
AESIS-1, a Rheumatoid Arthritis Therapeutic Peptide, Accelerates Wound Healing by Promoting Fibroblast Migration in a CXCR2-Dependent Manner

Abstract
:1. Introduction
2. Results
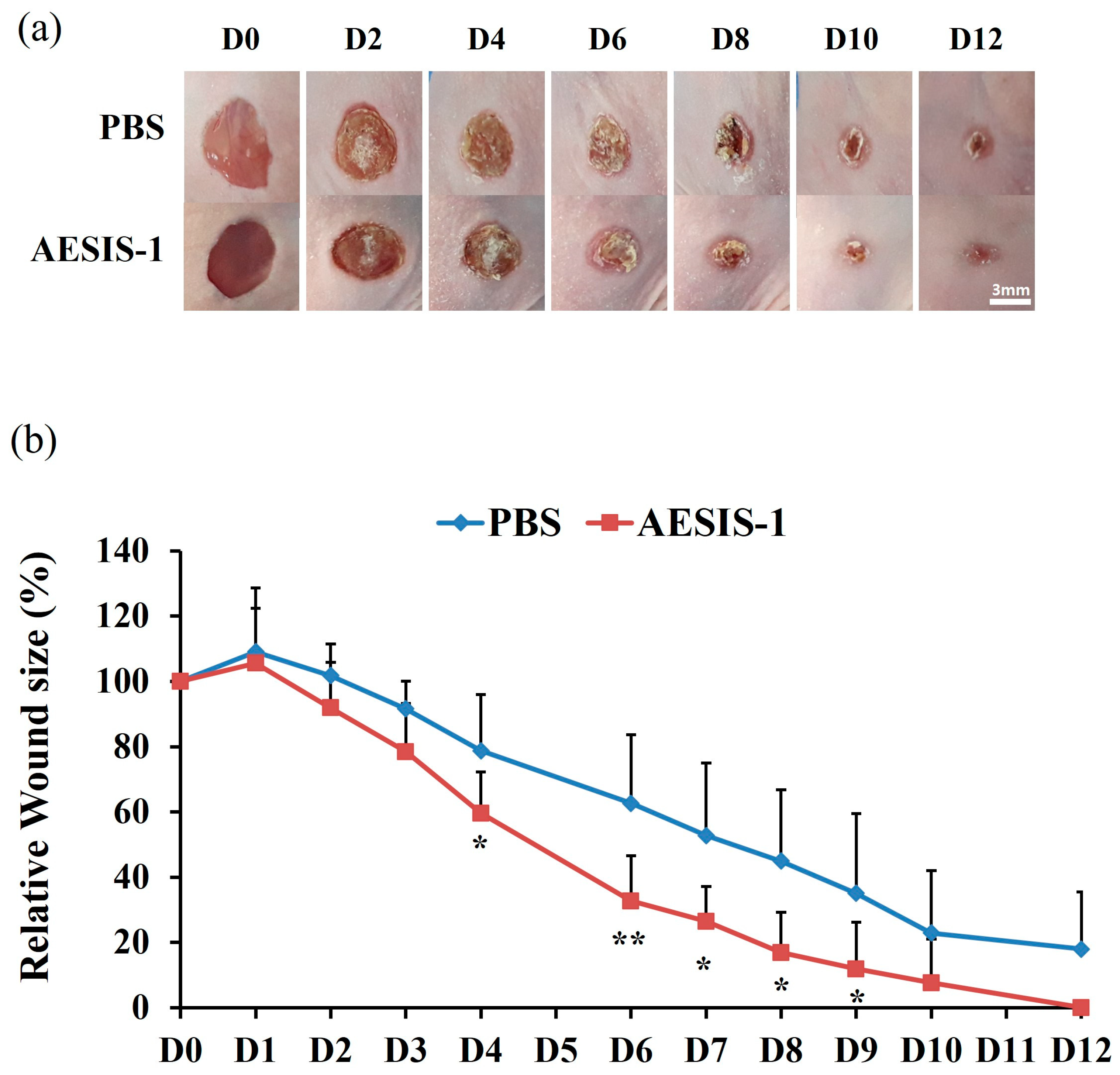
2.1. AESIS-1 Exerted Acceleration of Wound Healing In Vivo
2.2. AESIS-1 Increases the Migration Activity of Fibroblasts without Affecting Fibroblast Proliferation
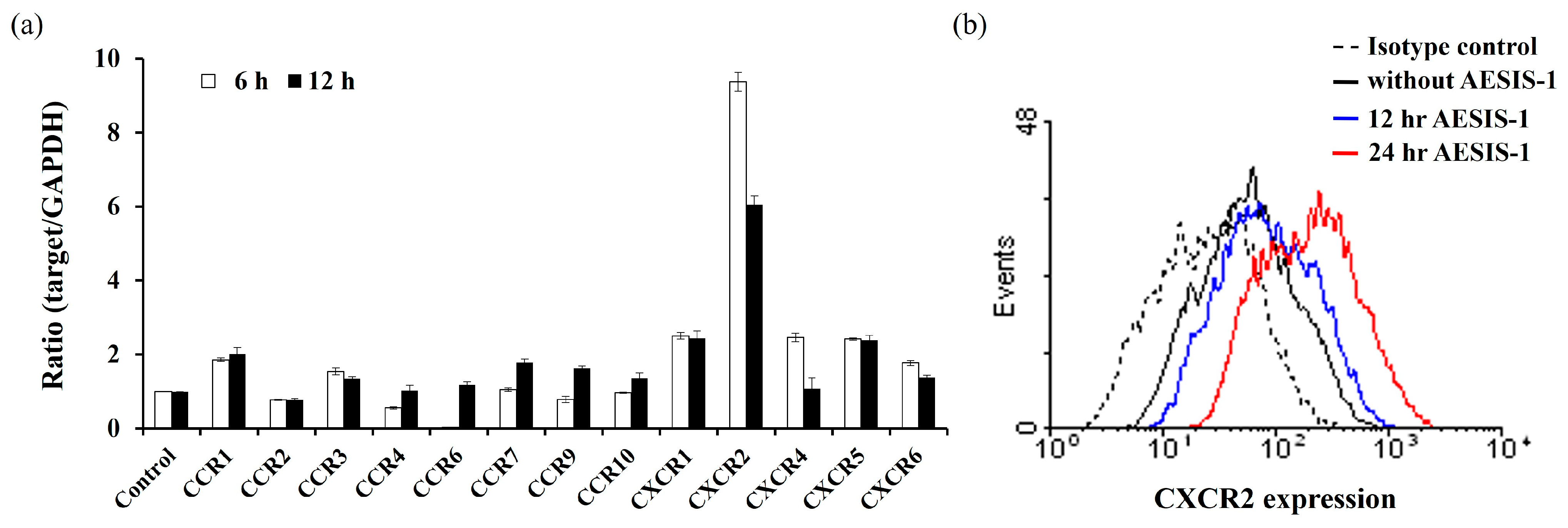
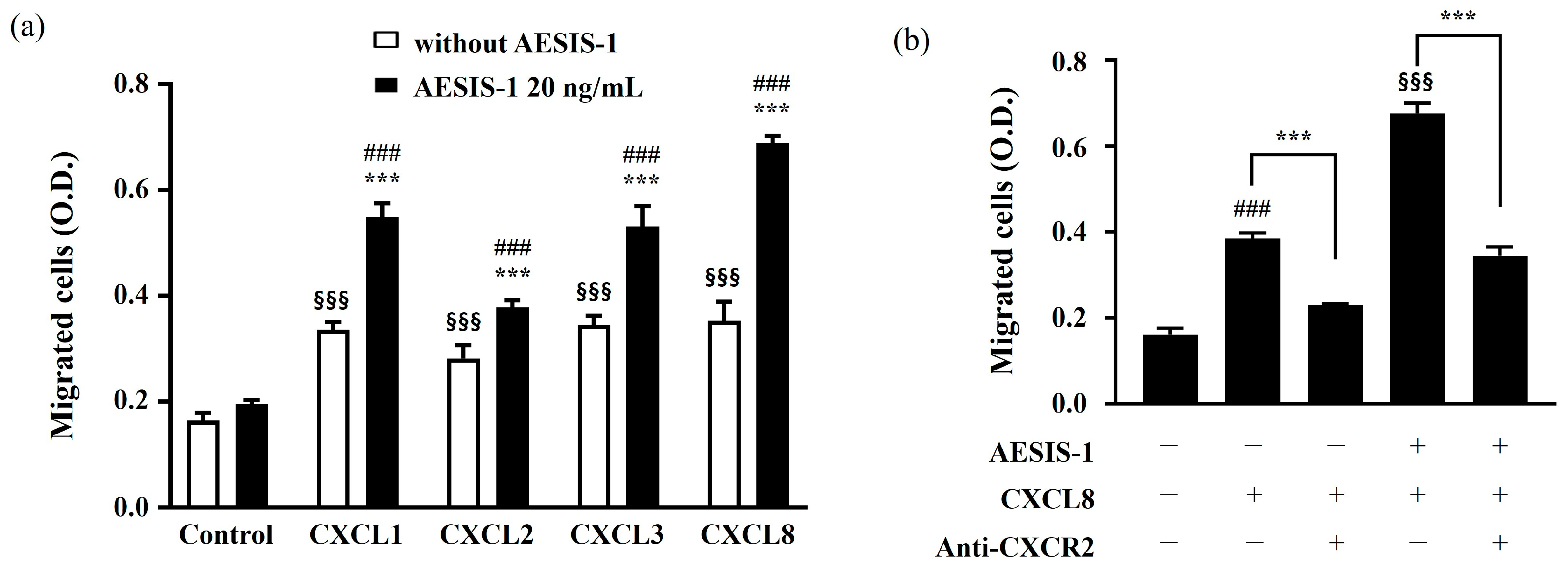
2.3. Enhanced Fibroblast Migration by AESIS-1 Is Mediated by Upregulating CXCR2 Expression
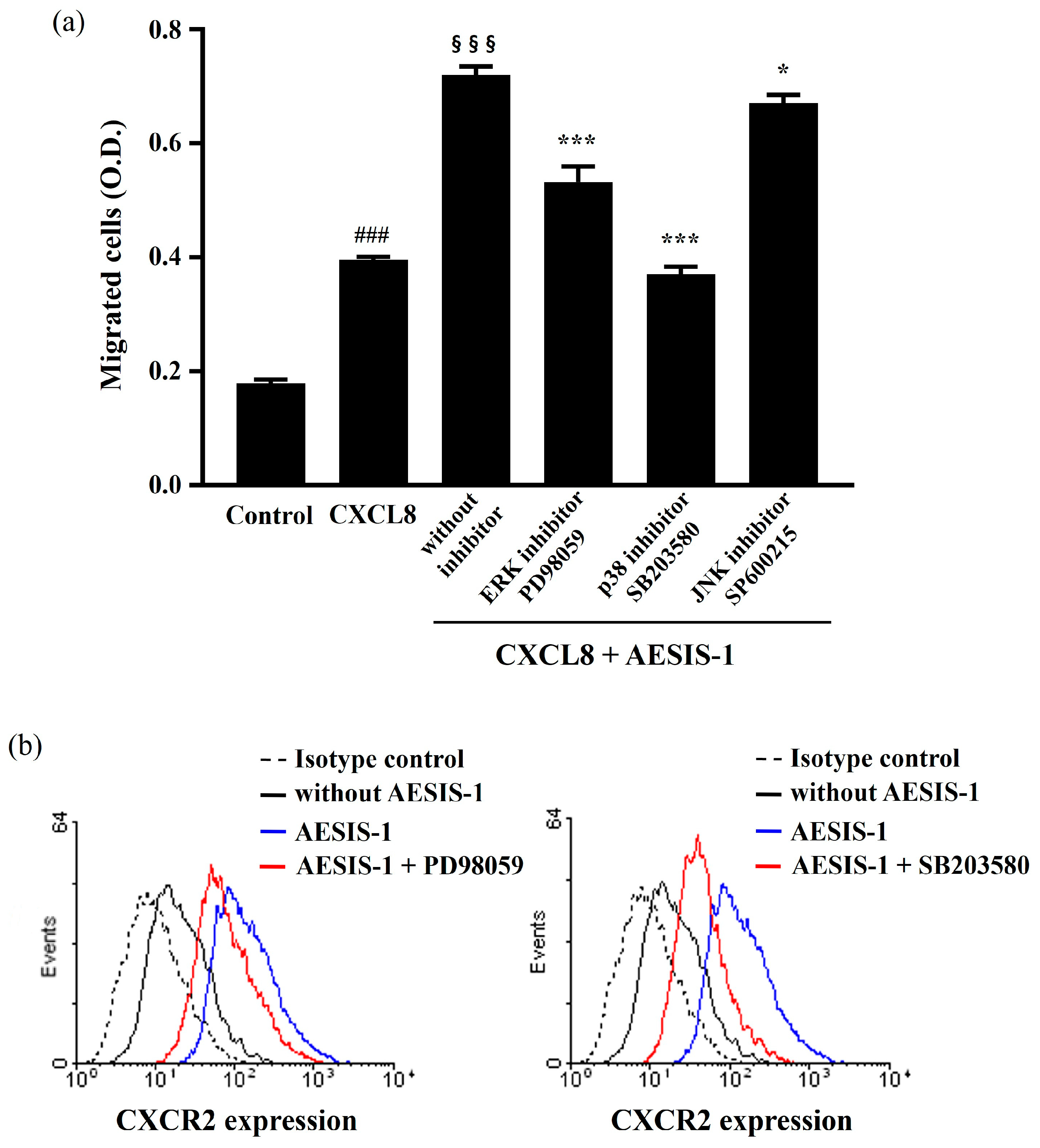
2.4. ERK and p38 MAPK Signaling Cascades Are Involved in AESIS-1-Enhanced Cell Migration and CXCR2 Expression
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Cell Culture and Cell Viability
4.3. Wound-Healing Model in Diabetic Mice
4.4. Transwell Migration Assay
4.5. Real-Time PCR
4.6. Flow Cytometry
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ishie, S.; Ito, H.; Azukizawa, M.; Furu, M.; Ishikawa, M.; Ogino, H.; Hamamoto, Y.; Matsuda, S. Delayed wound healing after forefoot surgery in patients with rheumatoid arthritis. Mod. Rheumatol. 2015, 25, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Jebakumar, A.J.; Udayakumar, P.D.; Crowson, C.S.; Gabriel, S.E.; Matteson, E.L. Occurrence and effect of lower extremity ulcer in rheumatoid arthritis—A population-based Study. J. Rheumatol. 2014, 41, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Vickers, A. Delayed wound healing in patients with rheumatoid arthritis. Nurs. Times 2004, 100, 61–63. [Google Scholar] [PubMed]
- Kirloskar, K.M.; Dekker, P.K.; Kiene, J.; Zhou, S.; Bekeny, J.C.; Rogers, A.; Zolper, E.G.; Fan, K.L.; Evans, K.K.; Benedict, C.D.; et al. The Relationship between Autoimmune Disease and Disease-Modifying Antirheumatic Drugs on Wound Healing. Adv. Wound Care 2022, 11, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, E.; Jamaledin, R.; Familsattarian, F.; Nejaddehbashi, F.; Bagheri, M.; Chehelgerdi, M.; Nazarzadeh Zare, E.; Akhavan, O. Bioactive chitosan/poly(ethyleneoxide)/CuFe2O4 nanofibers for potential wound healing. Environ. Res. 2023, 239, 117448. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Guarino, M.; Hernández-Bule, M.L.; Bacci, S. Cellular and Molecular Processes in Wound Healing. Biomedicines 2023, 11, 2526. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Sun, W.; Wang, W.; Zhang, Y.; Ma, Q. Robust and Wet Adhesive Self-Gelling Powders for Rapid Hemostasis and Efficient Wound Healing. ACS Appl. Mater. Interfaces 2024, 16, 6756–6771. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Talbott, H.E.; Mascharak, S.; Griffin, M.; Wan, D.C.; Longaker, M.T. Wound healing, fibroblast heterogeneity, and fibrosis. Cell Stem Cell 2022, 29, 1161–1180. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Hewitson, T.D. Fibroblast differentiation in wound healing and fibrosis. Int. Rev. Cytol. 2007, 257, 143–179. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, P. Wound healing and the role of fibroblasts. J. Wound Care 2013, 22, 407–408, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Plikus, M.V.; Wang, X.; Sinha, S.; Forte, E.; Thompson, S.M.; Herzog, E.L.; Driskell, R.R.; Rosenthal, N.; Biernaskie, J.; Horsley, V. Fibroblasts: Origins, definitions, and functions in health and disease. Cell 2021, 184, 3852–3872. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Fan, J.; Chen, M.; Guan, S.; Sawcer, D.; Bokoch, G.M.; Woodley, D.T. Mechanism of human dermal fibroblast migration driven by type I collagen and platelet-derived growth factor-BB. Mol. Biol. Cell 2004, 15, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.; Stalfort, J.; Barker, T.H.; Abebayehu, D. The interplay of fibroblasts, the extracellular matrix, and inflammation in scar formation. J. Biol. Chem. 2022, 298, 101530. [Google Scholar] [CrossRef] [PubMed]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef] [PubMed]
- Devalaraja, R.M.; Nanney, L.B.; Du, J.; Qian, Q.; Yu, Y.; Devalaraja, M.N.; Richmond, A. Delayed wound healing in CXCR2 knockout mice. J. Investig. Dermatol. 2000, 115, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Zaja-Milatovic, S.; Richmond, A. CXC chemokines and their receptors: A case for a significant biological role in cutaneous wound healing. Histol. Histopathol. 2008, 23, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Al-Alwan, L.A.; Chang, Y.; Mogas, A.; Halayko, A.J.; Baglole, C.J.; Martin, J.G.; Rousseau, S.; Eidelman, D.H.; Hamid, Q. Differential roles of CXCL2 and CXCL3 and their receptors in regulating normal and asthmatic airway smooth muscle cell migration. J. Immunol. 2013, 191, 2731–2741. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Wong, M.M.; Potter, C.M.; Simpson, R.M.; Karamariti, E.; Zhang, Z.; Zeng, L.; Warren, D.; Hu, Y.; Wang, W.; et al. Vascular Stem/Progenitor Cell Migration Induced by Smooth Muscle Cell-Derived Chemokine (C-C Motif) Ligand 2 and Chemokine (C-X-C motif) Ligand 1 Contributes to Neointima Formation. Stem Cells 2016, 34, 2368–2380. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jacobson, K.; Schaller, M.D. MAP kinases and cell migration. J. Cell Sci. 2004, 117, 4619–4628. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Jinnin, M.; Muchemwa, F.C.; Fukushima, S.; Kogushi-Nishi, H.; Moriya, C.; Igata, T.; Fujisawa, A.; Johno, T.; Ihn, H. Basic fibroblast growth factor stimulates the proliferation of human dermal fibroblasts via the ERK1/2 and JNK pathways. Br. J. Dermatol. 2010, 162, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Dolivo, D.M.; Larson, S.A.; Dominko, T. Crosstalk between mitogen-activated protein kinase inhibitors and transforming growth factor-β signaling results in variable activation of human dermal fibroblasts. Int. J. Mol. Med. 2019, 43, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Nong, N.T.P.; Hsu, J.-L. Bioactive Peptides: An Understanding from Current Screening Methodology. Processes 2022, 10, 1114. [Google Scholar] [CrossRef]
