp. 43–166. [Google Scholar]
- Akiyama, S.; Gornall, R.J. Saxifraga L. In Flora of Nepal, vol. 3, Magnoliaceae to Rosaceae; Watson, M.F., Akiyama, S., Ikeda, H., Pendry, C.A., Rajbhandari, K.R., Shrestha, K.K., Eds.; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2012; pp. 254–303. [Google Scholar]
- Tkach, N.; Röser, M.; Miehe, G.; Muellner-Riehl, A.N.; Ebersbach, J.; Favre, A.; Hoffmann, M.H. Molecular phylogenetics, morphology and a revised classification of the complex genus Saxifraga (Saxifragaceae). Taxon 2015, 64, 1159–1187. [Google Scholar] [CrossRef]
- Ebersbach, J.; Schnitzler, J.; Favre, A.; Muellner-Riehl, A.N. Evolutionary radiations in the species-rich mountain genus Saxifraga L. BMC Evol. Biol. 2017, 17, 1–13. [Google Scholar]
- Ebersbach, J.; Muellner-Riehl, A.N.; Favre, A.; Paule, J.; Winterfeld, G.; Schnitzler, J. Driving forces behind evolutionary radiations: Saxifraga section Ciliatae (Saxifragaceae) in the region of the Qinghai–Tibet Plateau. Bot. J. Linn. Soc. 2018, 186, 304–320. [Google Scholar]
- McGregor, M. Saxifrages. A Definitive Guide to the 2000 Species, Hybrids & Cultivars; Timber Press: Portland, OR, USA, 2008. [Google Scholar]
- McGregor, M.; Harding, W. Saxifrages: The Complete List of Species; The Saxifrage Society: Hutton, East Yorkshire, UK, 1998. [Google Scholar]
- Webb, D.A.; Gornall, R.J. A Manual of Saxifrages and Their Cultivation, 1st ed.; Timber Press, Incorporated: Portland, OR, USA, 1989. [Google Scholar]
- Brochmann, C.; Nilsson, T.; Gabrielson, T.M. A classic example of postglacial allopolyploid speciation re-examined using RAPD markers and nucleotide sequences: Saxifraga osloensis (Saxifragaceae). Symb. Bot. Upsal. 1996, 31, 75–89. [Google Scholar]
- Nilsson, T.; Jorde, P.E. Allozyme variation in allotetraploid Saxifraga osloensis and its diploid progenitors. Nordic J. Bot. 1998, 18, 425–430. [Google Scholar]
- Tkach, N.; Röser, M.; Suchan, T.; Cieślak, E.; Schönswetter, P.; Ronikier, M. Contrasting evolutionary origins of two mountain endemics: Saxifraga wahlenbergii (Western Carpathians) and S. styriaca (Eastern Alps). BMC Evol. Biol. 2019, 19, 18. [Google Scholar] [CrossRef] [Green Version]
- Ebersbach, J.; Muellner-Riehl, A.N.; Michalak, I.; Tkach, N.; Hoffmann, M.H.; Röser, M.; Sun, H.; Favre, A. In and out of the Qinghai-Tibet Plateau: Divergence time estimation and historical biogeography of the large arctic-alpine genus Saxifraga L. J. Biogeogr. 2017, 44, 900–910. [Google Scholar] [CrossRef]
- Kahle, D.; Wickham, H. ggmap: Spatial visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Zhang, F.; Li, Y.; Gao, Q.; Lei, S.; Khan, G.; Yang, H.; Chen, S. Development and characterization of polymorphic microsatellite loci for Saxifraga egregia (Saxifragaceae). Appl. Plant. Sci. 2015, 3. [Google Scholar] [CrossRef]
- Wakabayashi, M.; Ohba, H. Cytotaxonomic study of the Himalayan Saxifraga. In The Himalayan Plants; Ohba, H., Malla, S.B., Eds.; University Tokyo Press: Tokyo, Japan, 1988; pp. 71–90. [Google Scholar]
