
Serum Induces the Subunit-Specific Activation of NF-κB in Proliferating Human Cardiac Stem Cells

 ,
,
Abstract
:1. Introduction
2. Results
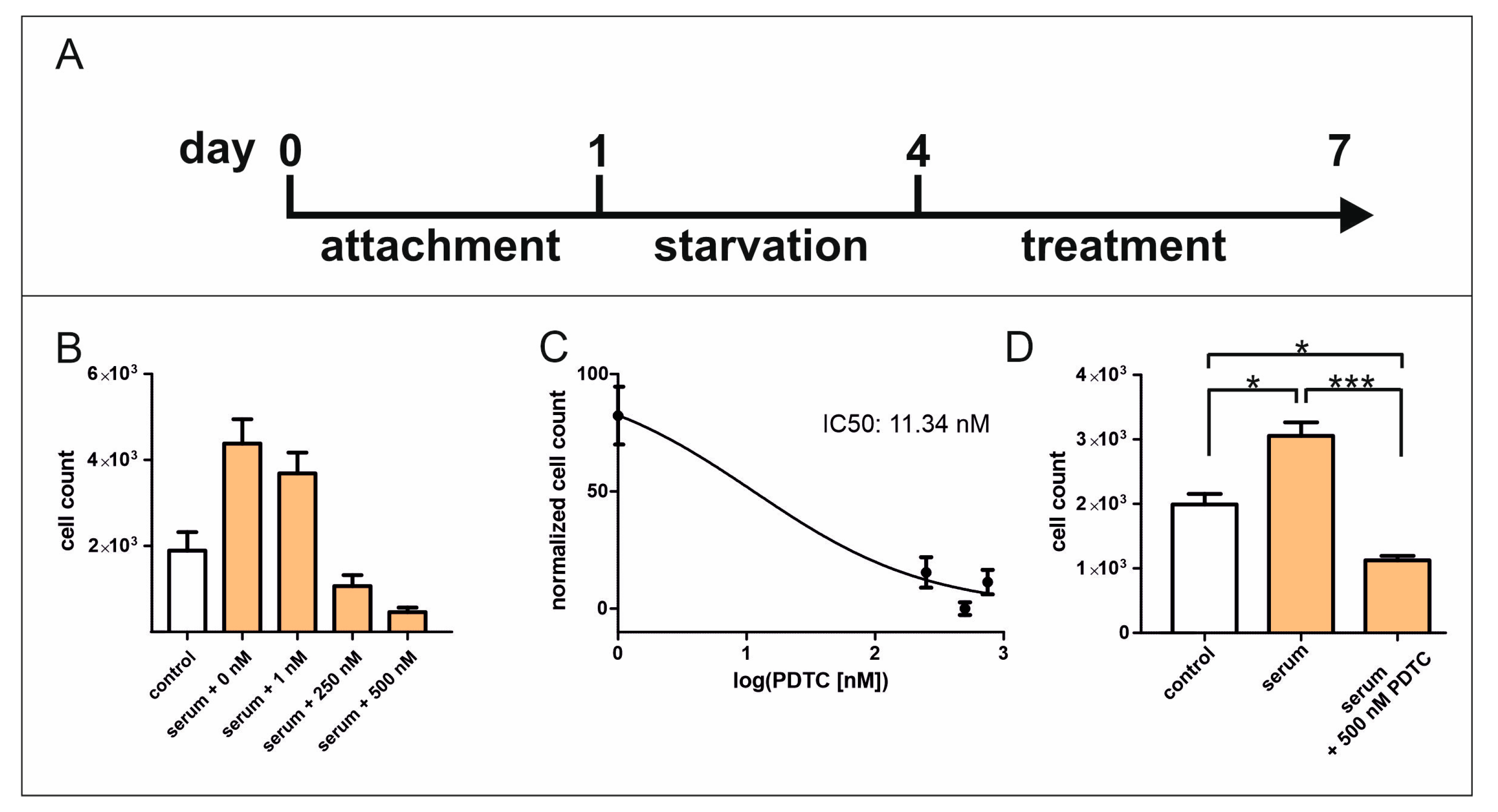
2.1. Inhibition of NF-κB Signalling Reduces Serum-Induced Cell Proliferation of hCSCs
2.2. Human Blood Serum Induces NF-κB Subunit Translocation in hCSCs
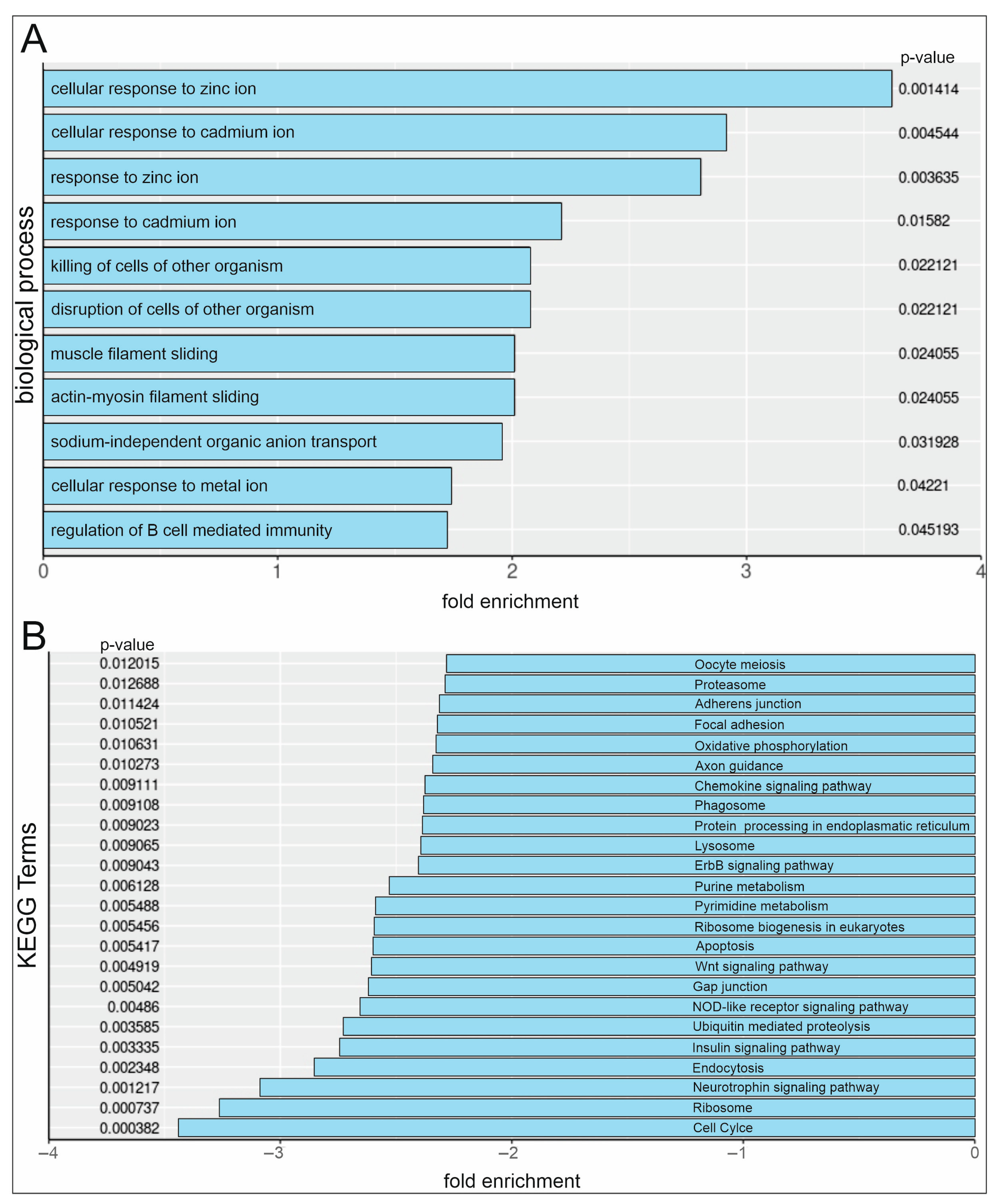
2.3. RNA-Sequencing Reveals Upregulation of Cellular Ion Response GO-Terms and Downregulation of Cell Cycle KEGG Pathways in Serum and PDTC-Treated hCSCs
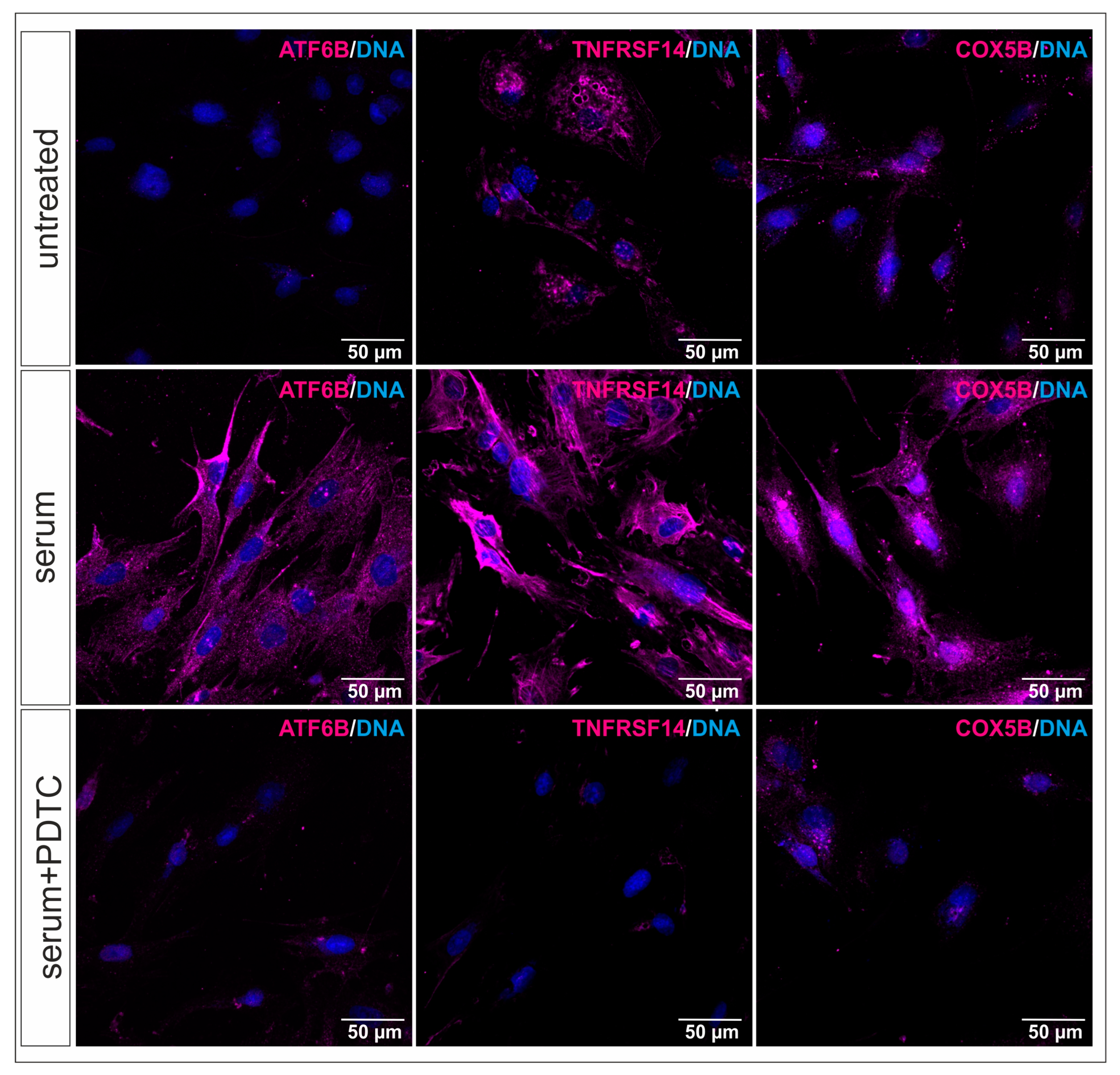
2.4. RNA-Sequencing Reveals New NF-κB Target Genes in Serum-Treated hCSCs
3. Discussion
4. Materials and Methods
4.1. Isolation and Cultivation of Human Cardiac Stem Cells
4.2. Collection of Human Blood Serum Samples
4.3. Proliferation Assay
4.4. Immunocytochemical Staining
4.5. RNA-Sequencing
4.6. Promotor Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, M.A.; Murry, C.E. Heart Regeneration. Nature 2011, 473, 326–335. [Google Scholar] [CrossRef]
- Aguilar-Sanchez, C.; Michael, M.; Pennings, S. Cardiac Stem Cells in the Postnatal Heart: Lessons from Development. Stem Cells Int. 2018, 2018, 1247857. [Google Scholar] [CrossRef]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult Cardiac Stem Cells Are Multipotent and Support Myocardial Regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Bearzi, C.; Rota, M.; Hosoda, T.; Tillmanns, J.; Nascimbene, A.; De Angelis, A.; Yasuzawa-Amano, S.; Trofimova, I.; Siggins, R.W.; LeCapitaine, N.; et al. Human Cardiac Stem Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar] [CrossRef]
- Smits, A.M.; van Vliet, P.; Metz, C.H.; Korfage, T.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.-J. Human Cardiomyocyte Progenitor Cells Differentiate into Functional Mature Cardiomyocytes: An In Vitro Model for Studying Human Cardiac Physiology and Pathophysiology. Nat. Protoc. 2009, 4, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Höving, A.L.; Schmidt, K.E.; Merten, M.; Hamidi, J.; Rott, A.-K.; Faust, I.; Greiner, J.F.W.; Gummert, J.; Kaltschmidt, B.; Kaltschmidt, C.; et al. Blood Serum Stimulates P38-Mediated Proliferation and Changes in Global Gene Expression of Adult Human Cardiac Stem Cells. Cells 2020, 9, 1472. [Google Scholar] [CrossRef]
- Höving, A.L.; Schmidt, K.E.; Kaltschmidt, B.; Kaltschmidt, C.; Knabbe, C. The Role of Blood-Derived Factors in Protection and Regeneration of Aged Tissues. Int. J. Mol. Sci. 2022, 23, 9626. [Google Scholar] [CrossRef]
- Höving, A.L.; Sielemann, K.; Greiner, J.F.W.; Kaltschmidt, B.; Knabbe, C.; Kaltschmidt, C. