Seascape Configuration Leads to Spatially Uneven Delivery of Parrotfish Herbivory across a Western Indian Ocean Seascape

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
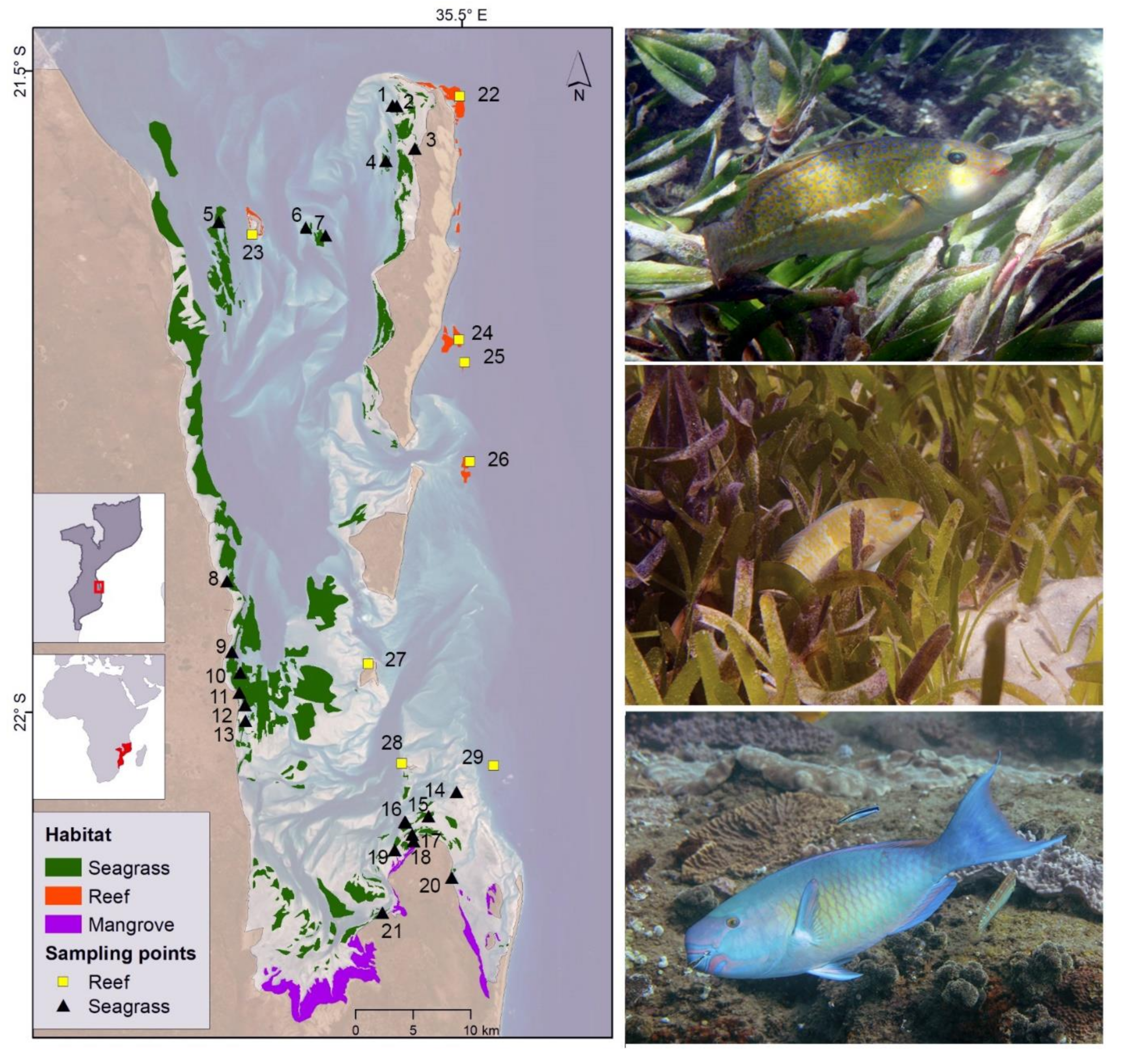
2.1. Study Site
2.2. Fish and Habitat Surveys
2.3. Spatial Metrics
2.4. Bite Rate and Substrate Selectivity
2.5. Data Analyses
3. Results
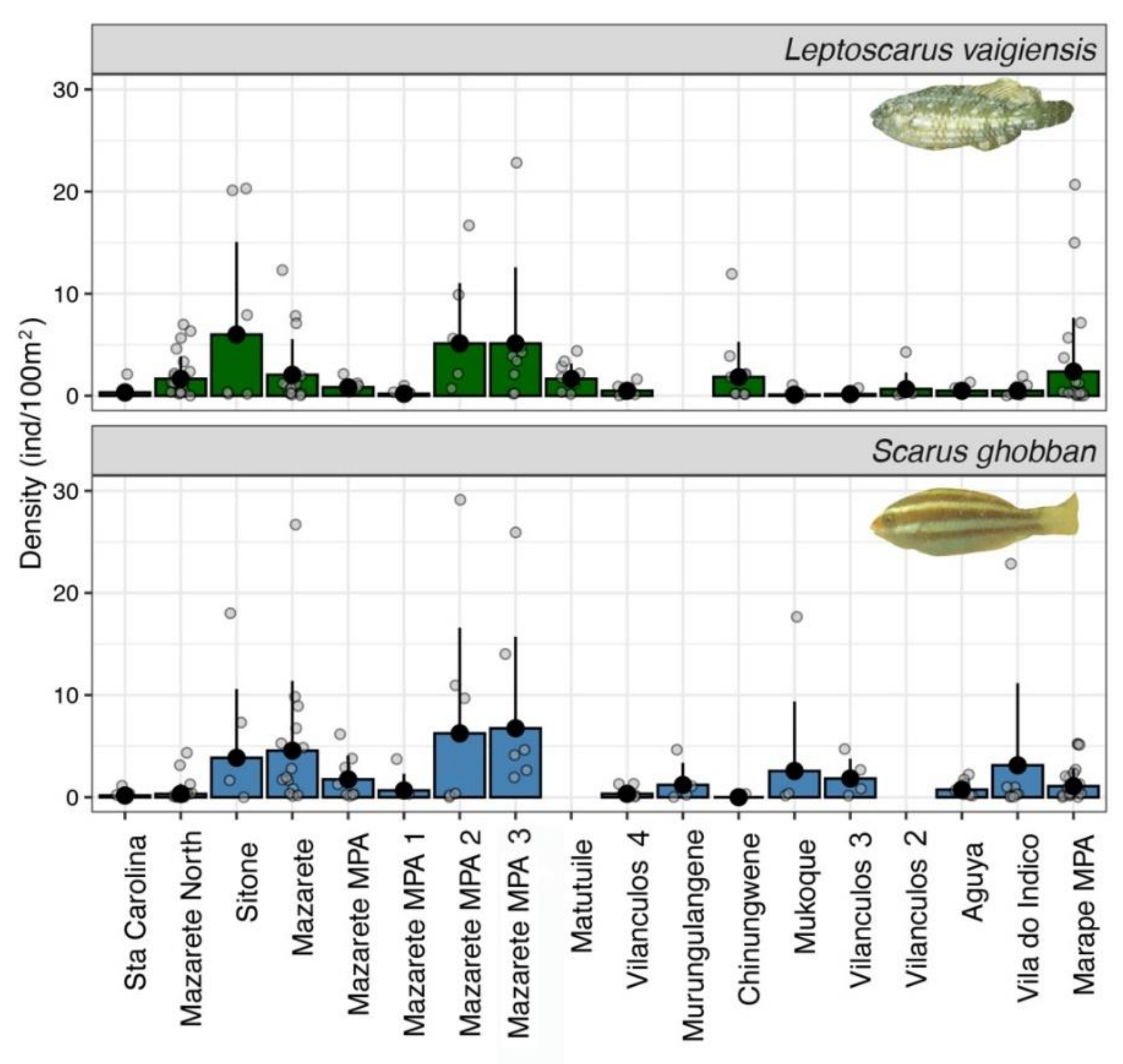
3.1. Fish Abundance and Biomass Patterns
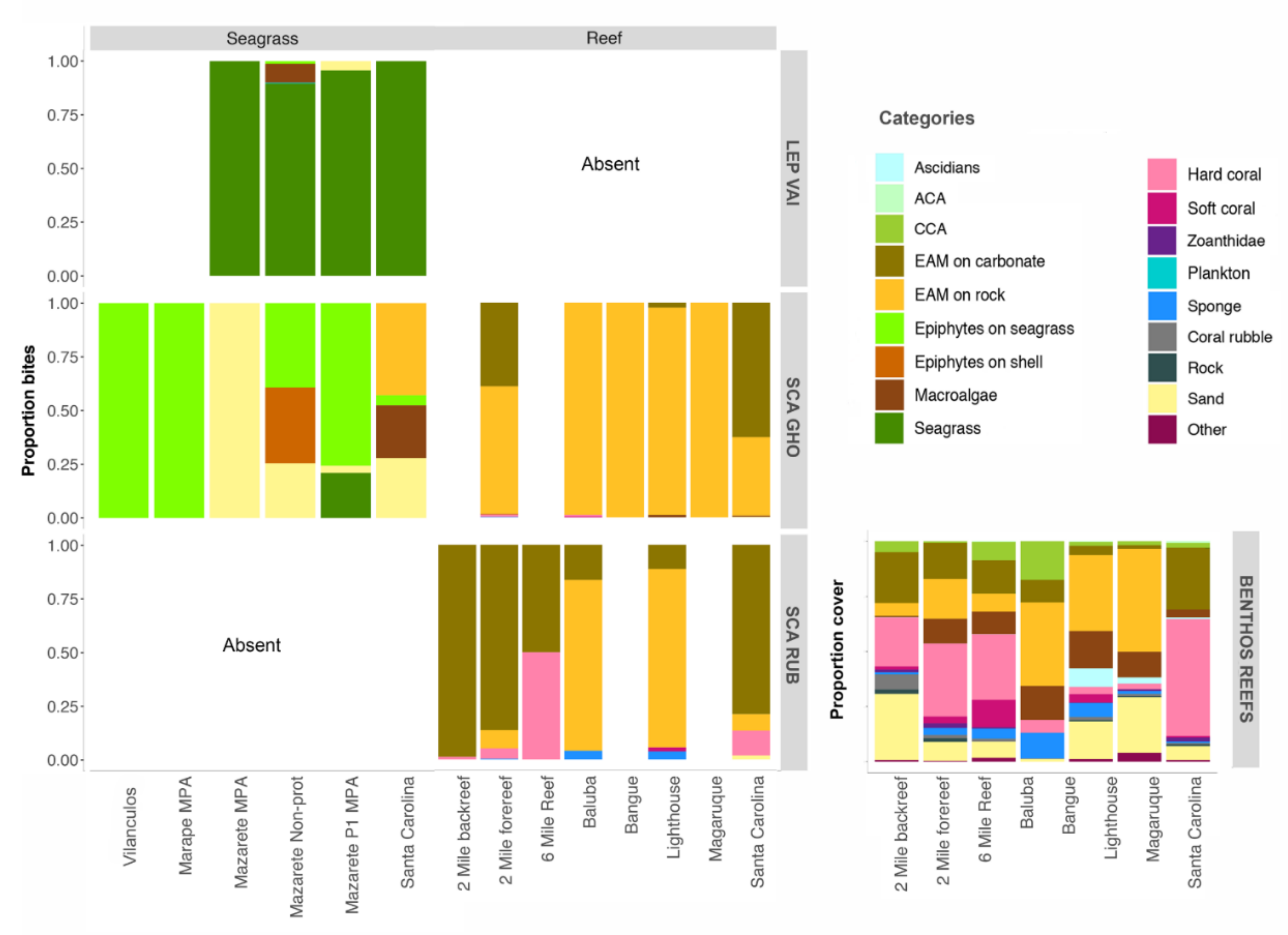
3.2. Habitat Structure
3.2.1. Reefs
3.2.2. Seagrass Meadows
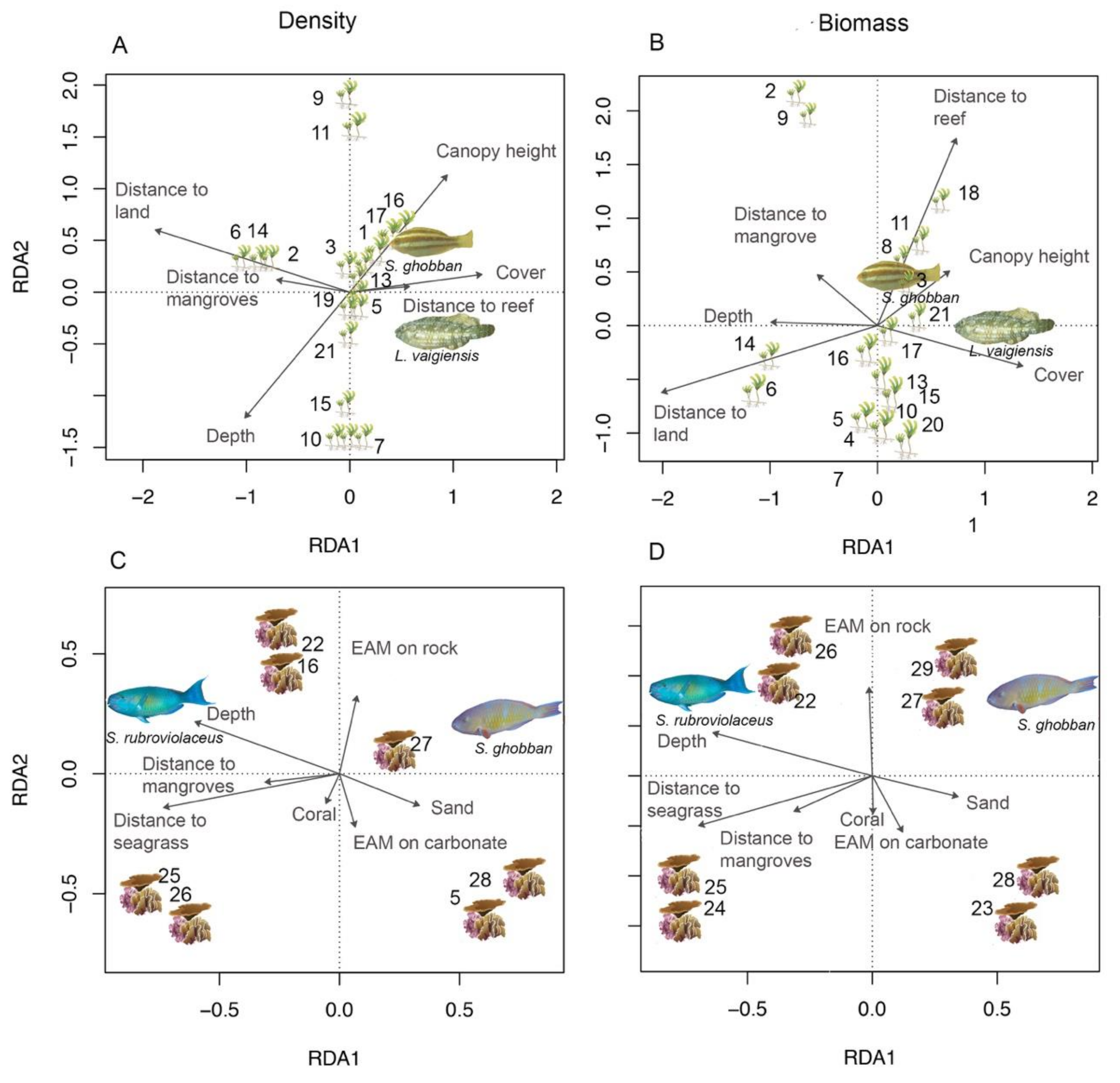
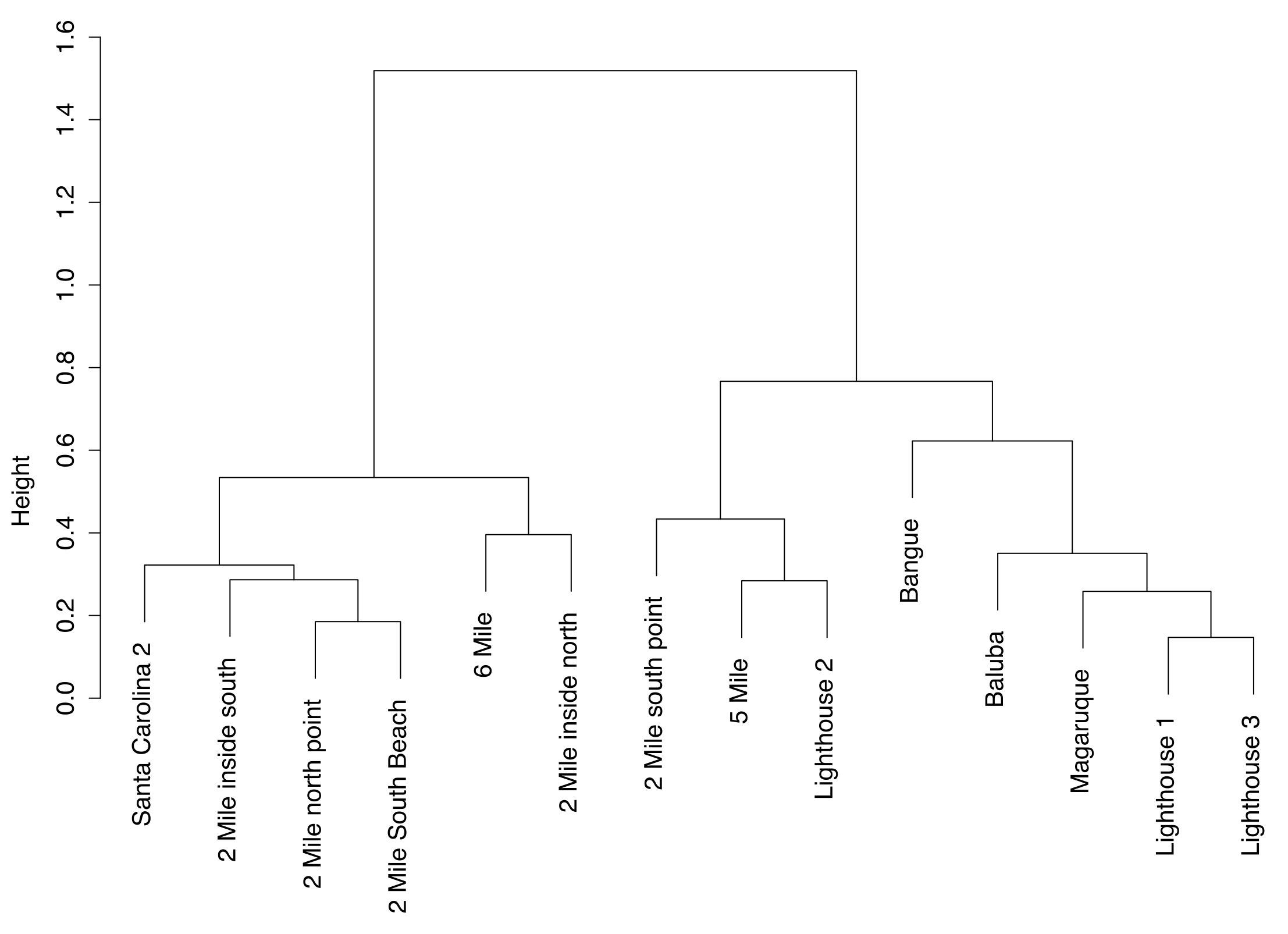
3.3. Spatial Metrics
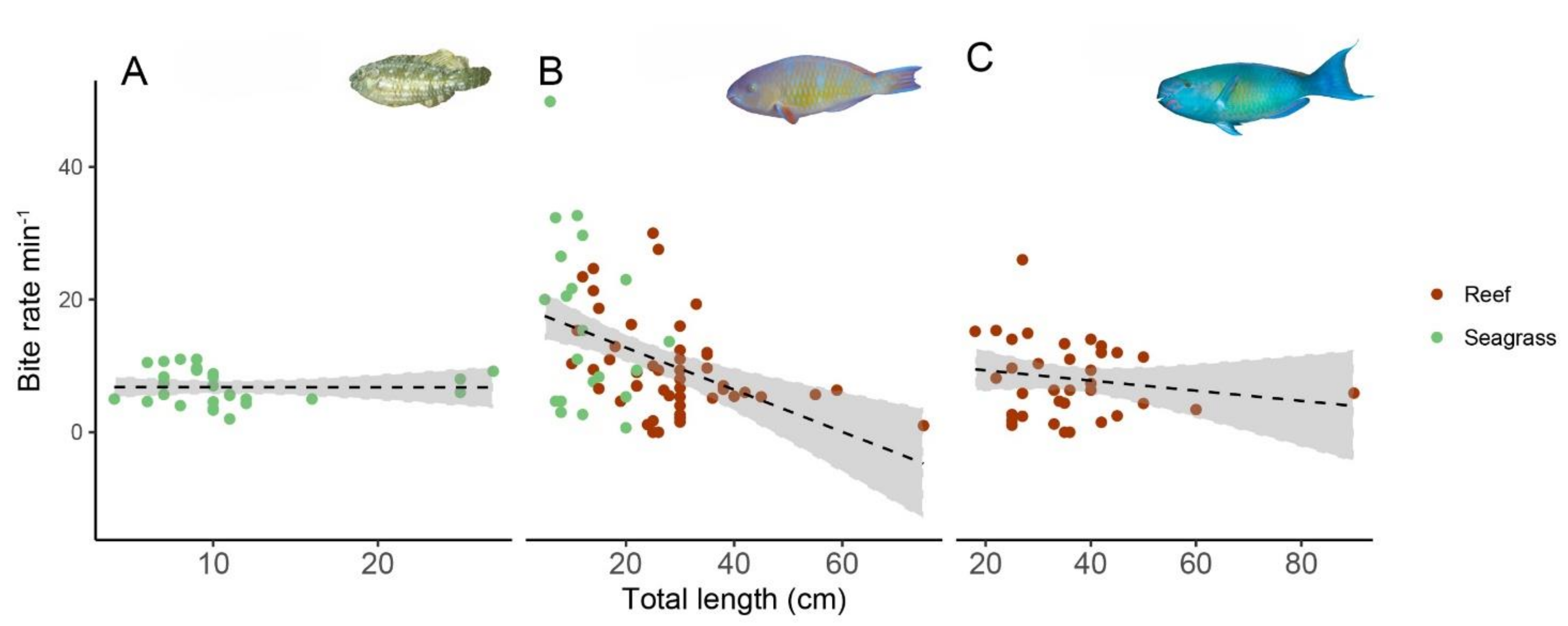
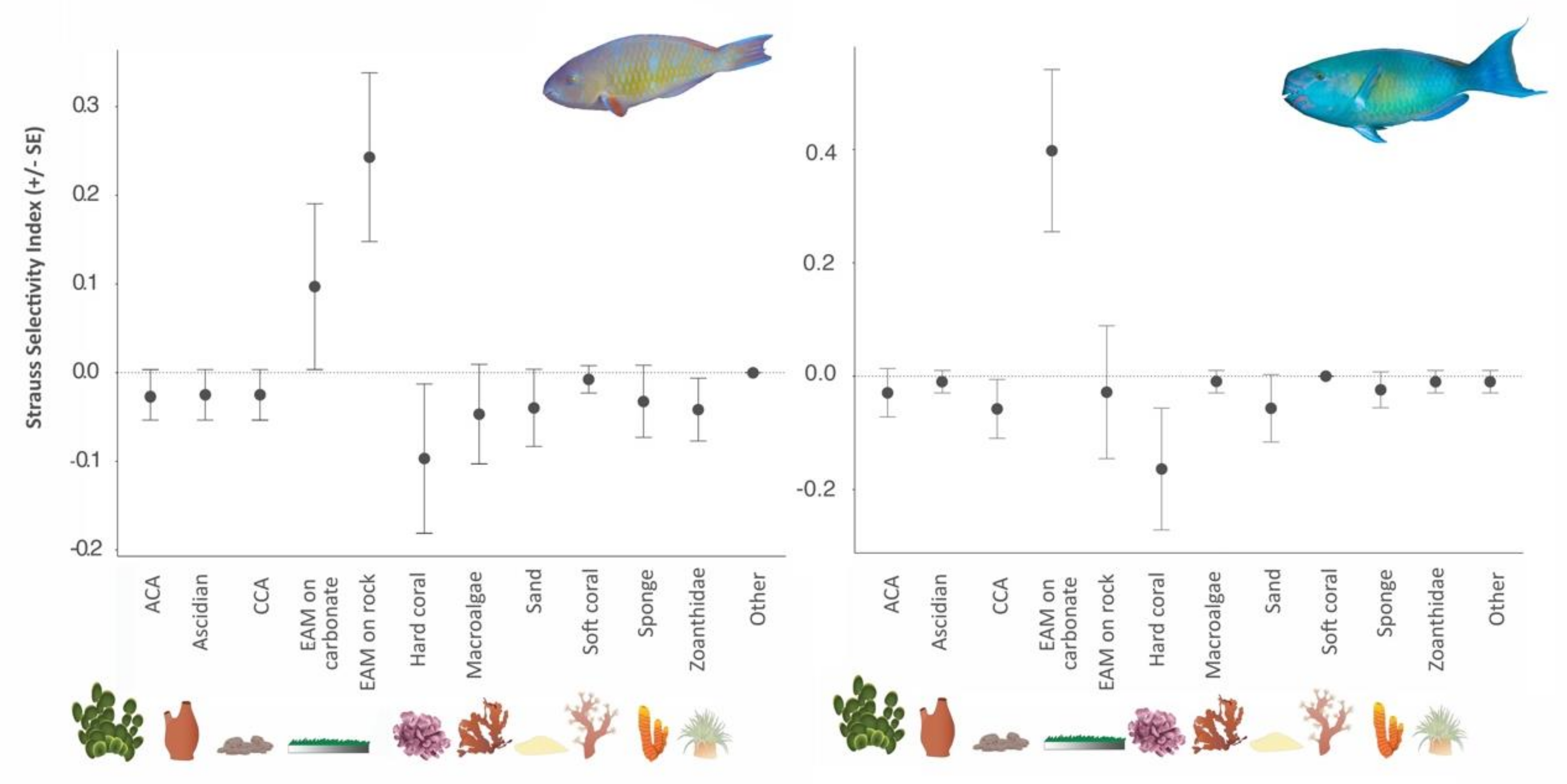
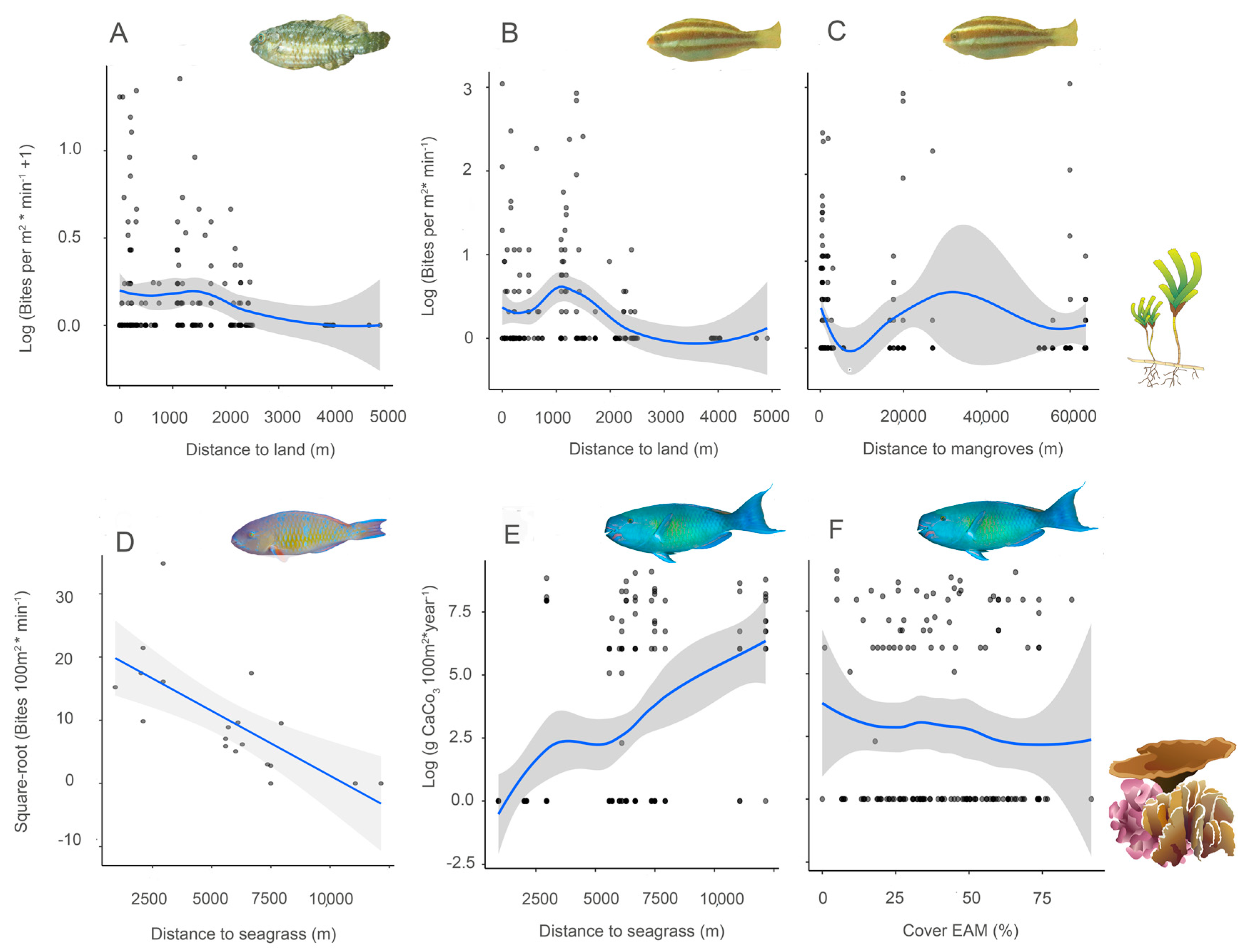
3.4. Bite Rate and Food Selectivity
3.5. Grazing and Bioerosion
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A.—Reef Characteristics


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reef | |||||||
|---|---|---|---|---|---|---|---|
| Site | Average Depth [m] | Coral Cover [%] | EAM Cover [%] | Distance to Seagrass [m] | Distance to Mangroves [m] | ||
| 2 Mile forereef (26) | 9.3 ± 1.2 | 31.5 ± 4.3 | 39.5 ± 3.5 | 6265 | 32,187 | ||
| 2 Mile backreef (26) | 4.8 ± 1.9 | 26.4 ± 6.3 | 29.9 ± 4.2 | 5381 | 32,266 | ||
| 5 Mile (25) | 10.1 ± 0.8 | 21.2 ± 2.9 | 31.2 ± 1.8 | 9999 | 41,755 | ||
| 6 Mile (24) | 8.3 ± 1.5 | 29.7 ± 3.1 | 26.5 ± 4.4 | 11550 | 43,671 | ||
| Baluba (29) | 11.0 ± 1.2 | 6.5 ± 0.6 | 71.0 ± 7.8 | 4546 | 9309 | ||
| Bangue (28) | 1.1 ± 0.1 | 0 ± 0 | 43.7 ± 5.0 | 274 | 6923 | ||
| Lighthouse (22) | 7.7 ± 1.3 | 3.3 ± 0.7 | 41.8 ± 4.7 | 1910 | 64,655 | ||
| Magaruque (27) | 6.3 ± 0.6 | 1.2 ± 0.5 | 58.6 ± 3.7 | 3030 | 15,684 | ||
| St Carolina (23) | 1.9 ± 0.7 | 43.2 ± 5.6 | 43.4 ± 6.6 | 2099 | 54,090 | ||
| Seagrass | |||||||
| Average Depth [m] | Dominant Seagrass Species | Canopy Height [cm] | Seagrass Cover [%] | Distance to land [m] | Distance to Mangrove [m] | Distance to Reef [m] | |
| Aguya (1) | 1.3 ± 0.2 | T. ciliatum | 17.6 ± 4.7 | 35.3 ± 18.0 | 2233 ± 56 | 63,729 ± 51 | 10,909 ± 44 |
| Canal de Deus 1 (6) | 2.4 ± 0.4 | Cymodocea spp | 8.5 ± 0 | 5 ± 0 | 4774 ± 118 | 52,575 ± 17 | 5455 ± 119 |
| Canal de Deus 2 (7) | 2.4 ± 0.1 | Cymodocea spp | 9 ± 0 | 27.5 ± 5 | 4026 ± 8 | 53,825 ± 63 | 4788 ± 9 |
| Chinungwene (20) | 0.9 ± 0.2 | T. hemprichii | 14.8 ± 6.6 | 33.8 ± 17.1 | 177 ± 26 | 1058 ± 88 | 10,175 ± 95 |
| Marape MPA (21) | 1.0 ± 0.1 | T. hemprichii | 19.4 ± 6.6 | 39.1 ± 17.3 | 203 ± 73 | 1026 ± 299 | 13,218 ± 200 |
| Matutuile (4) | 1.8 ± 0.1 | Cymodocea spp and T. hemprichii | 23 ± 6.4 | 80.3 ± 17.4 | 2247 ± 52 | 58,991 ± 50 | 7991 ± 40 |
| Mazarete MPA 2 (17) | 0.9 ± 0.2 | Cymodocea spp and T. hemprichii | 14 ± 3.1 | 37.1 ± 16.5 | 190 ± 36 | 598 ± 62 | 6338 ± 52 |
| Mazarete MPA 1 (18) | 0.9 ± 0.2 | T. hemprichii | 16.8 ± 7.1 | 63.8 ± 20.0 | 41 ± 0 | 172 ± 0 | 6842 ± 0 |
| Mazarete (19) | 1.1 ± 0.5 | Cymodocea spp and T. hemprichii | 11.8 ± 4.4 | 27.4 ± 13.0 | 1139 ± 43 | 484 ± 56 | 7366 ± 213 |
| Mazarete North (15) | 1.0 ± 0.3 | Cymodocea spp and T. hemprichii | 9.2 ± 2.1 | 14.5 ± 5.7 | 1888 ± 199 | 2864 ± 243 | 4997 ± 72 |
| Mukoque (9) | 1.2 ± 0.2 | T. ciliatum | 39.9 ± 8.5 | 52.9 ± 22.9 | 1404 ± 51 | 19,888 ± 41 | 10,218 ± 3 |
| Murungulangene (2) | 1.3 ± 0.2 | T. ciliatum | 11.3 ± 1.2 | 32.9 ± 14.3 | 2365 ± 47 | 63,731 ± 50 | 10,048 ± 0 |
| Palumba (14) | 1.1 ± 0.3 | Halodule uninervis | 12.5 ± 4.4 | 10.6 ± 7.5 | 3913 ± 38 | 5557 ± 30 | 3822 ± 30 |
| Sitone (3) | 1.3 ± 0.4 | Cymodocea spp/T. hemprichii | 17 ± 4.0 | 41.3 ± 24.4 | 16 ± 28 | 59,959 ± 54 | 5521 ± 74 |
| St Carolina (23) | 0.8 ± 0.2 | T. ciliatum | 29.9 ± 5.0 | 78.3 ± 2.6 | 2468 ± 26 | 55791 ± 41 | 2915 ± 28 |
| Vila do Indico (8) | 1.5 ± 0.3 | T. ciliatum | 23.7 ± 7.7 | 74.9 ± 21.3 | 821 ± 470 | 24,322 ± 3650 | 12,240 ± 1647 |
| Vilanculos 1 (10) | 2.3 ± 0.4 | T. ciliatum | 35.7 ± 6.9 | 82.1 ± 4.5 | 195 ± 1 | 18,696 ± 32 | 10,566 ± 10 |
| Vilanculos 3 (12) | 1.0 ± 0.2 | T. ciliatum | 31 ± 14.6 | 88 ± 5.9 | 491 ± 64 | 17,588 ± 39 | 10,290 ± 56 |
| Vilanculos 4 (13) | 0.9 ± 0.1 | T. ciliatum | 41.1 ± 18.8 | 75 ± 16.0 | 690 ± 40 | 16,718 ± 36 | 10,026 ± 43 |
Appendix B.—Fish Data






References
- Lewis, S.M. The Role of Herbivorous Fishes in the Organization of a Caribbean Reef Community. Ecol. Monogr. 1986, 56, 183–200. [Google Scholar] [CrossRef]
- Nyström, M.; Folke, C. Spatial Resilience of Coral Reefs. Ecosystems 2001, 4, 406–417. [Google Scholar] [CrossRef]
- Valentine, J.F.; Duffy, J.E. The Central Role of Grazing in Seagrass Ecology. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 463–501. [Google Scholar]
- Burkepile, D.E.; Hay, M.E. Herbivore Vs. Nutrient Control of Marine Primary Producers: Context-Dependent Effects. Ecology 2006, 87, 3128–3139. [Google Scholar] [CrossRef] [Green Version]
- Burkepile, D.E.; Hay, M.E. Herbivore species richness and feeding complementarity affect community structure and function on a coral reef. Proc. Natl. Acad. Sci. USA 2008, 105, 16201–16206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergés, A.; Vanderklift, M.A.; Doropoulos, C.; Hyndes, G.A. Spatial Patterns in Herbivory on a Coral Reef Are Influenced by Structural Complexity but Not by Algal Traits. PLoS ONE 2011, 6, e17115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, G.P.; Andrew, N.L. Herbivory and patch dynamics on rocky reefs in temperate Australasia: The roles of fish and sea urchins. Austral Ecol. 1990, 15, 505–520. [Google Scholar] [CrossRef]
- McClanahan, T.; Nugues, M.; Mwachireya, S. Fish and sea urchin herbivory and competition in Kenyan coral reef lagoons: The role of reef management. J. Exp. Mar. Biol. Ecol. 1994, 184, 237–254. [Google Scholar] [CrossRef]
- McCook, L.J. Macroalgae, nutrients and phase shifts on coral reefs: Scientific issues and management consequences for the Great Barrier Reef. Coral Reefs 1999, 18, 357–367. [Google Scholar] [CrossRef]
- Swindells, K.L.; Murdoch, R.J.; Bazen, W.D.; Harman, N.W.; Unsworth, R.K. Habitat Configuration Alters Herbivory across the Tropical Seascape. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Rasher, D.B.; Hoey, A.S.; Hay, M.E. Consumer diversity interacts with prey defenses to drive ecosystem function. Ecology 2013, 94, 1347–1358. [Google Scholar] [CrossRef] [Green Version]
- Bonaldo, R.M.; Bellwood, D.R. Spatial variation in the effects of grazing on epilithic algal turfs on the Great Barrier Reef, Australia. Coral Reefs 2010, 30, 381–390. [Google Scholar] [CrossRef]
- Ledlie, M.H.; Graham, N.A.J.; Bythell, J.C.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Hardcastle, J. Phase shifts and the role of herbivory in the resilience of coral reefs. Coral Reefs 2007, 26, 641–653. [Google Scholar] [CrossRef]
