Interactions among Shade, Caching Behavior, and Predation Risk May Drive Seed Trait Evolution in Scatter-Hoarded Plants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Seed Dispersal by Scatter-Hoarding Granivores
1.2. Scatter-Hoarding as a Behavioral and Evolutionary Strategy
2. Materials and Methods
2.1. Study System
2.2. Model Description
2.3. Analysis
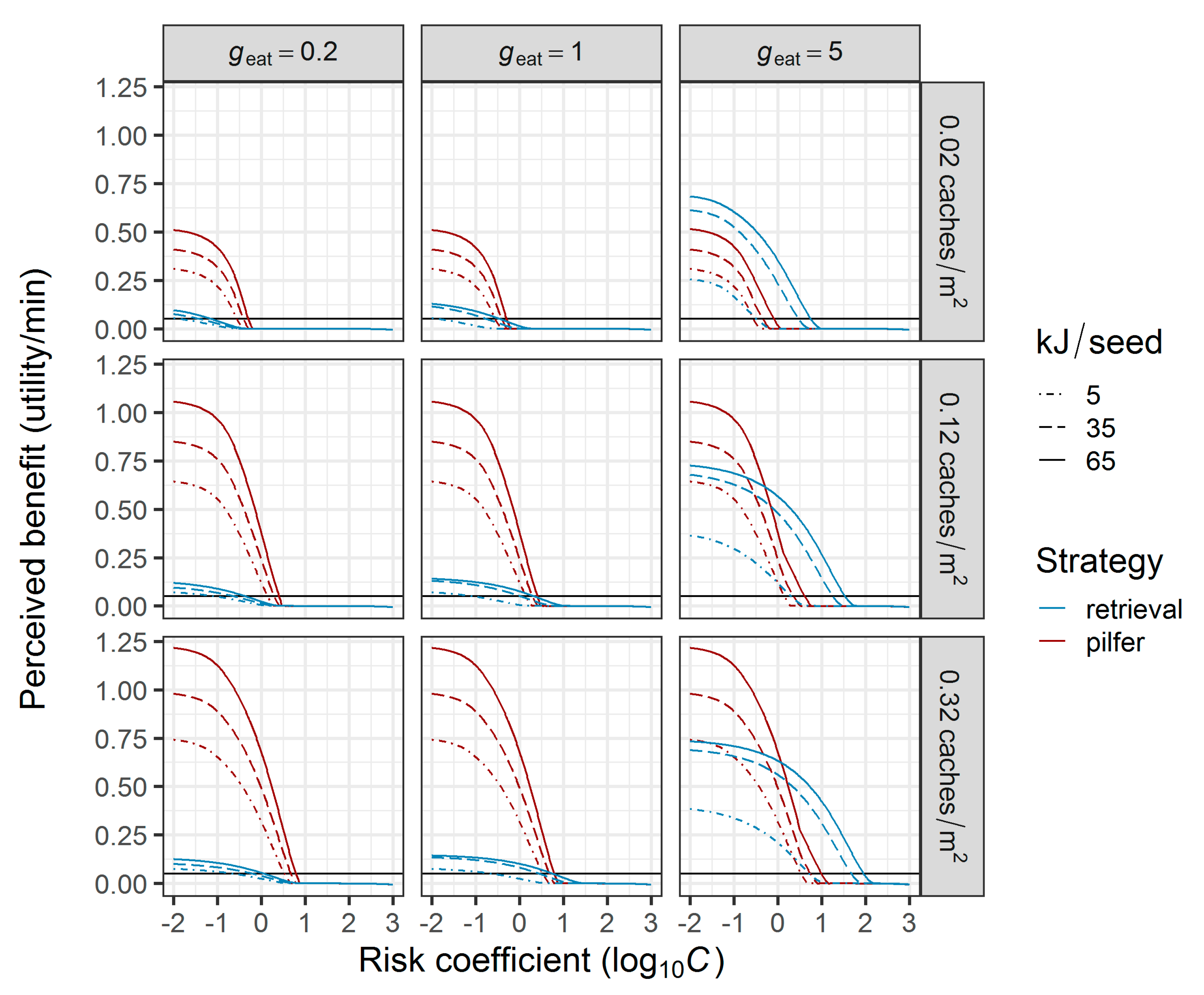
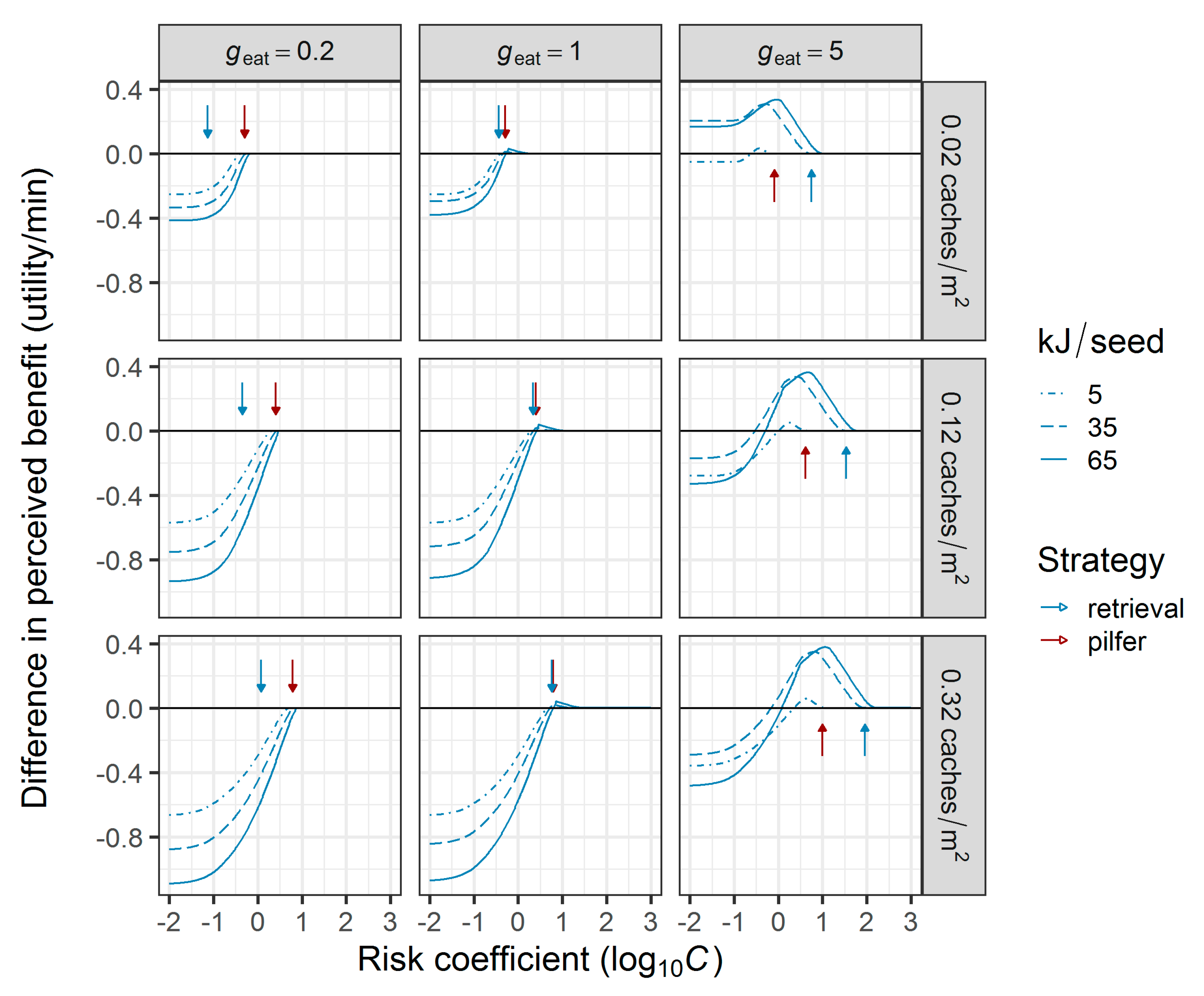
2.3.1. Comparison of Foraging Strategies under Predation Risk
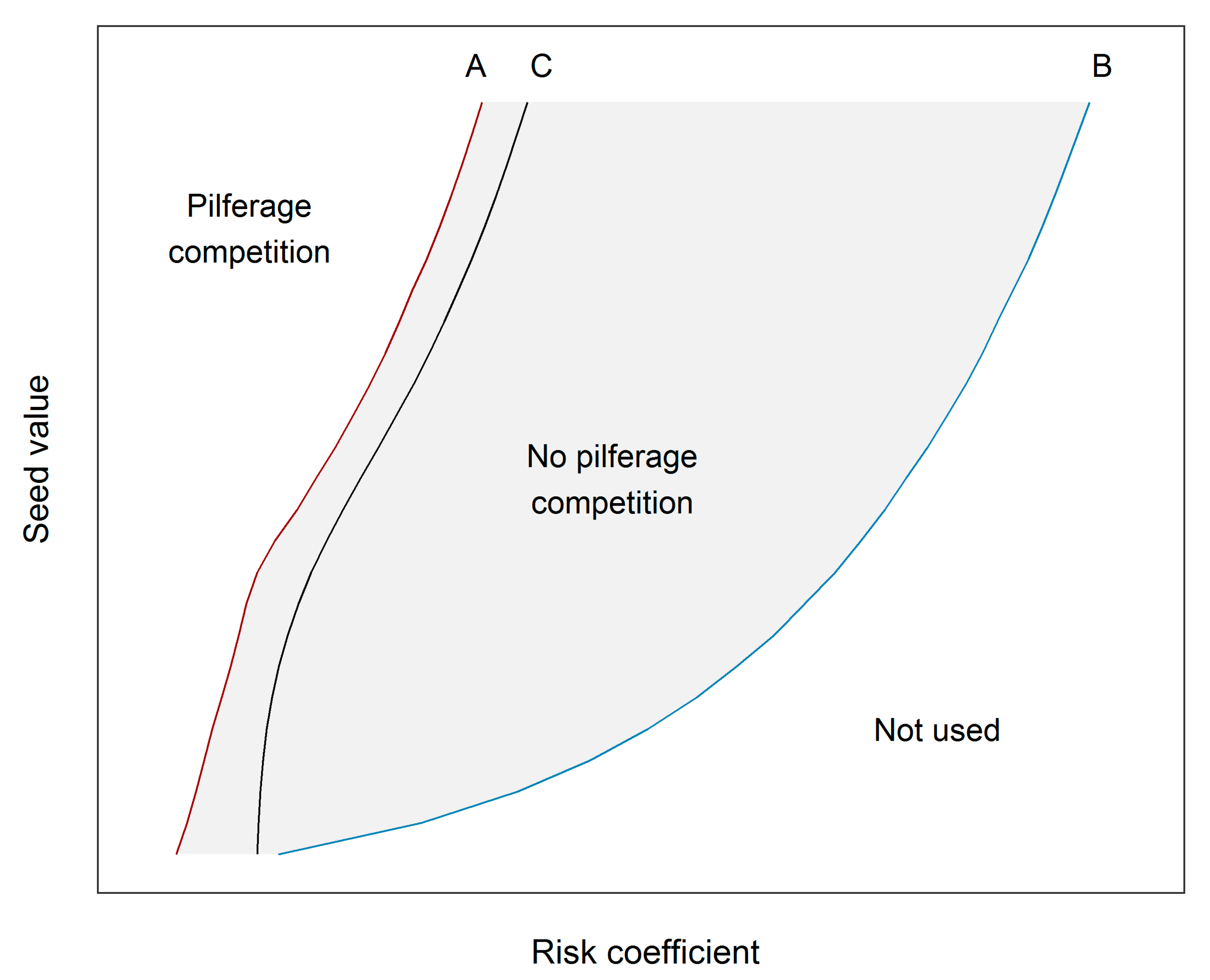
2.3.2. Effects of Seed Value on the Risk Level Needed to Protect a Cache
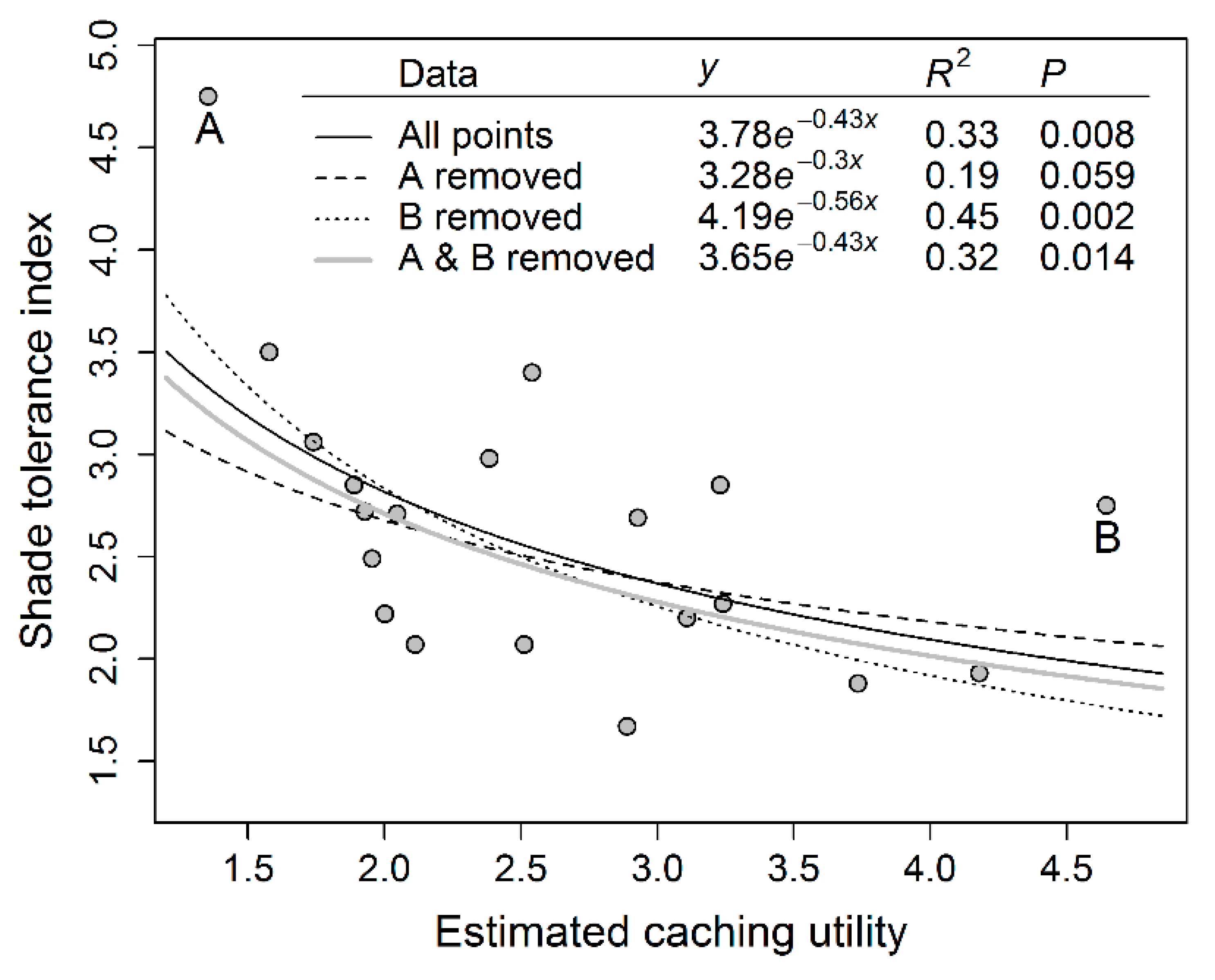
2.3.3. Seed Traits and Shade Tolerance
3. Results
3.1. Comparison of Foraging Strategies under Predation Risk
3.2. Effects of Seed Value on the Risk Level Needed to Protect a Cache
3.3. Seed Traits and Shade Tolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schupp, E.W.; Jordano, P.; Gómez, J.M. Seed dispersal effectiveness revisited: A conceptual review. New Phytol. 2010, 188, 333–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConkey, K.R.; Prasad, S.; Corlett, R.T.; Campos-Arceiz, A.; Brodie, J.F.; Rogers, H.; Santamaria, L. Seed dispersal in changing landscapes. Biol. Conserv. 2012, 146, 1–13. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Schupp, E.W. Seed seedling conflicts, habitat choice, and patterns of plant recruitment. Am. J. Bot. 1995, 82, 399–409. [Google Scholar] [CrossRef]
- Schupp, E.W.; Fuentes, M. Spatial patterns of seed dispersal and unification of plant population ecology. Ecoscience 1995, 2, 267–275. [Google Scholar] [CrossRef]
- Crawley, M.; Long, C. Alternate bearing, predator satiation and seedling recruitment in Quercus robur L. J. Ecol. 1995, 83, 683–696. [Google Scholar] [CrossRef]
- Hulme, P.E.; Kollmann, J. Seed Predator Guilds, Spatial Variation in Post-Dispersal Seed Predation and Potential Effects on Plant Demography: A Temperate Perspective; Forget, P.M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CAB International: Wallingford, UK, 2005; pp. 9–30. [Google Scholar]