- Gao, Q.-B.; Li, Y.; Gengji, Z.-M.; Gornall, R.J.; Wang, J.-L.; Liu, H.-R.; Jia, L.-K.; Chen, S.-L. Population genetic differentiation and taxonomy of three closely related species of Saxifraga (Saxifragaceae) from Southern Tibet and the Hengduan Mountains. Front. Plant. Sci. 2017, 8, 1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xu, Y.; Tang, Y.; Du, W.; Shao, C.; Wu, J.; Zhao, L.; Zhang, L.; Liu, J.; Xu, X. The importance of including soil properties when disentangling the drivers of species richness: The case of the alpine genus Saxifraga L. in China. Front. Ecol. Evol. 2020, 8, 388. [Google Scholar] [CrossRef]
- Bürgel, J. Hybridisation in Saxifraga subsection Kabschia (Saxifragaceae) from the Central Himalaya. Phyton 2007, 47, 191–204. [Google Scholar]
- Smith, H. Saxifraga of the Himalaya: 1. Section Kabschia. Bull. Brit. Mus. (Nat. Hist.) Bot. 1958, 2, 83–129. [Google Scholar]
- Conti, E.; Soltis, D.E.; Hardig, T.M.; Schneider, J. Phylogenetic relationships of the silver saxifrages (Saxifraga, sect. Ligulatae Haworth): Implications for the evolution of substrate specificity, life histories, and biogeography. Mol. Phylogenet. Evol. 1999, 13, 536–555. [Google Scholar]
- DeChaine, E.G.; Anderson, S.A.; McNew, J.M.; Wendling, B.M. On the evolutionary and biogeographic history of Saxifraga sect. Trachyphyllum (Gaud.) Koch (Saxifragaceae Juss.). PLoS ONE 2013, 8, e69814. [Google Scholar] [CrossRef] [Green Version]
- Kay, K.M. Reproductive isolation between two closely related hummingbird-pollinated Neotropical gingers. Evolution 2006, 60, 538–552. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.; Bradshaw, H.D., Jr.; Schemske, D.W. Components of reproductive isolation between the monkeyflowers Mimulus lewisii and M. cardinalis (Phrymaceae). Evolution 2003, 57, 1520–1534. [Google Scholar] [PubMed]
- Schluter, D. Ecology and the origin of species. Trends Ecol. Evol. 2001, 16, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Seehausen, O.; Butlin, R.K.; Keller, I.; Wagner, C.E.; Boughman, J.W.; Hohenlohe, P.A.; Peichel, C.L.; Saetre, G.-P.; Bank, C.; Brännström, A.; et al. Genomics and the origin of species. Nat. Rev. Genet. 2014, 15, 176–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, P.; Nieto Feliner, G. Cytotaxonomical study of Saxifraga sect. Ceratophyllae s.l. (Saxifragaceae). Plant. Syst. Evol. 1995, 197, 209–223. [Google Scholar]
- Mas de Xaxars, G.; García-Fernández, A.; Barnola, P.; Martín, J.; Mercadé, A.; Vallès, J.; Vargas, P.; Vigo, J.; Garnatje, T. Phylogenetic and cytogenetic studies reveal hybrid speciation in Saxifraga subsect. Triplinervium (Saxifragaceae). J. Syst. Evol. 2015, 53, 53–62. [Google Scholar] [CrossRef]
- García-Maroto, F.; Garrido-Cárdenas, J.A.; Gómez-Mercado, F.; Guil-Guerrero, J.; López Alonso, D. Natural hybridisation and phylogenetic position of Saxifraga trabutiana (Saxifragaceae) inferred from ISSR markers and ITS sequences. Ann. Bot. Fenn. 2003, 40, 5–13. [Google Scholar]
- Gustafsson, A.L.S. Speciation in Arctic and Alpine Diploid Plants. Doctoral Thesis, University of Oslo, Oslo, Norway, 2013. [Google Scholar]
- Dymshakova, O.S.; Semerikov, V.L.; Lascoux, M. AFLP analysis to estimate the genetic contribution of parents to progeny from hybridization between Saxifraga sibirica L. and S. cernua L. Russ. J. Ecol. 2012, 43, 347–351. [Google Scholar] [CrossRef]