- Morgan, K.; Kench, P.S. Parrotfish erosion underpins reef growth, sand talus development and island building in the Maldives. Sediment. Geol. 2016, 341, 50–57. [Google Scholar] [CrossRef]
- Henderson, C.; Olds, A.D.; Lee, S.Y.; Gilby, B.; Maxwell, P.; Connolly, R.M.; Stevens, T. Marine reserves and seascape context shape fish assemblages in seagrass ecosystems. Mar. Ecol. Prog. Ser. 2017, 566, 135–144. [Google Scholar] [CrossRef]
- Dorenbosch, M.; Grol, M.G.G.; Nagelkerken, I.; Van Der Velde, G. Different Surrounding Landscapes may Result in Different Fish Assemblages in East African Seagrass Beds. Hydrobiologia 2006, 563, 45–60. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in browsing intensity on the Great Barrier Reef. Coral Reefs 2010, 29, 499–508. [Google Scholar] [CrossRef]
- Pittman, S.; Caldow, C.; Hile, S.; Monaco, M. Using Seascape Types to Explain the Spatial Patterns of Fish in the Mangroves of SW Puerto Rico. Mar. Ecol. Prog. Ser. 2007, 348, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Streit, R.P.; Cumming, G.S. Patchy delivery of functions undermines functional redundancy in a high diversity system. Funct. Ecol. 2019, 33, 1144–1155. [Google Scholar] [CrossRef]
- Mumby, P.J.; Edwards, A.J.; Arias-Gonzàlez, E.; Lindeman, K.C.; Blackwell, P.G.; Gall, A. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 2004, 427, 533–536. [Google Scholar] [CrossRef] [Green Version]
- Harborne, A.R.; Nagelkerken, I.; Wolff, N.H.; Bozec, Y.-M.; Dorenbosch, M.; Grol, M.G.G.; Mumby, P.J. Direct and indirect effects of nursery habitats on coral-reef fish assemblages, grazing pressure and benthic dynamics. Oikos 2016, 125, 957–967. [Google Scholar] [CrossRef]
- Martin, T.S.H.; Olds, A.D.; Olalde, A.B.H.; Berkström, C.; Gilby, B.L.; Schlacher, T.A.; Butler, I.R.; Yabsley, N.A.; Zann, M.; Connolly, R.M. Habitat proximity exerts opposing effects on key ecological functions. Landsc. Ecol. 2018, 33, 1273–1286. [Google Scholar] [CrossRef] [Green Version]
- Yabsley, N.A.; Olds, A.D.; Connolly, R.M.; Martin, T.S.H.; Gilby, B.L.; Maxwell, P.S.; Huijbers, C.M.; Schoeman, D.S.; Schlacher, T.A. Resource type influences the effects of reserves and connectivity on ecological functions. J. Anim. Ecol. 2015, 85, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Knudby, A.; Jupiter, S.D.; Roelfsema, C.M.; Lyons, M.B.; Phinn, S.R. Mapping Coral Reef Resilience Indicators Using Field and Remotely Sensed Data. Remote. Sens. 2013, 5, 1311–1334. [Google Scholar] [CrossRef] [Green Version]
- Claydon, J.A.B.; Calosso, M.; De Leo, G.A.; Peachey, R. Spatial and demographic consequences of nursery-dependence in reef fishes: An empirical and simulation study. Mar. Ecol. Prog. Ser. 2015, 525, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerken, I.; Dorenbosch, M.; Verbeck, W.C.E.P.; Cocheret de la Moriniere, E.; van der Velde, G. Day-night shifts of fishes between shallow-water biotopes of a Caribbean bay, with emphasis on the nocturnal feeding of Haemulidae and Lutjanidae. Mar Ecol. 2000, 194, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.L.; Schultz, E.T. Migrating haemulid fishes as a source of nutrients and organic matter on coral reefs. Limnol Oceanogr. 1985, 30, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.G.E.; Bennett, E.M.; Gonzalez, A. Linking Landscape Connectivity and Ecosystem Service Provision: Current Knowledge and Research Gaps. Ecosystems. 2013, 16, 894–908. [Google Scholar] [CrossRef]