- Da Silva, J.; Leal, E.C.; Carvalho, E. Bioactive Antimicrobial Peptides as Therapeutic Agents for Infected Diabetic Foot Ulcers. Biomolecules 2021, 11, 1894. [Google Scholar] [CrossRef] [PubMed]
- Nasseri, S.; Sharifi, M. Therapeutic Potential of Antimicrobial Peptides for Wound Healing. Int. J. Pept. Res. Ther. 2022, 28, 38. [Google Scholar] [CrossRef]
- Kieslich, C.A.; Alimirzaei, F.; Song, H.; Do, M.; Hall, P. Data-driven prediction of antiviral peptides based on periodicities of amino acid properties. In Computer Aided Chemical Engineering; Türkay, M., Gani, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 50, pp. 2019–2024. [Google Scholar]
- Wang, G.; Vaisman, I.I.; van Hoek, M.L. Machine Learning Prediction of Antimicrobial Peptides. Methods Mol. Biol. 2022, 2405, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Feijoo-Coronel, M.L.; Mendes, B.; Ramírez, D.; Peña-Varas, C.; de Los Monteros-Silva, N.Q.E.; Proaño-Bolaños, C.; de Oliveira, L.C.; Lívio, D.F.; da Silva, J.A.; da Silva, J.; et al. Antibacterial and Antiviral Properties of Chenopodin-Derived Synthetic Peptides. Antibiotics 2024, 13, 78. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.E.; Jeon, S.; Song, J.; Kim, T.S.; Jung, M.K.; Kim, M.S.; Park, S.; Park, S.B.; Park, J.M.; Park, H.J.; et al. The Novel Synthetic Peptide AESIS-1 Exerts a Preventive Effect on Collagen-Induced Arthritis Mouse Model via STAT3 Suppression. Int. J. Mol. Sci. 2020, 21, 378. [Google Scholar] [CrossRef] [PubMed]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Gillitzer, R.; Goebeler, M. Chemokines in cutaneous wound healing. J. Leukoc. Biol. 2001, 69, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Dasari, N.; Jiang, A.; Skochdopole, A.; Chung, J.; Reece, E.M.; Vorstenbosch, J.; Winocour, S. Updates in Diabetic Wound Healing, Inflammation, and Scarring. Semin. Plast. Surg. 2021, 35, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, Z.; Miao, L.; Jiang, X.; Ruan, H.; Xuan, R.; Xu, S. Mechanisms underlying pathological scarring by fibroblasts during wound healing. Int. Wound J. 2023, 20, 2190–2206. [Google Scholar] [CrossRef] [PubMed]
- Gattringer, J.; Gruber, C.W.; Hellinger, R. Peptide modulators of cell migration: Overview, applications and future development. Drug Discov. Today 2023, 28, 103554. [Google Scholar] [CrossRef]
- Bünemann, E.; Hoff, N.P.; Buhren, B.A.; Wiesner, U.; Meller, S.; Bölke, E.; Müller-Homey, A.; Kubitza, R.; Ruzicka, T.; Zlotnik, A.; et al. Chemokine ligand-receptor interactions critically regulate cutaneous wound healing. Eur. J. Med. Res. 2018, 23, 4. [Google Scholar] [CrossRef] [PubMed]
- Addison, C.L.; Daniel, T.O.; Burdick, M.D.; Liu, H.; Ehlert, J.E.; Xue, Y.Y.; Buechi, L.; Walz, A.; Richmond, A.; Strieter, R.M. The CXC chemokine receptor 2, CXCR2, is the putative receptor for ELR+ CXC chemokine-induced angiogenic activity. J. Immunol. 2000, 165, 5269–5277. [Google Scholar] [CrossRef] [PubMed]
- Mehrad, B.; Keane, M.P.; Strieter, R.M. Chemokines as mediators of angiogenesis. Thromb. Haemost. 2007, 97, 755–762. [Google Scholar] [CrossRef] [PubMed]
- IUIS/WHO Subcommittee on Chemokine Nomenclature. Chemokine/chemokine receptor nomenclature. Cytokine 2003, 21, 48–49. [Google Scholar] [CrossRef]
- Hu, H.; Tang, Y.; Pang, L.; Lin, C.; Huang, W.; Wang, D.; Jia, W. Angiogenesis and Full-Thickness Wound Healing Efficiency of a Copper-Doped Borate Bioactive Glass/Poly(lactic-co-glycolic acid) Dressing Loaded with Vitamin E in Vivo and in Vitro. ACS Appl. Mater. Interfaces 2018, 10, 22939–22950. [Google Scholar] [CrossRef] [PubMed]