- Darwin, C. On the action of sea-water on the germination of seeds. J. Proc. Linn. Soc. Lond. 1857, 1, 130–140. [Google Scholar]
- Vander Wall, S.B. How plants manipulate the scatter-hoarding behaviour of seed-dispersing animals. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 989–997. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Z.; Chang, G.; Yi, X.; Lu, J.; Xiao, Z.; Zhang, H.; Cao, L.; Want, F.; Li, H.; et al. Trade-off between seed defensive traits and impacts on interaction patterns between seeds and rodents in forest ecosystems. Plant Ecol. 2016, 217, 253–265. [Google Scholar] [CrossRef]
- Levin, S.A.; Müller-Landau, H.C.; Nathan, R.; Chave, J. The ecology and evolution of seed dispersal: A theoretical perspective. Ann. Rev. Ecol. Evol. Syst. 2003, 34, 575–604. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, A.; Guisan, M.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Cousens, R.D.; Hill, J.; French, K.; Bishop, I.D. Towards better prediction of seed dispersal by animals. Funct. Ecol. 2010, 24, 1163–1170. [Google Scholar] [CrossRef]
- Lichti, N.I.; Steele, M.A.; Swihart, R.K. Seed fate and decision-making processes in scatter- hoarding rodents. Biol. Rev. 2017, 92, 474–504. [Google Scholar] [CrossRef]
- Vander Wall, S.B. Food Hoarding in Animals; University of Chicago Press: Chicago, IL, USA, 1990; pp. 1–44. [Google Scholar]
- Morris, D. The behaviour of the green acouchi (Myoprocta pratti) with special reference to scatter hoarding. Proc. Zool. Soc. Lond. 1962, 139, 701–732. [Google Scholar] [CrossRef]
- Steele, M.A.; Smallwood, P.D. Acorn Dispersal by Birds and Mammals. In Oak Forest Ecosystems: Ecology and Management for Wildlife; McShea, W., Healy, W., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2002; pp. 182–195. [Google Scholar]
- Price, M.V.; Jenkins, S.H. Rodents as seed consumers and dispersers. In Seed Dispersal; Murray, D.R., Ed.; Academic Press: Sydney, Australia, 1986; pp. 191–235. [Google Scholar]