- Steen, S.W.; Gielly, L.; Taberlet, P.; Brochmann, C. Same parental species, but different taxa: Molecular evidence for hybrid origins of the rare endemics Saxifraga opdalensis and S. svalbardensis (Saxifragaceae). Bot. J. Linn. Soc. 2000, 132, 153–164. [Google Scholar]
- Díaz González, T.E.; Fernández Areces, M.P.; Pérez Carro, J. Nuevos híbridos y otros datos biométricos del genero Saxifraga L., sección Dactyloides Tausch en el NW de la Península Ibérica. Anales Jard. Bol. Madrid. 1990, 47, 65–85. [Google Scholar]
- Gugerli, F. Hybridization of Saxifraga oppositifolia and S. biflora (Saxifragaceae) in a mixed alpine population. Plant. Syst. Evol. 1997, 207, 255–272. [Google Scholar]
- Köckinger, H. Saxifraga styriaca spec. nova (Saxifragaceae)—ein Endemit der östlichen Niederen Tauern (Steiermark, Österreich). Phyton 2003, 43, 79–108. [Google Scholar]
- Sambatti, J.B.M.; Strasburg, J.L.; Ortiz-Barrientos, D.; Baack, E.J.; Rieseberg, L.H. Reconciling extremely strong barriers with high levels of gene exchange in annual sunflowers. Evolution 2012, 66, 1459–1473. [Google Scholar] [CrossRef] [PubMed]
- Hersch-Green, E.I. Polyploidy in Indian paintbrush (Castilleja; Orobanchaceae) species shapes but does not prevent gene flow across species boundaries. Am. J. Bot. 2012, 99, 1680–1690. [Google Scholar] [CrossRef]
- Carrió, E.; Güemes, J. The effectiveness of pre- and post-zygotic barriers in avoiding hybridization between two snapdragons (Antirrhinum L.: Plantaginaceae). Bot. J. Linn. Soc. 2014, 176, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Vargas, P.; Nieto Feliner, G. Artificial hybridization within Saxifraga pentadactylis (Saxifragaceae). Nordic J. Bot. 1996, 16, 257–266. [Google Scholar] [CrossRef]
- Baack, E.; Melo, M.C.; Rieseberg, L.H.; Ortiz-Barrientos, D. The origins of reproductive isolation in plants. New Phytol. 2015, 207, 968–984. [Google Scholar] [CrossRef]
- von Drygalski, U. Über die Entstehung einer tetraploiden, genetisch ungleichmäßigen F2 aus der Kreuzung Saxifraga adscendens L. × Saxifraga tridactylites L. Z. Indukt. Abstammungs-Vererbungsl. 1935, 69, 278–300. [Google Scholar] [CrossRef]
- Favre, A.; Karrenberg, S. Stress tolerance in closely related species and their first-generation hybrids: A case study of Silene. J. Ecol. 2011, 99, 1415–1423. [Google Scholar] [CrossRef]
- Webb, D.A. Hybridization and variation in the Robertsonian saxifrages. Proc. R. Ir. Acad. B 1950, 53, 85–97. [Google Scholar]
- Beatty, G.E.; Barker, L.; Chen, P.-P.; Kelleher, C.T.; Provan, J. Cryptic introgression into the kidney saxifrage (Saxifraga hirsuta) from its more abundant sympatric congener Saxifraga spathularis, and the potential risk of genetic assimilation. Ann. Bot. 2015, 115, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmé, J.; Widmer, A.; Karrenberg, S. Pollen competition as an asymmetric reproductive barrier between two closely related Silene species. J. Evol. Biol. 2009, 22, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Grassi, F.; Labra, M.; Minuto, L.; Casazza, G.; Sala, F. Natural hybridization in Saxifraga callosa Sm. PLoS Biol. 2006, 8, 243–252. [Google Scholar] [CrossRef]
