Diversity of Species and Susceptibility Phenotypes toward Commercially Available Fungicides of Cultivable Fungi Colonizing Bones of Ursus spelaeus on Display in Niedźwiedzia Cave (Kletno, Poland)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Chemicals
2.3. Visual Inspection with Direct Microscopical Examination
2.4. Culture Conditions
2.5. In Vitro Susceptibility Testing
2.6. Molecular Identification
2.7. Data Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Gostinčar, C.; Grube, M.; De Hoog, S.; Zalar, P.; Gunde-Cimerman, N. Extremotolerance in fungi: Evolution on the edge. FEMS Microbiol. Ecol. 2010, 71, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.J.; Li, J.J.; Liang, Z.Z.; Zhao, C.Q. Novel Natural Products from Extremophilic Fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Adetutu, E.M.; Ball, A.S. Microbial diversity and activity in caves. Microbiol. Aust. 2014, 35, 192–194. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhao, P.; Cai, L. Origin of cave fungi. Front. Microbiol. 2018, 9, 1407. [Google Scholar] [CrossRef]
- Candiroglu, B.; Dogruoz-Gungor, N. Cave Ecosystems: Microbiological View. Eur. J. Biol. 2017, 76, 36–42. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Xue, C.; Idnurm, A.; Rutherford, J.C.; Heitman, J.; Cardenas, M.E. Sensing the environment: Lessons from fungi. Nat. Rev. Microbiol. 2007, 5, 57–69. [Google Scholar] [CrossRef]
- Sephton-Clark, P.C.S.; Voelz, K. Spore Germination of Pathogenic Filamentous Fungi. Adv. Appl. Microbiol. 2018, 102, 117–157. [Google Scholar]
- Hassan, N.; Rafiq, M.; Hayat, M.; Shah, A.A.; Hasan, F. Psychrophilic and psychrotrophic fungi: A comprehensive review. Rev. Environ. Sci. Biotechnol. 2016, 15, 147–172. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Cave features, seasonality and subterranean distribution of non-obligate cave dwellers. PeerJ 2017, 5, e3169. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Lejman, A.; Matkowski, K. Fungi isolated from Niedźwiedzia Cave in Kletno (Lower Silesia, Poland). Int. J. Speleol. 2013, 42, 161–166. [Google Scholar] [CrossRef]
- Nováková, A.; Kubátová, A.; Sklenář, F.; Hubka, V. Microscopic fungi on cadavers and skeletons from cave and mine environments. Czech Mycol. 2018, 70, 101–121. [Google Scholar]
- Borda, D.R.; Năstase-Bucur, R.M.; Spȋnu, M.; Uricariu, R.; Mulec, J. Aerosolized microbes from organic rich materials: Case study of bat guano from caves in Romania. J. Cave Karst Stud. 2014, 76, 114–126. [Google Scholar] [CrossRef]