- Vander Wall, S.B. The evolutionary ecology of nut dispersal. Bot. Rev. 2001, 67, 74–117. [Google Scholar] [CrossRef]
- Shimada, T. Hoarding behaviors of two wood mouse species: Different preference for acorns of two Fagaceae species. Ecol. Res. 2001, 16, 127–133. [Google Scholar] [CrossRef]
- Klinger, R.; Rejmánek, M. The numerical and functional responses of a granivorous rodent and the fate of Neotropical tree seeds. Ecology 2009, 90, 1549–1563. [Google Scholar] [CrossRef]
- Lichti, N.I.; Steele, M.A.; Zhang, H.; Swihart, R.K. Mast species composition alters seed fate in North American rodent-dispersed hardwoods. Ecology 2014, 95, 1746–1758. [Google Scholar] [CrossRef]
- Sivy, K.J.; Ostoja, S.M.; Schupp, E.W.; Durham, S. Effects of rodent species, seed species, and predator cues on seed fate. Acta Oecol. 2011, 37, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Jansen, P.A.; Bongers, F.; Hemerik, L. Seed mass and mast seeding enhance dispersal by a neotropical scatter-hoarding rodent. Ecol. Monogr. 2004, 74, 569–589. [Google Scholar] [CrossRef]
- Galvez, D.; Kranstauber, B.; Kays, R.W.; Jansen, P.A. Scatter hoarding by the Central American agouti: A test of optimal cache spacing theory. Anim. Behav. 2009, 78, 1327–1333. [Google Scholar] [CrossRef]
- Perea, R.; San Miguel, A.; Martínez-Jauregui, M.; Valbuena-Carabaña, M.; Gil, L. Effects of seed quality and seed location on the removal of acorns and beechnuts. Eur. J. For. Res. 2012, 131, 623–631. [Google Scholar] [CrossRef]
- Wang, B.; Chen, J. Effects of fat and protein levels on foraging preferences of tannin in scatter- hoarding rodents. PLoS ONE 2012, 7, e40640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Zhang, Z.; Wang, Y. Dispersal and germination of big and small nuts of Quercus serrata in a subtropical broad-leaved evergreen forest. For. Ecol. Manag. 2004, 195, 141–150. [Google Scholar] [CrossRef]
- Cheng, J.; Xiao, Z.; Zhang, Z. Seed consumption and caching on seeds of three sympatric tree species by four sympatric rodent species in a subtropical forest, China. For. Ecol. Manag. 2005, 216, 331–341. [Google Scholar] [CrossRef]