- Montfrans, J.; Wetzel, R.L.; Orth, R.J. Epiphyte-Grazer Relationships in Seagrass Meadows Consequences for Seagrass Growth and Production I. Estuaries 1984, 7, 289–309. [Google Scholar] [CrossRef]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase Shifts, Herbivory and the Resilience of Coral Reefs to Climate Change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Hauxwell, J.; Cebrián, J.; Valiela, I. Eelgrass Zostera marina loss in temperate estuaries: Relationship to land-derived nitrogen loads and effect of light limitation imposed by algae. Mar. Ecol. Prog. Ser. 2003, 247, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.; Bando, K.; Rodriguez, L.; Williams, S. Relative effects of grazers and nutrients on seagrasses: A meta-analysis approach. Mar. Ecol. Prog. Ser. 2004, 282, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, P.S.; Ekl, J.S.; Katwijk, M.M.; Van Brien, K.R.O. The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems—A review. Biol. Rev. 2016, 92, 1521–1538. [Google Scholar] [CrossRef] [Green Version]
- Thyresson, M.; Nyström, M.; Crona, B. Trading with Resilience: Parrotfish Trade and the Exploitation of Key-Ecosystem Processes in Coral Reefs. Coast. Manag. 2011, 39, 396–411. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Hughes, T.P.; Bellwood, D.R.; Folke, C.; Steneck, R.S.; Wilson, J. New paradigms for supporting the resilience of marine ecosystems. Trends Ecol. Evol. 2005, 20, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R.; Choat, J.H. A functional analysis of grazing in parrotfishes (family Scaridae): The ecological implications. Environ. Biol. Fishes 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Bonaldo, R.; Hoey, A.S.; Bellwood, D.R. The Ecosystem Roles of Parrotfishes on Tropical Reefs. Oceanogr. Mar. Biol. 2014, 52, 81–132. [Google Scholar]
- Wilson, S.K.; Bellwood, D.R.; Choat, J.H.; Furnas, M.J. Detritus in the Epilithic Algal Matrix and Its Use by Coral Reef Fishes. Oceanogr. Mar. Biol. Annu. Rev. 2003, 41, 279–309. [Google Scholar]
- Choat, J.H.; Robbins, W.D.; Clements, K.D. The trophic status of herbivorous fishes on coral reefs. Mar. Biol. 2004, 145, 445–454. [Google Scholar] [CrossRef]
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.; Choat, J.H. Integrating ecological roles and trophic diversification on coral reefs: Multiple lines of evidence identify parrotfishes as microphages. Biol. J. Linn. Soc. 2016, 120, 729–751. [Google Scholar] [CrossRef]
- Hunte, W.; Wittenberg, M. Effects on eutrophication and sedimentation on juvenile corals. Mar Biol. 1992, 631, 625–631. [Google Scholar] [CrossRef]
- Birrell, C.L.; McCook, L.J.; Willis, B.L. Effects of algal turfs and sediment on coral settlement. Mar. Pollut. Bull. 2005, 51, 408–414. [Google Scholar] [CrossRef]
- Mumby, P.J. The Impact of Exploiting Grazers (Scaridae) on the Dynamics of Caribbean Coral Reefs. Ecol. Appl. 2006, 16, 747–769. [Google Scholar] [CrossRef] [Green Version]
- Perry, C.T.; Kench, P.S.; Leary, M.J.O.; Morgan, K.M. Linking reef ecology to island building: Parrotfish identified as major producers of island-building sediment in the Maldives. Geology 2015, 43, 503–506. [Google Scholar] [CrossRef]
- Goatley, C.H.R.; Bellwood, D.R. Sediment suppresses herbivory across a coral reef depth gradient. Biol. Lett. 2012, 8, 1016–1018. [Google Scholar] [CrossRef]