- Alfaro, A. Psychrophilic and psychrotolerant fungi on bats and the presence of Geomyces spp. on bat wings prior to the arrival of white nose syndrome. Appl. Environ. Microb. 2013, 79, 54–65. [Google Scholar]
- Oliveira, M.P.A.; Bernardi, L.F.O.; Zeppelini, D.; Ferreira, R.L. First report of cave springtail (Collembola, Paronellidae) parasitized by mite (Parasitengona, Microtrombidiidae). Subterr. Biol. 2016, 17, 133–139. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.; Novaákovaá, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. The impact of arthropods on fungal community structure in Lascaux Cave. J. Appl. Microbiol. 2009, 106, 1456–1462. [Google Scholar] [CrossRef]
- Shapiro, J.; Pringle, A. Anthropogenic influences on the diversity of fungi isolated from caves in Kentucky and Tennessee. Am. Midl. Nat. 2010, 163, 76–86. [Google Scholar] [CrossRef]
- Bastian, F.; Jurado, V.; Nováková, A.; Alabouvette, C.; Saiz-Jimenez, C. The microbiology of Lascaux Cave. Microbiology 2010, 156, 644–652. [Google Scholar] [CrossRef]
- Griffin, D.W.; Gray, M.A.; Lyles, M.B.; Northup, D.E. The Transport of Nonindigenous Microorganisms into Caves by Human Visitation: A Case Study at Carlsbad Caverns National Park. Geomicrobiol. J. 2014, 31, 175–185. [Google Scholar]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Cultivable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Pusz, W.; Król, M.; Zwijacz-Kozica, T. Airborne fungi as indicators of ecosystem disturbance: An example from selected Tatra Mountains caves (Poland). Aerobiologia 2018, 34, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Burford, E.P.; Fomina, M.; Gadd, G.M. Fungal involvement in bioweathering and biotransformation of rocks and minerals. Miner. Mag. 2003, 67, 1127–1155. [Google Scholar]
- Dupont, J.; Jacquet, C.; Dennetiere, B.; Lacoste, S.; Bousta, F.; Orial, G.; Cruaud, C.; Couloux, A.; Roquebert, M.F. Invasion of the French Paleolithic painted cave of Lascaux by members of the Fusarium solani species complex. Mycologia 2007, 99, 526–533. [Google Scholar] [CrossRef]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeterior. Biodegrad. 2008, 62, 325–330. [Google Scholar]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biotechnol. 2013, 97, 9637–9646. [Google Scholar] [CrossRef]
- Salyers, A.A.; Whitt, D.D.; Wessner, D.; Elmendorf, H.; Stephens, C.M. Microbiology: Diversity, Disease, and the Environment, 2nd ed.; Fitzgerald Science Press, Inc.: Bethesda, MD, USA, 2001; pp. 297–298. [Google Scholar]
- Official Website of Niedźwiedzia Cave. Available online: http://jaskinianiedzwiedzia.pl/ (accessed on 21 October 2019). (In Polish).