- Vander Wall, S.B. Effects of seed size of wind dispersed pines (Pinus) on secondary seed dispersal and the caching behavior of rodents. Oikos 2003, 100, 25–34. [Google Scholar] [CrossRef]
- Yi, X.; Yang, Y. Scatterhoarding of Manchurian walnut (Juglans mandshurica) by small mammals: Response to seed familiarity and seed size. Acta Theriol. 2011, 56, 141–147. [Google Scholar] [CrossRef]
- Yi, X.; Yang, Y.; Zhang, Z. Intra-and inter-specific effects of mast seeding on seed fates of two sympatric Corylus species. Plant Ecol. 2011, 212, 785–793. [Google Scholar] [CrossRef]
- Hadj-Chikh, L.Z.; Steele, M.A.; Smallwood, P.D. Caching decisions by grey squirrels: A test of the handling time and perishability hypotheses. Anim. Behav. 1996, 52, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.A.; Wauters, W.A.; Larsen, K.W. Selection, predation and dispersal of seeds by tree squirrels in temperate and boreal forests: Are tree squirrels keystone granivores? In Seed Fate: Predation, Dispersal, and Seedling Establishment; Forget, P.-M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CAB International: Cambridge, MA, USA, 2005; pp. 205–221. [Google Scholar]
- Moore, J.E.; McEuen, A.B.; Swihart, R.K.; Contreras, T.A.; Steele, M.A. Determinants of seed removal distance by scatter-hoarding rodents in deciduous forests. Ecology 2007, 88, 2529–2540. [Google Scholar] [CrossRef]
- Steele, M.A. Evolutionary Interactions between Tree Squirrels and Trees: A Review and Synthesis. Curr. Sci. 2008, 10, 871–876. [Google Scholar]
- Xiao, Z.; Gao, X.; Jiang, M.; Zhang, Z. Behavioral adaptation of Pallas’s squirrels to germination schedule and tannins in acorns. Behav. Ecol. 2009, 20, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Gao, X.; Zhang, Z. The combined effects of seed perishability and seed size on hoarding decisions by Pére David’s rock squirrels. Behav. Ecol. Sociobiol. 2013, 67, 1067–1075. [Google Scholar] [CrossRef]