- Goatley, C.H.R.; Bellwood, D.R. Ecological Consequences of Sediment on High-Energy Coral Reefs. PLoS ONE 2013, 8, e77737. [Google Scholar] [CrossRef]
- Airoldi, L. The effects of sedimentation on rocky coast assemblages. Oceanogr. Mar. Biol. 2003, 41, 161–236. [Google Scholar]
- Lobel, P.S.; Ogden, J.C. Foraging by the herbivorous parrotfish Sparisoma radians. Mar. Biol. 1981, 64, 173–183. [Google Scholar] [CrossRef]
- Locham, A.G.; Kaunda-Arara, B.; Wakibia, J.G.; Muya, S. Diet and niche breadth variation in the marbled parrotfish, Leptoscarus vaigiensis, among coral reef sites in Kenya. Afr. J. Ecol. 2015, 53, 560–571. [Google Scholar] [CrossRef]
- Lee, C.-L.; Huang, Y.-H.; Chen, C.-H.; Lin, H.-J. Remote underwater video reveals grazing preferences and drift export in multispecies seagrass beds. J. Exp. Mar. Biol. Ecol. 2016, 476, 1–7. [Google Scholar] [CrossRef]
- Cheal, A.J.; Emslie, M.; MacNeil, M.A.; Miller, I.; Sweatman, H. Spatial variation in the functional characteristics of herbivorous fish communities and the resilience of coral reefs. Ecol. Appl. 2013, 23, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Henderson, C.J.; Gilby, B.L.; Lee, S.Y.; Stevens, T. Contrasting effects of habitat complexity and connectivity on biodiversity in seagrass meadows. Mar. Biol. 2017, 164, 117. [Google Scholar] [CrossRef]
- Pittman, S.J.; McAlpine, C.A.; Pittman, K.M. Linking fish and prawns to their environment: A hierarchical landscape approach. Mar. Ecol. Prog. Ser. 2004, 283, 233–254. [Google Scholar] [CrossRef]
- Bierwagen, S.L.; Heupel, M.R.; Chin, A.; Simpfendorfer, C.A. Trophodynamics as a Tool for Understanding Coral Reef Ecosystems. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Olds, A.D.; Pitt, K.A.; Connolly, R.M.; Maxwell, P.S. Primacy of seascape connectivity effects in structuring reef fish assemblages. Mar. Ecol. Prog. Ser. 2012, 462, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Everett, B.I.; Van der Elst, R.P.; Schleyer, M.H. A Natural History of the Bazaruto Archipelago, 1st ed.; Everett, B.I., Van der Elst, R.P., Schleyer, M.H., Eds.; South African Association for Marine Biological Research: Durban, South Africa, 2008; pp. 1–126. [Google Scholar]
- D’Agata, C. Social and Ecological Factors Influencing Small-Scale Fisheries in the Bay of Bazaruto, Mozambique. Master’s Thesis, Stockholm University, Stockholm, Sweden, 2016. [Google Scholar]
- Díaz, P.; Luís, L.; Mafambissa, M.; Uetimane, A.; Madeira, P.D.; Chambal, E.M. Parque Nacional de Arquipélago de Bazaruto (PNAB). Plano de Maneio 2016–2025 de uma área de Conservação Marinha, Província de Inhambane, Moçambique; Ministério da Terra, Ambiente e Desenvolvimento Rural (MITADER): Maputo, Mozambique, 2016; Volume 1. (In Portuguese) [Google Scholar]
- Taquet, M.; Diringer, A. Fishes of the Indian Ocean and Red Sea, 2nd ed; Editions Quae: Versailles Cedex, France, 2012; p. 224. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication Version (12/2019). 2019. Available online: www.fishbase.org (accessed on 2 November 2020).
- Maggs, J.Q.; Floros, C.; Pereira, M.A.M.; Schleyer, M.H. Short Communication: Rapid Visual Assessment of Fish Communities on Selected Reefs in the Bazaruto Archipelago. West Indian Ocean J. Mar. Sci. 2010, 9, 115–134. [Google Scholar]
- McClanahan, T.R.; Graham, N.A.J.; Maina, J.M.; Chabanet, P.; Bruggemann, J.; Polunin, N.V.C. Influence of instantaneous variation on estimates of coral reef fish populations and communities. Mar. Ecol. Prog. Ser. 2007, 340, 221–234. [Google Scholar] [CrossRef]