- Devor Mousse (Product Specification Sheet). Available online: http://oxi-peintures.com/media/ft_antimousse_concentre_devor_mousse__034946500_1601_20082013 (accessed on 21 October 2019).
- Altax (Product Specification Sheet). Available online: http://www.promesa-farby.pl/download/Karty%20±1oCharakterystyki%20i%20atesty/ALTAX/Karty%20±1oCharakterystyki/BORAMON%20%C2%8CRODEK%20GRZYBOB%C3%93JCZY_karta%20±1oCharakterystyki.pdf (accessed on 21 October 2019).
- Pufmax (Product Specification Sheet). Available online: http://www.cherub.com.pl/upload/impregnaty//Dokumenty//PUFMAKS-POGROMCA-PLESNI-v-3-22-05-2014.pdf (accessed on 21 October 2019).
- McGinnis, M.R. Laboratory Handbook of Medical Mycology; Academic Press, Inc.: New York, NY, USA, 1980; pp. 132–133. [Google Scholar]
- Nishimoto, K.; Takemoto, H. A simple staining and mounting method of KOH preparations for mycological examination using Parker-blue-black ink. Jpn. J. Med. Mycol. 1979, 20, 145–147. [Google Scholar] [CrossRef]
- Garrett, S.D. Soil Fungi and Soil Fertility: An Introduction to Soil Mycology, 2nd ed.; Pergamon Press: Oxford, UK, 1981; pp. 1–64. [Google Scholar]
- Choi, Y.W.; Hyde, K.D.; Ho, W.H. Single spore isolation of fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compedium of Soil Fungi, 2nd ed.; IHW Verlag: Eching, Germany, 2007; pp. 1–672. [Google Scholar]
- de Hoog, G.S. Atlas of Clinical Fungi, 2nd ed.; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2000; pp. 1–1126. [Google Scholar]
- Samson, R.A.; Varga, J. Aspergillus Systematics in the Genomic Era; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2017. [Google Scholar]
- Samson, R.A.; Varga, J.; Frisvad, J.C. Taxonomic studies on the genus Aspergillus. Stud. Mycol. 2011, 69, 1–97. [Google Scholar]
- Samson, R.A.; Houbraken, J. Phylogenetic and taxonomic studies on the genera Penicillium and Talaromyces. Stud. Mycol. 2011, 70, iii. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Samson, R.A.; Cobus, M.; Visagie, C.M.; Houbraken, J. Species Diversity in Aspergillus, Penicillium and Talaromyces; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2014. [Google Scholar]
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes; CBS Biodiversity Series; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; pp. 1–997. [Google Scholar]
- Guarro, J.; Gene, J.; Stchigel, A.M.; Figueras, M.J. Atlas of Soil Ascomycetes; CBS Biodiversity Series; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; Volume 10, pp. 1–486. [Google Scholar]