- Wang, B.; Corlett, R.T. Scatter-hoarding rodents select different caching habitats for seeds with different traits. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- Wang, B.; Ye, C.-X.; Cannon, C.H.; Chen, J. Dissecting the decision making process of scatter- hoarding rodents. Oikos 2013, 122, 1027–1034. [Google Scholar] [CrossRef]
- Sundaram, M.; Lichti, N.I.; Steele, M.A.; Dalgleish, H.J.; Swihart, R.K. Frequency-dependent hoarding by Sciurus carolinensis occurs with seeds of similar perceived value. J. Mammal. 2017, 98, 124–134. [Google Scholar]
- Andersson, M.; Krebs, J.R. On the evolution of hoarding behavior. Anim. Behav. 1978, 26, 707–711. [Google Scholar] [CrossRef]
- Vander Wall, S.B.; Jenkins, S.H. Reciprocal pilferage and the evolution of food-hoarding behavior. Behav. Ecol. 2003, 14, 656–667. [Google Scholar] [CrossRef] [Green Version]
- Kamil, A.C.; Gould, K.L. Memory in Food Caching Animals. In Learning and Memory: A Comprehensive Reference. In Learning Theory and Behaviour; Menzel, R., Byrne, J.H., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2008; pp. 419–439. [Google Scholar]
- Brodin, A. The history of scatter-hoarding studies. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 869–881. [Google Scholar] [CrossRef]
- Hirsch, B.; Kays, R.; Jansen, P. Evidence for cache surveillance by a scatter-hoarding rodent. Anim. Behav. 2013, 85, 1511–1516. [Google Scholar] [CrossRef]
- Bartlow, A.W.; Lichti, N.; Curtis, R.; Swihart, R.; Steele, M.A. Re-caching of acorns by rodents: Cache management in eastern deciduous forests of North America. Acta Oecol. 2018, 92, 117–122. [Google Scholar] [CrossRef]
- Jacobs, L.F. Memory for cache locations in Merriam’s kangaroo rats. Anim. Behav. 1992, 43, 585–593. [Google Scholar] [CrossRef]
- Jacobs, L.F.; Liman, E.R. Grey squirrels remember the locations of buried nuts. Anim. Behav. 1991, 41, 103–110. [Google Scholar] [CrossRef]
