Chloroplast Genome-Based Hypervariable Markers for Rapid Authentication of Six Korean Pyropia Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chloroplast Genome Comparison and Identification of Hypervariable InDels
2.2. Sample Collection and DNA Isolation
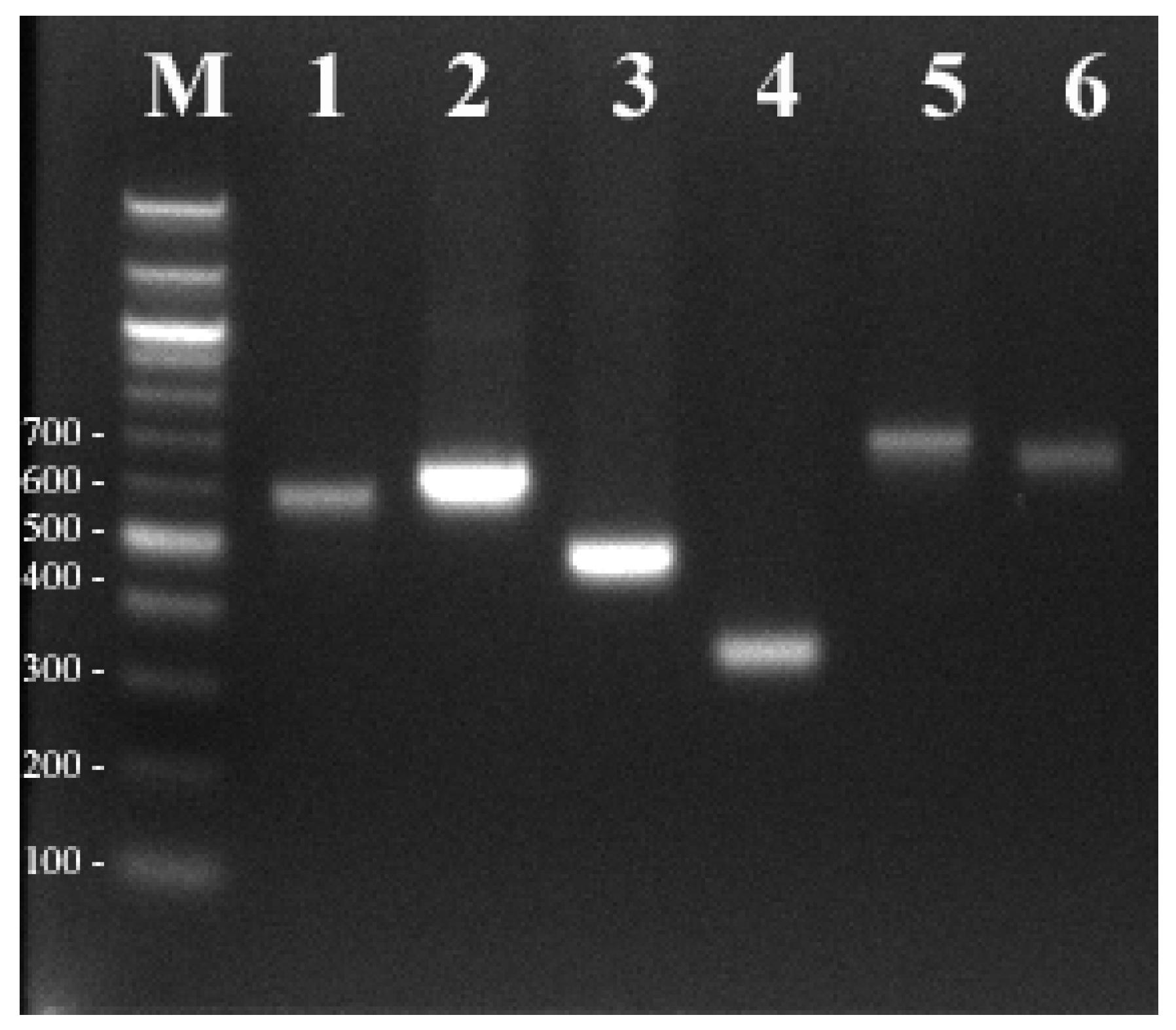
2.3. Development and Validation of the InDel Molecular Marker
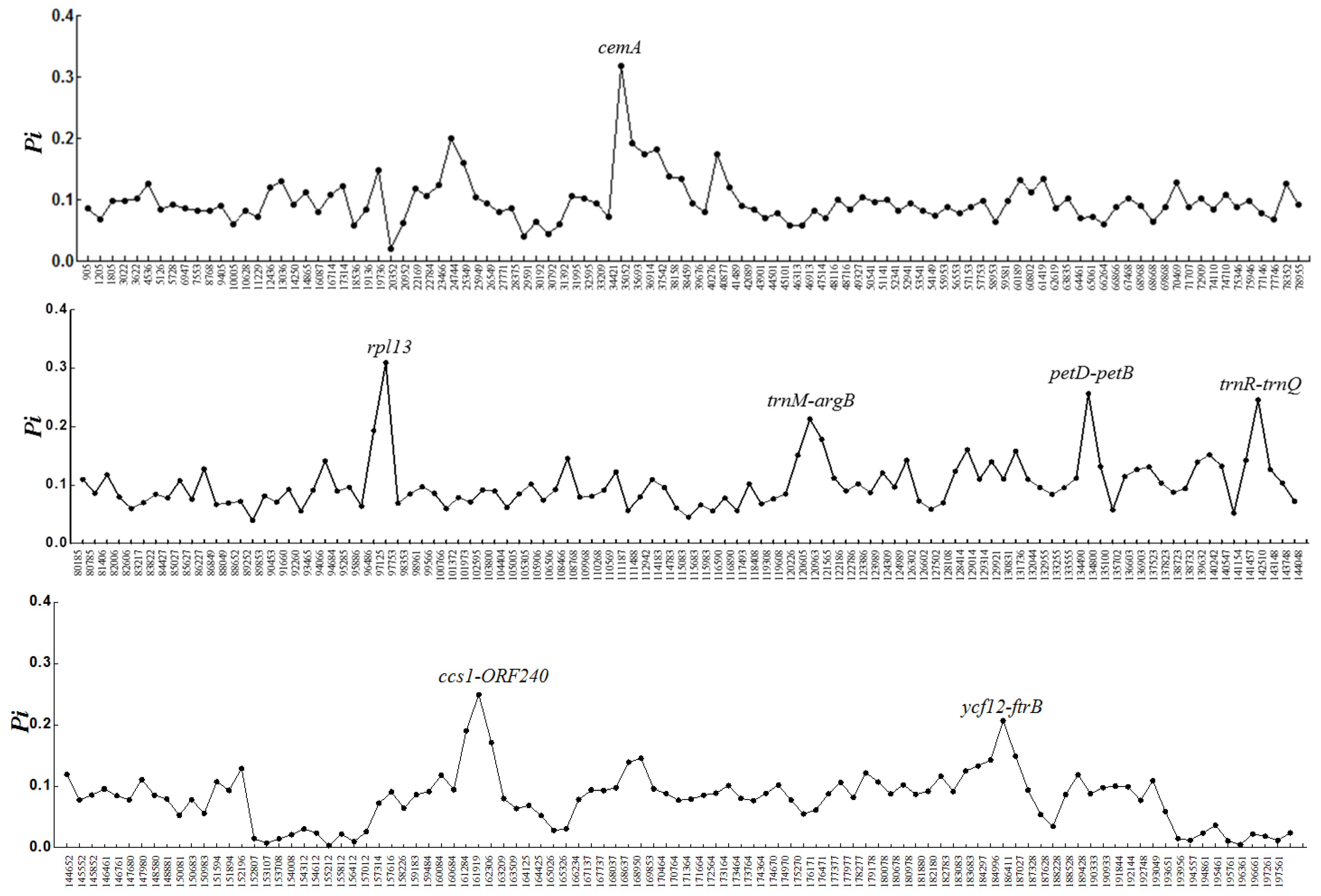
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vergés, A.; Sánchez, N.; Peteiro, C.; Polo, L.; Brodie, J. Pyropiasuborbiculata (Bangiales, Rhodophyta): First records from the northeastern Atlantic and Mediterranean of this North Pacific species. Phycologia 2013, 52, 121–129. [Google Scholar] [CrossRef]
- Hallmann, A. Algae biotechnology–green cell-factories on the rise. Curr. Biotechnol. 2015, 4, 389–415. [Google Scholar] [CrossRef]
- Hwang, M.-S.; Lee, I.G. Character analysis and numerical taxonomy of Porphyra (Bangiales, Rhodophyta) from Korea. Algae 2002, 17, 217–233. [Google Scholar] [CrossRef]
- Kim, Y.; Choi, S.J.; Choi, C. An Efficient PCR-RFLP Method for the Rapid Identification of Korean Pyropia Species. Molecules 2017, 22, 2182. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.W. Applying DNA barcoding to red macroalgae: A preliminary appraisal holds promise for future application. Philos. Trans. R. Soc. Ser. B 2005, 9, 1879–1888. [Google Scholar] [CrossRef] [PubMed]
- Hughey, J.; Silva, P.; Hommersand, M. Solving taxonomic and nomenclatural problems in Pacific Gigartinaceae (Rhodophyta) using DNA from type material. J. Phycol. 2001, 1109, 1091–1109. [Google Scholar] [CrossRef]
- Freshwater, D.W.; Tudor, K.; O’shaughnessy, K.; Wysor, B. DNA barcoding in the red algal order Gelidiales: Comparison of COI with rbcL and verification of the barcoding gap. Cryptogam. Algol. 2010, 31, 435–449. [Google Scholar]