- Kapralov, M.V.; Gabrielsen, T.M.; Sarapultsev, I.E.; Brochmann, C. Genetic enrichment of the arctic clonal plant Saxifraga cernua at its southern periphery via the alpine sexual Saxifraga sibirica. Molec. Ecol. 2006, 15, 3401–3411. [Google Scholar] [CrossRef]
- Suarez-Gonzalez, A.; Lexer, C.; Cronk, Q.C.B. Adaptive introgression: A plant perspective. Biol. Lett. 2018, 14. [Google Scholar] [CrossRef]
- Stankowski, S.; Streisfeld, M.A. Introgressive hybridization facilitates adaptive divergence in a recent radiation of monkeyflowers. Proc. Biol. Sci. 2015, 282, 20151666. [Google Scholar] [CrossRef] [Green Version]
- Leroy, T.; Louvet, J.-M.; Lalanne, C.; Le Provost, G.; Labadie, K.; Aury, J.-M.; Delzon, S.; Plomion, C.; Kremer, A. Adaptive introgression as a driver of local adaptation to climate in European white oaks. New Phytol. 2020, 226, 1171–1182. [Google Scholar] [CrossRef]
- Holderegger, R.; Abbott, R.J. Phylogeography of the arctic-alpine Saxifraga oppositifolia (Saxifragaceae) and some related taxa based on cpDNA and ITS sequence variation. Am. J. Bot. 2003, 90, 931–936. [Google Scholar] [CrossRef]
- Karrenberg, S.; Liu, X.; Hallander, E.; Favre, A.; Herforth-Rahmé, J.; Widmer, A. Ecological divergence plays an important role in strong but complex reproductive isolation in campions (Silene). Evolution 2019, 73, 245–261. [Google Scholar] [CrossRef]
- Brochmann, C.; Xiang, Q.-Y.; Brunsfeld, S.J.; Soltis, D.E.; Soltis, P.S. Molecular evidence of polyploid origins in Saxifraga (Saxifragaceae): The narrow arctic endemic S. svalbardensis and its widespread allies. Am. J. Bot. 1998, 85, 135–143. [Google Scholar]
- Jørgensen, M.H.; Elven, R.; Tribsch, A.; Gabrielsen, T.M.; Brochmann, C. Taxonomy and evolutionary relationships in the Saxifraga rivularis complex. Syst. Bot. 2006, 31, 702–729. [Google Scholar]
- Böcher, T.W. The allotetraploid Saxifraga nathorsti and its probable progenitors S. aizoides and S. oppositifolia; Meddel. Grønland Biosci. 1983, 11, 1–22. [Google Scholar]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar]
- Vallejo-Marín, M.; Buggs, R.J.A.; Cooley, A.M.; Puzey, J.R. Speciation by genome duplication: Repeated origins and genomic composition of the recently formed allopolyploid species Mimulus peregrinus. Evolution 2015, 69, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Kovarík, A.; Zozomová-Lihová, J.; Shimizu-Inatsugi, R.; Shimizu, K.K.; Mummenhoff, K.; Marhold, K.; Lysak, M.A. The more the merrier: Recent hybridization and polyploidy in Cardamine. Plant. Cell 2013, 25, 3280–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapralov, M.V. Genotypic variation in populations of the clonal plant Saxifraga cernua in the central and peripheral regions of the species range. Russ. J. Ecol. 2004, 35, 413–416. [Google Scholar] [CrossRef]
- Eidesen, P.B.; Ehrich, D.; Bakkestuen, V.; Alsos, I.G.; Gilg, O.; Taberlet, P.; Brochmann, C. Genetic roadmap of the Arctic: Plant dispersal highways, traffic barriers and capitals of diversity. New Phytol. 2013, 200, 898–910. [Google Scholar] [CrossRef] [Green Version]