- Hubka, V.; Nováková, A.; Peterson, S.W.; Frisvad, J.; Sklenár, F.; Matsuzawa, T.; Kubátová, A.; Kolarik, M. A reappraisal of Aspergillus section Nidulantes with descriptions of two new sterigmatocystin-producing species. Plant Syst. Evol. 2016, 302, 1267–1299. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Guarro, J.; Cano-Lira, J.F.; Sutton, D.A.; Wiederhold, N.P.; de Hoog, G.S.; Abbott, S.P.; Decock, C.; Sigler, L.; Gene, J. Phylogeny and taxonomic revision of Microascaceae with emphasis on synnematous fungi. Stud. Mycol. 2016, 83, 193–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.J.; Frisvad, J.C.; Sun, B.D.; Varga, J.; Kocsube, S.; Dijksterhuis, J.; Kim, D.H.; Hong, S.B.; Houbraken, J.; Samson, R.A. Aspergillus section Nidulantes (formerly Emericella): Polyphasic taxonomy, chemistry and biology. Stud. Mycol. 2016, 84, 1–118. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi-Approved Standard, CLSI Document M38-A2, 2nd ed.; CLSI: Wayne, PA, USA, 2008; pp. 3–30. [Google Scholar]
- Kaiser, C.; Michaelis, S.; Mitchel, A.A. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1994; pp. 195–197. [Google Scholar]
- Espinel-Ingroff, A.; Walsh, T.J.; Fothergill, A.; Peter, J.; Rinaldi, M.G.; Walsh, T.J. Testing conditions for determination of Minimum Fungicidal Concentrations of new and established antifungal agents for Aspergillus spp. NCCLS collaborative study. J. Clin. Microbiol. 2002, 40, 3204–3208. [Google Scholar] [CrossRef]
- Dyląg, M.; Pruchnik, H.; Pruchnik, F.; Majkowska-Skrobek, G.; Ułaszewski, S. Antifungal activity of organotin compounds with functionalized carboxylates evaluated by the micro-dilution bioassay in vitro. Med. Mycol. 2010, 48, 373–383. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, X.; Wen, H.; Wang, M. A simple method of genomic DNA extraction suitable for analysis of bulk fungal strains. Lett. Appl. Microbiol. 2010, 51, 114–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bliss, C.T. The method of probits. Science 1934, 79, 38–39. [Google Scholar] [CrossRef]
- Ana-Maria, B.; Ion, S. Monitoring of pollutants in museum environment. PESD 2015, 9, 173–180. [Google Scholar] [CrossRef]
- Nittérus, M. Fungi in archives and libraries, a literary survey. Restaurator 2000, 21, 25–40. [Google Scholar]
- Montanari, M.; Melloni, V.; Pinzari, F.; Innocenti, G. Fungal biodeterioration of historical library materials stored in Compactus movable shelves. Int. Biodeterior. Biodegrad. 2012, 75, 83–88. [Google Scholar] [CrossRef]