- Yang, E.C.; Kim, M.S.; Geraldino, P.J.L.; Sahoo, D.; Shin, J.A.; Boo, S.M. Mitochondrial cox1 and plastid rbcL genes of Gracilariavermiculophylla (Gracilariaceae, Rhodophyta). J. Appl. Phycol. 2008, 20, 161–168. [Google Scholar] [CrossRef]
- Yang, E.C.; Boo, S.M. A red alga-specific phycoerythrin gene for biodiversity surveys of callithamnioid red algae. Mol. Ecol. Notes 2006, 6, 533–535. [Google Scholar] [CrossRef]
- Xu, K.; Tang, X.; Bi, G.; Cao, M.; Wang, L.; Mao, Y. The first complete organellar genomes of an Antarctic red alga, Pyropiaendiviifolia: Insights into its genome architecture and phylogenetic position within genus Pyropia (Bangiales, Rhodophyta). J. Oceanol. Limnol. 2018, 36, 1315–1328. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Research Group | NCBI Accessions of Chloroplast Genome | Sequence Length (bp) |
|---|---|---|---|---|
| 1 | P. yezoensis | National Research Institute of Fishery Science, Aquatic genomics research center, Kanagawa, Japan | NC007932 | 191,952 |
| 2 | P. haitanensis | National Center for Biotechnology Information, NIH, USA | NC021189 | 195,597 |
| 3 | P. endiviifolia | College of Marine Life Sciences, Ocean University, China | KT716756 | 195,784 |
| 4 | P. perforata | Math and Sciences, Hartnell College, Central Ave., USA | KC904971 | 189,789 |
| 5 | P. kanakaensis | KJ776836 | 189,931 | |
| 6 | P. fucicola | KJ776837 | 187,282 | |
| 7 | P. pulchra | Biological Sciences, Sungkyunkwan, University, Korea | KT266789 | 194,175 |
| No. | Scientific Name (n = 3) | Common Name | Collection Site | Location |
|---|---|---|---|---|
| 1 | P. yezoensis | Bangsamunuigim | Songji-myeon, Haenam-gun, Jeollanam-do | 34°21′05.92″ N 126°27′40.76″ E |
| 2 | P. dentata | Itbadidolgim | Yuldo-dong, Mokpo-si, Jeollanam-do | 34°48′13.22″ N 126°18′34.88″ E |
| 3 | P. seriata | Momunuidolgim | Songji-myeon, Haenam-gun, Jeollanam-do | 34°45′49.37″ N 126°07′50.54″ E |
| 4 | P. suborbiculata | Dunggeundolgim | Nam-myeon, Yeosu-si, Jeollanam-do | 34°25′32.73″ N 127°47′31.33″ E |
| 5 | P. pseudolinearis | Ginipdolgim | Ulleung-gun, Gyeongsangbuk-do | 37°27′31.55″ N 130°54′14.98″ E |
| 6 | P. haitanensis | Haitanensisgim | Dried laver product from China |
| No. | Locus | Location | Forward Primer (Sequence 5′ to 3′) | Reverse Primer (Sequence 5′ to 3′) | Product Range (bp) | AS | No. of InDels | Mean Pairwise Distance |
|---|---|---|---|---|---|---|---|---|
| 1 | cemA | 35053..35693 | ATTGCAATTTGNCTTTGTCCAG | GAAAAAGTTGGGCCAATACCTA | 506–529 | 100 | 4 | 0.108 |
| 2 | rpl13 | 96807..97443 | AACACCNTTAACTGCATTACGTT | GTCACNGAAAAGTCATGGTAATT | 583–600 | 83.3 | N/A | N/A |
| 3 | trnM-argB | 120227..120963 | TGAGCTACTGAGCCATAATA | CTGATCAAGGTATTGGCTCGAT | 341–696 | 100 | 51 | 0.295 |
| 4 | petD-petB | 134166..134800 | CTTCTAAAAGGATTTTGAAACTT | CAGATGCTGTTCCAGTTGTTGGA | 629 | 50 | N/A | N/A |
| 5 | trnR-trnQ | 141451..143365 | GGTTGTAGCTCAGANGGATAG | GGGTGTAGCCAAGTGGTAAG | 534–1260 | 100 | 37 | 0.572 |
| 6 | ccs1-orf24 | 161285..162306 | TGTTCAATAATAGTTCCTATAATGC | TGGAATAATCTNTGGGCTCCTTT | 870–916 | 66.6 | N/A | N/A |
| 7 | ycf12-ftrB | 184298..186411 | GAAAAGAGGCAATCTTTAGTAAT | TGGAACTGNCCATGTGTACCAATG | 583 | 16.6 | N/A | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.-J.; Kim, Y.; Choi, C. Chloroplast Genome-Based Hypervariable Markers for Rapid Authentication of Six Korean Pyropia Species. Diversity 2019, 11, 220. https://doi.org/10.3390/d11120220
Choi S-J, Kim Y, Choi C. Chloroplast Genome-Based Hypervariable Markers for Rapid Authentication of Six Korean Pyropia Species. Diversity. 2019; 11(12):220. https://doi.org/10.3390/d11120220
Chicago/Turabian StyleChoi, Sung-Je, Yonguk Kim, and Chulyung Choi. 2019. "Chloroplast Genome-Based Hypervariable Markers for Rapid Authentication of Six Korean Pyropia Species" Diversity 11, no. 12: 220. https://doi.org/10.3390/d11120220