- Veith, A.P.; Henderson, K.; Spencer, A.; Sligar, A.D.; Baker, A.B. Therapeutic strategies for enhancing angiogenesis in wound healing. Adv. Drug Deliv. Rev. 2019, 146, 97–125. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.D.; Laird, R.E.; Brown, R.D.; Long, C.S. IL-1beta stimulates rat cardiac fibroblast migration via MAP kinase pathways. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1139–H1147. [Google Scholar] [CrossRef]
- Juffermans, N.P.; Dekkers, P.E.; Peppelenbosch, M.P.; Speelman, P.; van Deventer, S.J.; van Der Poll, T. Expression of the chemokine receptors CXCR1 and CXCR2 on granulocytes in human endotoxemia and tuberculosis: Involvement of the p38 mitogen-activated protein kinase pathway. J. Infect. Dis. 2000, 182, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Dobreva, I.; Waeber, G.; James, R.W.; Widmann, C. Interleukin-8 secretion by fibroblasts induced by low density lipoproteins is p38 MAPK-dependent and leads to cell spreading and wound closure. J. Biol. Chem. 2006, 281, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.S.R.; Mousavi, S.; Checco, J.W. Identifying Receptors for Neuropeptides and Peptide Hormones: Challenges and Recent Progress. ACS Chem. Biol. 2021, 16, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target Ther. 2022, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Carretero, M.; Escámez, M.J.; García, M.; Duarte, B.; Holguín, A.; Retamosa, L.; Jorcano, J.L.; Río, M.D.; Larcher, F. In vitro and in vivo wound healing-promoting activities of human cathelicidin LL-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Li, Y.; Tai, Z.; Zhang, Y.; Gao, Y.; Hu, M.; Zhu, Q. Antimicrobial Peptides: The Promising Therapeutics for Cutaneous Wound Healing. Macromol. Biosci. 2021, 21, e2100103. [Google Scholar] [CrossRef] [PubMed]
- John, J.V.; Sharma, N.S.; Tang, G.; Luo, Z.; Su, Y.; Weihs, S.; Shahriar, S.M.S.; Wang, G.; McCarthy, A.; Dyke, J.; et al. Nanofiber Aerogels with Precision Macrochannels and LL-37-Mimic Peptides Synergistically Promote Diabetic Wound Healing. Adv. Funct. Mater. 2023, 33, 2206936. [Google Scholar] [CrossRef] [PubMed]
- Fathi, F.; Ghobeh, M.; Tabarzad, M. Anti-Microbial Peptides: Strategies of Design and Development and Their Promising Wound-Healing Activities. Mol. Biol. Rep. 2022, 49, 9001–9012. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Du, J.; Xue, W.; Shao, K.; Jiang, X.; Peng, W.; Li, W.; Huang, S. Cathelicidin LL-37 promotes wound healing in diabetic mice by regulating TFEB-dependent autophagy. Peptides 2024, 175, 171183. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, L.; Zhou, J.; Qin, A.; Chen, Z. The protective effect of formononetin on cognitive impairment in streptozotocin (STZ)-induced diabetic mice. Biomed. Pharmacother. 2018, 106, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Hozzein, W.N.; Badr, G.; Al Ghamdi, A.A.; Sayed, A.; Al-Waili, N.S.; Garraud, O. Topical application of propolis enhances cutaneous wound healing by promoting TGF-beta/Smad-mediated collagen production in a streptozotocin-induced type I diabetic mouse model. Cell. Physiol. Biochem. 2015, 37, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, M.S.; Jung, S.J.; Kim, D.; Park, H.J.; Cho, D. ERK activating peptide, AES16-2M promotes wound healing through accelerating migration of keratinocytes. Sci. Rep. 2018, 8, 14398. [Google Scholar] [CrossRef]
- Yang, E.J.; Bang, S.I. Effects of conditioned medium from LL-37 treated adipose stem cells on human fibroblast migration. Exp. Ther. Med. 2017, 14, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Choi, H.; Seon, M.; Cho, D.; Bang, S.I. LL-37 stimulates the functions of adipose-derived stromal/stem cells via early growth response 1 and the MAPK pathway. Stem Cell Res. Ther. 2016, 7, 58. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Title 2 | Title 3 |
|---|---|---|
| GAPDH | 5′-ATCACCATCTTCCAGGAGCGA-3′ | 5′-TTCTCCATGGTGGTGAAGACG-3′ |