- Hueck, H.J. The biodeterioration of materials—An appraisal. Int Biodeterior. Biodegrad. 2001, 48, 5–11. [Google Scholar] [CrossRef]
- Morton, L.H.G. Things that go rot in the night—A review of biodeterioration. Microbiol. Today 2003, 30, 103–106. [Google Scholar]
- Child, A.M. Microbial taphonomy of archaeological bone. Stud. Conserv. 1995, 40, 19–30. [Google Scholar]
- Jans, M.M. Microbial bioerosion of bone—A review. Current Developments in Bioerosion. In Erlangen Earth Conference Series, 2nd ed.; Wisshak, M., Tapanila, L., Eds.; Springer: Berlin, Germany, 2008; pp. 397–413. [Google Scholar]
- Gutarowska, B.; Czyżowska, A. The ability of filamentous fungi to produce acids on indoor building materials. Ann. Microbiol. 2009, 59, 807–813. [Google Scholar] [CrossRef]
- Sazanova, K.; Shchiparev, S.; Vlasov, D. Formation of organic acids by fungi isolated from the surface of stone monuments. Microbiology 2014, 83, 516–522. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Slack, G.J.; Puniani, E.; Frisvad, J.C.; Samson, R.A.; Miller, J.D. Secondary metabolites from Eurotium species, Aspergillus calidoustus and A. insuetus common in Canadian homes with a review of their chemistry and biological activities. Mycol. Res. 2009, 113, 480–490. [Google Scholar] [CrossRef]
- Duarte, A.; dos Santos, J.A.; Vianna, M.V.; Freitas-Vieira, J.M.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, V.; DeFiglio, H.; Chaturvedi, S. Phenotype profiling of white-nose syndrome pathogen Pseudogymnoascus destructans and closely-related Pseudogymnoascus pannorum reveals metabolic differences underlying fungal lifestyles. F1000Research 2018, 7, e665. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Alias, A.S.; Wong, C.M.V.L.; Ka-Lai Pang, K.-L.; Convey, P. Extracellular hydrolase enzyme production by soil fungi from King George Island, Antarctica. Polar Biol. 2011, 34, 1535–1542. [Google Scholar] [CrossRef]
- Riordan, F. Minimum conditions for visible mold growth. ASHRAE J. 2016, 58, 32–43. [Google Scholar]
- Jans, M.M.E.; Nielsen-Marsh, C.M.; Smith, C.I.; Collins, M.J.; Kars, H. The characterisation of microbial attack in archaeological bone. J. Archaeol. Sci. 2004, 31, 87–95. [Google Scholar] [CrossRef]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.; Discher, S.; Voigt, K. Revision of the genus Absidia (Mucorales, Zygomycetes) based on physiological, phylogenetic, and morphological characters; thermotolerant Absidia spp. form a coherent group, Mycocladiaceae fam. nov. Mycol. Res. 2007, 111, 1169–1183. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Sutton, D.A.; Fothergill, A.W.; Cano-Lira, J.; Gené, J.; Decock, C.A.; de Hoog, G.S.; Guarro, J. Scopulariopsis, a poorly known opportunistic fungus: Spectrum of species in clinical samples and in vitro responses to antifungal drugs. J. Clin. Microbiol. 2013, 51, 3937–3943. [Google Scholar] [CrossRef] [Green Version]