- Grandcourt, E.M. Demographic characteristics of a selection of exploited reef fish from the Seychelles: Preliminary study. Mar. Freshw. Res. 2002, 53, 123–130. [Google Scholar] [CrossRef]
- Howard, K.G.; Claisse, J.T.; Clark, T.B.; Boyle, K.S.; Parrish, J.D. Home range and movement patterns of the Redlip Parrotfish (Scarus rubroviolaceus) in Hawaii. Mar. Biol. 2013, 160, 1583–1595. [Google Scholar] [CrossRef]
- Ong, L.; Holland, K.N. Bioerosion of coral reefs by two Hawaiian parrotfishes: Species, size differences and fishery implications. Mar. Biol. 2010, 157, 1313–1323. [Google Scholar] [CrossRef]
- Yarlett, R.T.; Perry, C.T.; Wilson, R.W.; Philpot, K.E. Constraining species—Size class variability in rates of parrotfish bioerosion on Maldivian coral reefs: Implications for regional-scale bioerosion estimates. Mar. Ecol. Prog. Ser. 2018, 590, 155–169. [Google Scholar] [CrossRef]
- Gullström, M.; Castro, M.D.L.T.; Bandeira, S.O.; Björk, M.; Dahlberg, M.; Kautsky, N.; Rönnbäck, P.; Öhman, M.C. Seagrass Ecosystems in the Western Indian Ocean. Ambio 2002, 31, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Eggertsen, L. Identification and Implications of Fish Nurseries in Tropical and Subtropical Seascapes. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 15 March 2019. [Google Scholar]
- Torre-Castro, M.; Eklöf, J.S.; Rönnbäck, P.; Björk, M. Seagrass Importance in Food Provisioning Services: Fish Stomach Content as a Link between Seagrass Meadows and Local Fisheries. West. Indian Ocean J Mar Sci. 2008, 7, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Jiddawi, N.; Öhman, M.C. Marine Fisheries in Tanzania. Ambio 2002, 31, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Cinner, J.E.; McClanahan, T.R.; Graham, N.A.J.; Pratchett, M.S.; Wilson, S.K.; Raina, J.-B. Gear-based fisheries management as a potential adaptive response to climate change and coral mortality. J. Appl. Ecol. 2009, 46, 724–732. [Google Scholar] [CrossRef]
- Gullström, M.; Bodin, M.; Nilsson, P.; Öhman, M. Seagrass structural complexity and landscape configuration as determinants of tropical fish assemblage composition. Mar. Ecol. Prog. Ser. 2008, 363, 241–255. [Google Scholar] [CrossRef] [Green Version]
- Berkström, C.; Eggertsen, L.; Goodell, W.; Cordeiro, C.A.M.M.; Lucena, M.B.; Gustafsson, R.; Bandeira, S.; Jiddawi, N.; Ferreira, C.L. Thresholds in seascape connectivity: The spatial arrangement of nursery habitats structure fish communities on nearby reefs. Ecography 2020, 43, 882–896. [Google Scholar] [CrossRef]
- Bonaldo, R.M.; Bellwood, D.R. Dynamics of parrotfish grazing scars. Mar. Biol. 2009, 156, 771–777. [Google Scholar] [CrossRef]
- Lokrantz, J.; Nystrom, M.; Thyresson, M.; Johansson, C. The non-linear relationship between body size and function in parrotfishes. Coral Reefs 2008, 27, 967–974. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; McQuaid, C.D.; Hill, J.M. The effects of tissue type and body size onδ13C andδ15N values in parrotfish (Labridae) from Zanzibar, Tanzania. J. Appl. Ichthyol. 2015, 31, 633–637. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; McQuaid, C.D.; Hill, J.M. Stable isotope analysis indicates a lack of inter- and intra-specific dietary redundancy among ecologically important coral reef fishes. Coral Reefs 2012, 32, 429–440. [Google Scholar] [CrossRef]