| CCR1 | 5′-ACCATAGGAGGCCAACCCAAAATA-3′ | 5′-TCCATGCTGTGCCAAGAGTCA-3′ |
| CCR2 | 5′-CTCTCCCATTGTGGGCTCACTCTG-3′ | 5′-GCAAACACAGCATGGACAATAGCC-3′ |
| CCR3 | 5′-TTTGTCATCATGGCGGTGTTTTTC-3′ | 5′-GGTTCATGCAGCAGTGGGAGTAG-3′ |
| CCR4 | 5′-GAGAAGAAGAACAAGGCGGTGAAGA-3′ | 5′-GGATTAAGGCAGCAGTGAACAAAAG-3′ |
| CCR6 | 5′-CTGCCTGAACCCTGTGCTCTACG-3′ | 5′-TTATCTGCGGTCTCACTGGTCTGC-3′ |
| CCR7 | 5′-GCCGAGACCACCACCACCTT-3′ | 5′-AGTCATTGCATCTGCTCCCTATCC-3′ |
| CCR9 | 5′-TATACAGCCAAATCAAGGAGGAATC-3′ | 5′-CATGACCACGAAGGGAAGGAAG-3′ |
| CCR10 | 5′-GGGCTGGAGTCTGGGAAGTGC-3′ | 5′-ACGATGACGGAGACCAAGTGTGC-3′ |
| CXCR1 | 5′-CTGAGCCCCAAGTGGAACGAGACA-3′ | 5′-GCACGGAACAGAAGCTTTATTAGGA-3′ |
| CXCR2 | 5′-CAATGAATGAATGAATGGCTAAG-3′ | 5′-AAAGTTTTCAAGGTTCGTCCGTGTT-3′ |
| CXCR4 | 5′-AATCTTCCTGCCCACCATCTACTCC-3′ | 5′-GCGGTCACAGATATATCTGTCATCTGCC-3′ |
| CXCR5 | 5′-TCCCCTCCTCACTCCCTTCCCATAA-3′ | 5′-CCTGCGGTTCCATCTGAGTGACATC-3′ |
| CXCR6 | 5′-TTGTTTATAGCTTGCGCATTCTCAT-3′ | 5′-ATCCCCCTTGGTTTCAGCATTCTT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.B.; Yang, Y.; Bang, S.I.; Kim, T.S.; Cho, D. AESIS-1, a Rheumatoid Arthritis Therapeutic Peptide, Accelerates Wound Healing by Promoting Fibroblast Migration in a CXCR2-Dependent Manner. Int. J. Mol. Sci. 2024, 25, 3937. https://doi.org/10.3390/ijms25073937
Park SB, Yang Y, Bang SI, Kim TS, Cho D. AESIS-1, a Rheumatoid Arthritis Therapeutic Peptide, Accelerates Wound Healing by Promoting Fibroblast Migration in a CXCR2-Dependent Manner. International Journal of Molecular Sciences. 2024; 25(7):3937. https://doi.org/10.3390/ijms25073937
Chicago/Turabian StylePark, Seung Beom, Yoolhee Yang, Sa Ik Bang, Tae Sung Kim, and Daeho Cho. 2024. "AESIS-1, a Rheumatoid Arthritis Therapeutic Peptide, Accelerates Wound Healing by Promoting Fibroblast Migration in a CXCR2-Dependent Manner" International Journal of Molecular Sciences 25, no. 7: 3937. https://doi.org/10.3390/ijms25073937