- Tola, M.; Kebede, B.; Yildiz, F. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Block, S.S. Disinfection, Sterilization, and Preservation, 5th ed.; Lippincott Williams and Wilkins: Walnut Street, PA, USA, 2001; pp. 110–1320. [Google Scholar]
- Szczepanowska, H.M. Conservation of Cultural Heritage: Key Principles and Approaches, 1st ed.; Routledge: London, UK, 2013; pp. 12–234. [Google Scholar]
- Diaz-Herraiz, M.; Jurado, V.; Cuezva, S.; Laiz, L.; Pallecchi, P.; Tiano, P.; Sanchez-Moral, S.; Saiz-Jimenez, C. The actinobacterial colonization of Etruscan paintings. Sci. Rep. 2013, 3, 1440. [Google Scholar] [CrossRef] [Green Version]
- Brycki, B. Gemini alkylammonium salts as biodeterioration inhibitors. Pol. J. Microbiol. 2010, 59, 227–231. [Google Scholar] [PubMed]
- Perinelli, D.R.; Petrelli, D.; Vitali, L.A.; Vllasaliu, D.; Cespi, M.; Giorgioni, G.; Elmowafy, E.; Bonacucina, G.; Palmieri, G.F. Quaternary ammonium surfactants derived from leucine and methionine: Novel challenging surface active molecules with antimicrobial activity. J. Mol. Liq. 2019, 283, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Gallo, F. Aerobiological research and problems in libraries. Aerobiologia 1993, 9, 117–130. [Google Scholar] [CrossRef]
- Kompatscher, K.; Kramer, R.P.; Ankersmit, B.; Schellen, H.L. Intermittent conditioning of library archives: Microclimate analysis and energy impact. Build. Environ. 2019, 147, 50–66. [Google Scholar] [CrossRef]
- de Freitas, C.R. The role and importance of cave microclimate in the sustainable use and management of show caves. Acta Carsologica 2010, 39, 477–489. [Google Scholar] [CrossRef]
- Cigna, A.A.; Forti, P. Caves: The most important geotouristic feature in the world. Tour. Karst Areas 2013, 6, 9–26. [Google Scholar]






{kind=link}
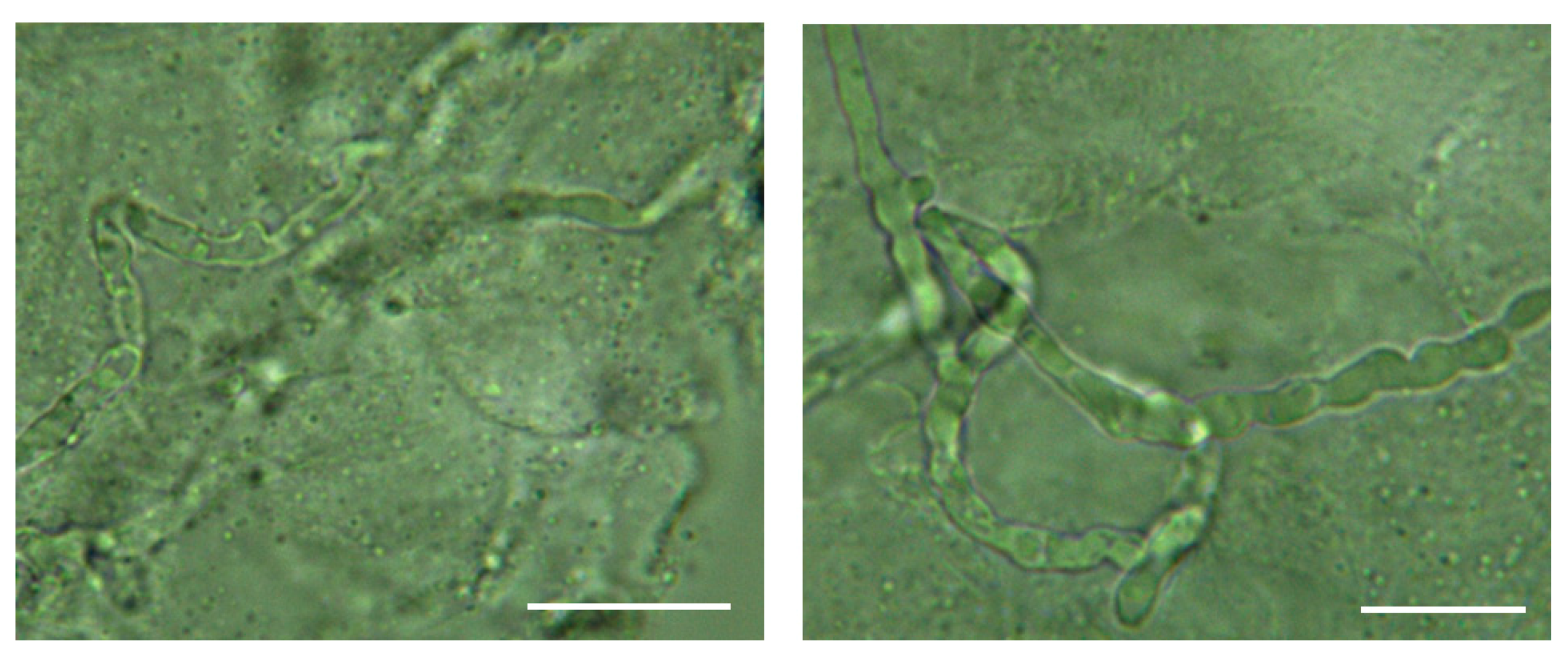{kind=link}
{kind=link}
{kind=link}
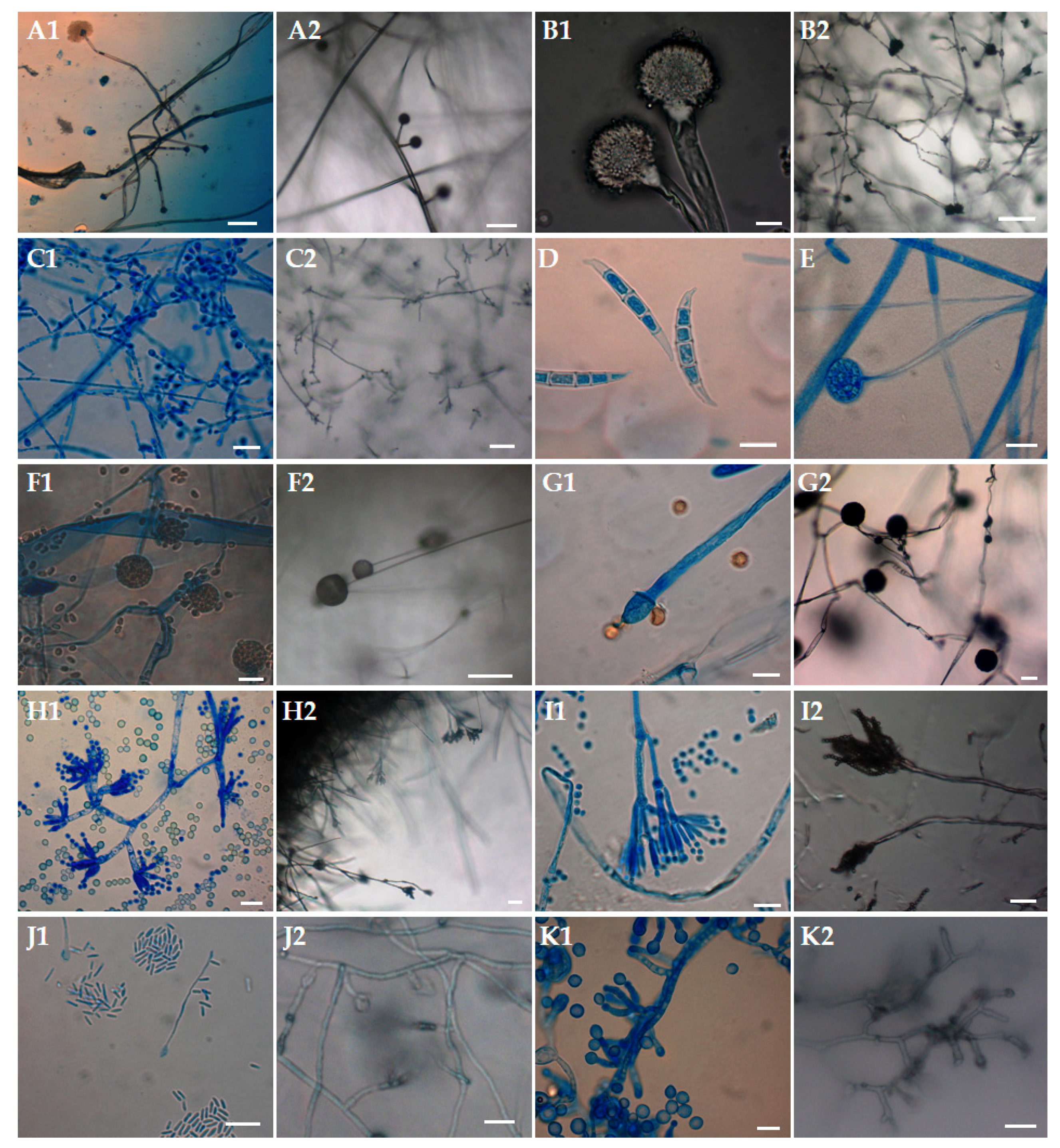{kind=link}
| Fungal Strains Isolated from Ursus spelaeus Bones | Identity with Sequence from GenBank | ||||
|---|---|---|---|---|---|
| Identified Species | GenBank Accession No. | The Sequence Length (bp) | Query Cover (%) | Identity (%) | Accession |
| Absidia glauca | KY465754.1 | 607 | 96 | 94.71 | MK690542.1 |
| Aspergillus fumigatus | KY465755.1 | 551 | 100 | 99.64 | LC333286.1 |
| Chrysosporium merdarium | KY465756.1 | 539 | 99 | 97.40 | KF225604.1 |
| Fusarium cerealis | KY465757.1 | 504 | 99 | 100.00 | MG979793.1 |
| Mortierella alpina | KY465758.1 | 636 | 99 | 98.12 | MG833809.1 |
| Mucor aligarensis | KY465759.1 | 648 | 99 | 99.38 | KM396375.1 |
| Mucor plumbeus | KY465760.1 | 610 | 98 | 98.17 | MK268150.1 |
| Penicillium chrysogenum | KY465761.1 | 548 | 100 | 99.82 | KU743899.1 |
| Penicillium expansum | KY465762.1 | 587 | 96 | 95.61 | MH630060.1 |
| Sarocladium strictum | KY465763.1 | 555 | 99 | 99.10 | KT878337.1 |
| Scopulariopsis candida | KY465764.1 | 545 | 98 | 99.81 | JN942891.1 |
| Species | Values of Active Concentrations (%) 1 | ||||||
|---|---|---|---|---|---|---|---|
| Pufmax | Devor Mousse | Boramon | |||||
| MIC 2 | MFC 3 | MIC | MFC | MIC | MFC | ||
| A. glauca | 0.63 ± 0.04 a 4 | 2.50 ± 0.30 a | 2.50 ± 0.03 a | 5.00 ± 0.05 a | 10.00 ± 2.00 a | 20.00 ± 3.00 a | |
| A. fumigatus | 0.31 ± 0.02 b | 1.25 ± 0.05 b | 0.63 ± 0.11 b | 2.50 ± 0.00 b | 5.00 ± 0.50 b | 10.00 ± 1.50 b | |
| C. merdarium | 0.16 ± 0.02 c | 0.63 ± 0.03 c | 0.31 ± 0.03 c | 1.25 ± 0.31 b | 2.50 ± 0.05 b | 5.00 ± 1.00 c | |
| F. cerealis | 0.63 ± 0.03 a | 2.50 ± 0.20 a | 0.63 ± 0.11 b | 2.50 ± 0.50 b | 5.00 ± 1.00 b | 10.00 ± 0.00 b | |
| M. alpina | 0.63 ± 0.05 a | 2.50 ± 0.10 a | 2.50 ± 0.10 a | 5.00 ± 0.50 a | 10.00 ± 1.50 a | 20.00 ± 0.87 a | |
| M. aligarensis | 0.63 ± 0.01 a | 2.50 ± 0.30 a | 2.50 ± 0.20 a | 5.00 ± 1.32 a | 10.00 ± 1.32 a | 20.00 ± 1.32 a | |
| M. plumbeus | 0.63 ± 0.04 a | 2.50 ± 0.36 a | 2.50 ± 0.36 a | 5.00 ± 1.00 a | 10.00 ± 1.00 a | 20.00 ± 1.00 a | |
| P. chrysogenum | 0.31 ± 0.03 b | 2.50 ± 0.10 a | 0.63 ± 0.05 b | 2.50 ± 0.40 b | 5.00 ± 1.00 b | 10.00 ± 1.73 b | |
| P. expansum | 0.31 ± 0.01 b | 2.50 ± 0.20 a | 0.63 ± 0.03 b | 2.50 ± 0.20 b | 2.50 ± 0.05 b | 5.00 ± 0.87 c | |
| S. strictum | 0.31 ± 0.02 b | 1.25 ± 0.09 b | 0.63 ± 0.06 b | 2.50 ± 0.20 b | 2.50 ± 0.05 b | 10.00 ± 1.73 b | |
| S. candida | 0.63 ± 0.03 a | 2.50 ± 0.10 a | 0.63 ± 0.04 b | 2.50 ± 0.10 b | 5.00 ± 1.73 b | 10.00 ± 0.87 b | |
| Mean | (%) | 0.47 ± 0.18 C4-1 | 2.10 ± 0.70 B4-2 | 1.28 ± 0.95 B4-1 | 3.34 ± 1.50 B4-2 | 6.14 ± 3.28 A4-1 | 12.73 ± 6.00 A4-2 |
| (µg · mL−1) | 22.60 ± 0.86 | 100.80 ± 3.36 | 64.00 ± 9.00/32 ± 4.5 | 167.00 ± 7.5/83.50 ± 3.75 | 202.60 ± 10.80 | 420.00 ± 5.90 | |
| Species | Colony Growth Inhibition (%)1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pufmax | Devor Mousse | Boramon | |||||||