- Brock, R.E. A critique of the visual census method for assessing coral reef fish populations. Bull. Mar. Sci. 1982, 32, 269–276. [Google Scholar]
- Trygonis, V.; Sini, M. photoQuad: A dedicated seabed image processing software and a comparative error analysis of four photoquadrat methods. J. Exp. Mar. Biol. Ecol. 2012, 424, 99–108. [Google Scholar] [CrossRef]
- Boström, C.; Jackson, E.L.; Simenstad, C.A. Seagrass landscapes and their effects on associated fauna: A review. Estuar. Coast. Shelf Sci. 2006, 68, 383–403. [Google Scholar] [CrossRef]
- Morgan, K.; Kench, P. Skeletal extension and calcification of reef-building corals in the central Indian Ocean. Mar. Environ. Res. 2012, 81, 78–82. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Team R DC. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Strauss, R.E. Reliability Estimates for Ivlev’s Electivity Index, the Forage Ratio and a Proposed Linear Index of Food Selection. Trans. Am. Fish. Soc. 1979, 108, 344–352. [Google Scholar] [CrossRef]
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: Fort Collins, CO, USA, 2002; p. 485. [Google Scholar]
- Wood, S.N.; Pya, N.; Säfken, B.; Wood, S.N.; Pya, N.; Säfken, B. Smoothing Parameter and Model Selection for General Smooth Models. J. Am. Stat. Assoc. 2016, 111, 1548–1575. [Google Scholar] [CrossRef]
- Ecological Connectivity among Tropical Coastal Ecosystems; Nagelkerken, I. (Ed.) Springer: Heidelberg, Germany, 2009. [Google Scholar]
- Mumby, P. Connectivity of reef fish between mangroves and coral reefs: Algorithms for the design of marine reserves at seascape scales. Biol. Conserv. 2006, 128, 215–222. [Google Scholar] [CrossRef]
- Dorenbosch, M.; Grol, M.; Christianen, M.; Nagelkerken, I.; Van Der Velde, G. Indo-Pacific seagrass beds and mangroves contribute to fish density and diversity on adjacent coral reefs. Mar. Ecol. Prog. Ser. 2005, 302, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Lugendo, B.R.; Pronker, A.; Cornelissen, I.; De Groene, A.; Nagelkerken, I.; Dorenbosch, M.; Van Der Velde, G.; Mgaya, Y.D. Habitat utilisation by juveniles of commercially important fish species in a marine embayment in Zanzibar, Tanzania. Aquat. Living Resour. 2005, 18, 149–158. [Google Scholar] [CrossRef]
- Mellin, C.; Andréfouët, S.; Ponton, D. Spatial predictability of juvenile fish species richness and abundance in a coral reef environment. Coral Reefs 2007, 26, 895–907. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving resource partitioning in parrotfishes (Scarini) using microhistology of feeding substrata. Coral Reefs 2020, 1–15. [Google Scholar] [CrossRef]
- Goatley, C.H.; Bellwood, D.R. Biologically mediated sediment fluxes on coral reefs: Sediment removal and off-reef transportation by the surgeonfish Ctenochaetus striatus. Mar. Ecol. Prog. Ser. 2010, 415, 237–245. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Fulton, C.J. Sediment-mediated suppression of herbivory on coral reefs: Decreasing resilience to rising sea-levels and climate change? Limnol. Oceanogr. 2008, 53, 2695–2701. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Bellwood, D.R. Fine sediments suppress detritivory on coral reefs. Mar. Pollut. Bull. 2017, 114, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Crossman, D.J.; Choat, H.J.; Clements, K.D.; Hardy, T.; McConochie, J. Detritus as food for grazing fishes on coral reefs. Limnol. Oceanogr. 2001, 46, 1596–1605. [Google Scholar] [CrossRef]