- Müller, E.; Eidesen, P.B.; Ehrich, D.; Alsos, I.G. Frequency of local, regional, and long-distance dispersal of diploid and tetraploid Saxifraga oppositifolia (Saxifragaceae) to Arctic glacier forelands. Am. J. Bot. 2012, 99, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Rieseberg, L.H. Homoploid reticulate evolution in Helianthus (Asteraceae): Evidence from ribosomal genes. Am. J. Bot. 1991, 78, 1218–1237. [Google Scholar] [CrossRef]
- Lai, Z.; Kane, N.C.; Kozik, A.; Hodgins, K.A.; Dlugosch, K.M.; Barker, M.S.; Matvienko, M.; Yu, Q.; Turner, K.G.; Pearl, S.A.; et al. Genomics of Compositae weeds: EST libraries, microarrays, and evidence of introgression. Am. J. Bot. 2012, 99, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.-P.; Abbott, R.J.; Zhou, Y.-F.; Zhang, L.-R.; Peng, Y.-L.; Liu, J.-Q. Genetic divergence, range expansion and possible homoploid hybrid speciation among pine species in Northeast China. Heredity 2012, 108, 552–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.-F.; Wang, X.-R. Distinct niche divergence characterizes the homoploid hybrid speciation of Pinus densata on the Tibetan Plateau. Am. Nat. 2011, 177, 424–439. [Google Scholar] [PubMed] [Green Version]
- Mannocci, M.I.; Feretti, G.; Mazzoncini, V.; Fiorni, G.; Foggi, B.; Lastrucci, L.; Lazzaro, L.; Viciani, D. Two new Saxifraga species (Saxifragaceae) endemic to Tuscan Archipelago (central-northern Mediterranean, Italy). Phytotaxa 2016, 284, 108–130. [Google Scholar] [CrossRef]
- Westergaard, K.B.; Jørgensen, M.H.; Gabrielsen, T.M.; Alsos, I.G.; Brochmann, C. The extreme Beringian/Atlantic disjunction in Saxifraga rivularis (Saxifragaceae) has formed at least twice. J. Biogeogr. 2010, 37, 1262–1276. [Google Scholar] [CrossRef]
- Guldahl, A.S.; Gabrielsen, T.M.; Scheen, A.-C.; Borgen, L.; Steen, S.W.; Spjelkavik, S.; Brochmann, C. The Saxifraga rivularis complex in Svalbard: Molecules, ploidy and morphology. Flora 2005, 200, 207–221. [Google Scholar] [CrossRef]
- de Storme, N.; Mason, A. Plant speciation through chromosome instability and ploidy change: Cellular mechanisms, molecular factors and evolutionary relevance. Curr. Plant Biol. 2014, 1, 10–33. [Google Scholar] [CrossRef] [Green Version]
- Muellner-Riehl, A.N.; Schnitzler, J.; Kissling, W.D.; Mosbrugger, V.; Rijsdijk, K.F.; Seijmonsbergen, A.C.; Versteegh, H.; Favre, A. Origins of global mountain plant biodiversity: Testing the ‘mountain-geobiodiversity hypothesis’. J. Biogeogr. 2019, 46, 2826–2838. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebersbach, J.; Tkach, N.; Röser, M.; Favre, A. The Role of Hybridisation in the Making of the Species-Rich Arctic-Alpine Genus Saxifraga (Saxifragaceae). Diversity 2020, 12, 440. https://doi.org/10.3390/d12110440
Ebersbach J, Tkach N, Röser M, Favre A. The Role of Hybridisation in the Making of the Species-Rich Arctic-Alpine Genus Saxifraga (Saxifragaceae). Diversity. 2020; 12(11):440. https://doi.org/10.3390/d12110440
Chicago/Turabian StyleEbersbach, Jana, Natalia Tkach, Martin Röser, and Adrien Favre. 2020. "The Role of Hybridisation in the Making of the Species-Rich Arctic-Alpine Genus Saxifraga (Saxifragaceae)" Diversity 12, no. 11: 440. https://doi.org/10.3390/d12110440