| 0.1% (4.8) 2 | 0.5% (24.0) | 1.0% (48.0) | 0.1% (5.0/2.5) | 0.5% (25.0/12.5) | 1.0% (50.0/25.0) | 0.1% (3.3) | 0.5% (16.5) | 1.0% (33.0) | |
| A. glauca | 54 ± 3 d3 | 100 ± 0 a | 100 ± 0 a | 30 ± 1 ef | 65 ± 3 b | 100 ± 0 a | 35 ± 1 de | 65 ± 2 def | 100 ± 0 a |
| A. fumigatus | 75 ± 6 c | 100 ± 0 a | 100 ± 0 a | 53 ± 3 c | 100 ± 0 a | 100 ± 0 a | 55 ± 3 b | 90 ± 4 b | 100 ± 0 a |
| C. merdarium | 100 ± 0 a | 100 ± 0 a | 100 ± 0 a | 93 ± 3 a | 100 ± 0 a | 100 ± 0 a | 79 ± 1 a | 100 ± 0 a | 100 ± 0 a |
| F. cerealis | 53 ± 1 d | 100 ± 0 a | 100 ± 0 a | 45 ± 1 cd | 100 ± 0 a | 100 ± 0 a | 33 ± 4.6 e | 68 ± 5 de | 100 ± 0 a |
| M. alpina | 48 ± 2 d | 100 ± 0 a | 100 ± 0 a | 20 ± 3 f | 67 ± 5 b | 100 ± 0 a | 40 ± 4 cde | 84 ± 4 bc | 100 ± 0 a |
| M. aligarensis | 46 ± 1 d | 100 ± 0 a | 100 ± 0 a | 32 ± 1 de | 70 ± 2 b | 100 ± 0 a | 38 ± 8 cde | 58 ± 7 ef | 100 ± 0 a |
| M. plumbeus | 56 ± 2 d | 100 ± 0 a | 100 ± 0 a | 28 ± 3 ef | 69 ± 2 b | 100 ± 0 a | 48 ± 2 bc | 86 ± 2 bc | 100 ± 0 a |
| P. chrysogenum | 78 ± 2 c | 100 ± 0 a | 100 ± 0 a | 48 ± 4 c | 100 ± 0 a | 100 ± 0 a | 34 ± 2 de | 85 ± 3 bc | 100 ± 0 a |
| P. expansum | 84 ± 4 bc | 100 ± 0 a | 100 ± 0 a | 86 ± 1 ab | 100 ± 0 a | 100 ± 0 a | 30 ± 4 e | 52 ± 2 f | 100 ± 0 a |
| S. strictum | 91 ± 6 b | 100 ± 0 a | 100 ± 0 a | 76 ± 6 b | 100 ± 0 a | 100 ± 0 a | 41 ± 2 cde | 65 ± 3 def | 100 ± 0 a |
| S. candida | 47 ± 2 d | 100 ± 0 a | 100 ± 0 a | 47 ± 5 c | 100 ± 0 a | 100 ± 0 a | 47 ± 1 bcd | 74 ± 2 cd | 100 ± 0 a |
| Mean | 66 ± 19 b4, A4-1, | 100 ± 0 a, A4-2 | 100 ± 0 a, A4-3 | 51 ± 24 c, B4-1 | 88 ± 16 b, B4-2 | 100 ± 0 a, A4-3 | 44 ± 14 c, B4-1 | 75 ± 15 b, C4-2 | 100 ± 0 a, A4-3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyląg, M.; Sawicki, A.; Ogórek, R. Diversity of Species and Susceptibility Phenotypes toward Commercially Available Fungicides of Cultivable Fungi Colonizing Bones of Ursus spelaeus on Display in Niedźwiedzia Cave (Kletno, Poland). Diversity 2019, 11, 224. https://doi.org/10.3390/d11120224
Dyląg M, Sawicki A, Ogórek R. Diversity of Species and Susceptibility Phenotypes toward Commercially Available Fungicides of Cultivable Fungi Colonizing Bones of Ursus spelaeus on Display in Niedźwiedzia Cave (Kletno, Poland). Diversity. 2019; 11(12):224. https://doi.org/10.3390/d11120224
Chicago/Turabian StyleDyląg, Mariusz, Artur Sawicki, and Rafał Ogórek. 2019. "Diversity of Species and Susceptibility Phenotypes toward Commercially Available Fungicides of Cultivable Fungi Colonizing Bones of Ursus spelaeus on Display in Niedźwiedzia Cave (Kletno, Poland)" Diversity 11, no. 12: 224. https://doi.org/10.3390/d11120224