- Steele, M.A.; Bugdal, M.; Yuan, A.; Bartlow, A.; Buzalewski, J.; Lichti, N.; Swihart, R. Cache placement, pilfering, and a recovery advantage in a seed-dispersing rodent: Could predation of scatter hoarders contribute to seedling establishment? Acta Oecol. 2011, 37, 554–560. [Google Scholar] [CrossRef]
- Jansen, P.A.; Hirsch, B.T.; Emsens, W.J.; Zamora-Gutierrez, V.; Wikelski, M.; Kays, R. Thieving rodents as substitute dispersers of megafaunal seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 12610–12615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapanian, M.A.; Smith, C.C. A model for seed scatterhoarding: Coevolution of fox squirrels and black walnuts. Ecology 1978, 59, 884–896. [Google Scholar] [CrossRef]
- Stapanian, M.A.; Smith, C.C. Density-dependent survival of scatterhoarded nuts: An experimental approach. Ecology 1984, 65, 1387–1396. [Google Scholar] [CrossRef]
- Hirsch, B.T.; Kays, R.; Pereira, V.E.; Jansen, P.A. Directed seed dispersal towards areas with low conspecific tree density by a scatter-hoarding rodent. Ecol. Lett. 2012, 15, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.S.; Vander Wall, S.B. Substrate type affects caching and pilferage of pine seeds by chipmunks. Behav. Ecol. 2004, 15, 666–672. [Google Scholar] [CrossRef]
- Brown, J.S. Patch use as an indicator of habitat preference, predation risk, and competition. Behav. Ecol. Sociobiol. 1988, 22, 37–47. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P. Hazardous duty pay and the foraging cost of predation. Ecol. Lett. 2004, 7, 999–1014. [Google Scholar] [CrossRef]
- Preston, S.D.; Jacobs, L.F. Cache decision making: The effects of competition on cache decisions in Merriam’s kangaroo rat (Dipodomys merriami). J. Comp. Psychol. 2005, 119, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Bonal, R. Linking seed dispersal to cache protection strategies. J. Ecol. 2011, 99, 1016–1025. [Google Scholar] [CrossRef]