- Purcell, S.; Bellwood, D. Spatial patterns of epilithic algal and detrital resources on a windward coral reef. Coral Reefs 2001, 20, 117–125. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2007, 27, 37–47. [Google Scholar] [CrossRef]
- Carlson, P.M.; Davis, K.; Warner, R.R.; Caselle, J.E. Fine-scale spatial patterns of parrotfish herbivory are shaped by resource availability. Mar Ecol Prog Ser. 2017, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Berkström, C.; Gullström, M.; Lindborg, R.; Mwandya, A.W.; Yahya, S.A.; Kautsky, N.; Nyström, M. Exploring ‘knowns’ and ‘unknowns’ in tropical seascape connectivity with insights from East African coral reefs. Estuar. Coast. Shelf Sci. 2012, 107, 1–21. [Google Scholar] [CrossRef]
- Lundberg, J.; Moberg, F. Mobile Link Organisms and Ecosystem Functioning: Implications for Ecosystem Resilience and Management. Ecosystems 2003, 6, 0087–0098. [Google Scholar] [CrossRef]





| Seagrass Site | L. vaigiensis | S. ghobban | ||
|---|---|---|---|---|
| Bites m−2 min−1 | Biomass g m−2 | Bites m−2 min−1 | Biomass g m−2 | |
| Aguya (1) | 0.034 | 0.000 | 0.282 | 0.000 |
| Chinungwene (20) | 0.247 | 0.188 ± 0.000 | 0.000 | 0.000 |
| Canal de Deus 1 (6) | 0.000 | 0.000 | 0.000 | 0.000 |
| Canal de Deus 2 (7) | 0.000 | 0.000 | 0.000 | 0.000 |
| Marape MPA (21) | 0.321 | 1.319 ± 0.098 | 0.423 | 2.400 ± 0.043 |
| Matutuile (4) | 0.229 | 2.302 ± 0.378 | 0.000 | 0.000 |
| Mazarete (19) | 0.245 | 0.116 ± 0.027 | 1.716 | 0.062 ± 0.015 |
| Mazarete MPA 1 (18) | 0.011 | 0.050 ± 0.027 | 0.125 | 0.026 ± 0.009 |
| Mazarete MPA 2 (17) | 0.692 | 0.304 ± 0.051 | 2.351 | 0.140 ± 0.024 |
| Mazarete North (15) | 0.225 | 0.265 ± 0.050 | 0.125 | 0.008 ± 0.007 |
| Mazarete MPA 3 (16) | 0.693 | 1.485 ± 0.214 | 2.473 | 0.580 ± 0.151 |
| Mukoque (9) | 0.015 | 0.020 ± 0.020 | 4.435 | 1.290 ± 0.185 |
| Murungulangene (2) | 0.000 | 0.000 | 0.000 | 0.021 ± 0.013 |
| Palumba (14) | 0.000 | 0.000 | 0.000 | 0.000 |
| Santa Carolina (23) | 0.047 | 1.124 ± 1.124 | 0.063 | 0.071 ± 0.071 |
| Sitone (3) | 0.810 | 0.039 ± 0.012 | 3.762 | 0.088 ± 0.010 |
| Vila do Indico (8) | 0.068 | 0.403 ± 0.256 | 0.893 | 0.494 ± 0.145 |
| Vilanculos 2 (11) | 0.022 | 0.059 ± 0.059 | 0.690 | 0.120 ± 0.041 |
| Vilanculos 3 (12) | 0.090 | 0.281 ± 0.106 | 0.000 | 0.000 |
| Vilanculos 4 (13) | 0.067 | 0.110 ± 0.056 | 0.125 | 0.045 ± 0.036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eggertsen, L.; Goodell, W.; Cordeiro, C.A.M.M.; Mendes, T.C.; Longo, G.O.; Ferreira, C.E.L.; Berkström, C. Seascape Configuration Leads to Spatially Uneven Delivery of Parrotfish Herbivory across a Western Indian Ocean Seascape. Diversity 2020, 12, 434. https://doi.org/10.3390/d12110434
Eggertsen L, Goodell W, Cordeiro CAMM, Mendes TC, Longo GO, Ferreira CEL, Berkström C. Seascape Configuration Leads to Spatially Uneven Delivery of Parrotfish Herbivory across a Western Indian Ocean Seascape. Diversity. 2020; 12(11):434. https://doi.org/10.3390/d12110434
Chicago/Turabian StyleEggertsen, Linda, Whitney Goodell, César A. M. M. Cordeiro, Thiago C. Mendes, Guilherme O. Longo, Carlos E. L. Ferreira, and Charlotte Berkström. 2020. "Seascape Configuration Leads to Spatially Uneven Delivery of Parrotfish Herbivory across a Western Indian Ocean Seascape" Diversity 12, no. 11: 434. https://doi.org/10.3390/d12110434