- Steele, M.A.; Contreras, T.A.; Hadj-Chikh, L.Z.; Agosta, S.J.; Smallwood, P.D.; Tomlinson, C.N. Do scatter hoarders trade off increased predation risks for lower rates of cache pilferage? Behav. Ecol. 2014, 25, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.; Rompré, G.; Stratford, J.; Zhang, H.; Suchocki, M.; Marino, S. Scatter-hoarding rodents favor higher predation risks for cache sites: The potential for predators to influence the seed dispersal process. Integr. Zool. 2015, 10, 257–266. [Google Scholar] [CrossRef]
- Holling, C.S. The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Can. Ent. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Brown, J.S. Vigilance, patch use and habitat selection: Foraging under predation risk. Evol. Ecol. Res. 1999, 1, 49–71. [Google Scholar]
- Dukas, R.; Ellner, S. Information processing and prey detection. Ecology 1993, 74, 1337–1346. [Google Scholar] [CrossRef]
- Kotler, B.P.; Brown, J.S.; Hickey, M. Food storability and the foraging behavior of fox squirrels (Sciurus niger). Am. Midl. Nat. 1999, 142, 77–86. [Google Scholar] [CrossRef]
- Luttbeg, B.; Ferrari, M.C.O.; Blumstein, D.T.; Chivers, D.P. Safety cues can give prey more valuable information than danger cues. Am. Nat. 2020, 195, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Bielby, J.; Cardillo, M.; Fritz, S.A.; O’Dell, J.; Orme, C.D.L.; Safi, K.; Sechrest, W.; Boakes, E.H.; Carbone, C.; et al. PanTHERIA: A species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology 2009, 90, 2648. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 June 2020).
- Byrd, R.H.; Lu, P.; Nocedal, J.; Zhu, C.A. limited memory algorithm for bound constrained optimization. SIAM J. Sci. Comp. 1995, 16, 1190–1208. [Google Scholar] [CrossRef]
- Mullen, K.M.; Ardia, D.; Gil, D.; Windover, D.; Cline, J. DEoptim: An R Package for Global Optimization by Differential Evolution. J. Stat. Softw. 2011, 40, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Charnov, E.L. Optimal foraging: The marginal value theorem. Theor. Popul. Biol. 1976, 9, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, M.; Willoughby, J.R.; Lichti, N.I.; Steele, M.A.; Swihart, R.K. Segregating the effects of seed traits and common ancestry of hardwood trees on eastern gray squirrel foraging decisions. PLoS ONE 2015, 10, e0130942. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, M.; Lichti, N.I.; Widmar, N.O.; Swihart, R.K. Insights from an application of discrete choice experiments: Eastern gray squirrels are consistent shoppers of seed traits. Integr. Zool. 2018, 13, 280–296. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- McNamara, J.M.; Houston, A.I. Partial preferences and foraging. Anim. Behav. 1987, 35, 1084–1099. [Google Scholar] [CrossRef]
- Lima, S.L. Stress and decision making under the risk of predation: Recent developments from behavioral, reproductive, and ecological perspectives. Adv. Study Behav. 1998, 27, 215–290. [Google Scholar]
- Theimer, T.C. Rodent scatterhoarders as conditional mutualists. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CAB International: Wallingford, UK, 2005; pp. 283–296. [Google Scholar]
- Zwolak, R.; Crone, E.E. Quantifying the outcome of plant–granivore interactions. Oikos 2012, 121, 20–27. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Dalling, J.W.; Davis, A.S.; Schutte, B.J.; Arnold, A.E. Seed survival in soil: Interacting effects of predation, dormancy and the soil microbial community. J. Ecol. 2011, 99, 89–95. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lichti, N.I.; Dalgleish, H.J.; Steele, M.A. Interactions among Shade, Caching Behavior, and Predation Risk May Drive Seed Trait Evolution in Scatter-Hoarded Plants. Diversity 2020, 12, 416. https://doi.org/10.3390/d12110416
Lichti NI, Dalgleish HJ, Steele MA. Interactions among Shade, Caching Behavior, and Predation Risk May Drive Seed Trait Evolution in Scatter-Hoarded Plants. Diversity. 2020; 12(11):416. https://doi.org/10.3390/d12110416
Chicago/Turabian StyleLichti, Nathanael I., Harmony J. Dalgleish, and Michael A. Steele. 2020. "Interactions among Shade, Caching Behavior, and Predation Risk May Drive Seed Trait Evolution in Scatter-Hoarded Plants" Diversity 12, no. 11: 416. https://doi.org/10.3390/d12110416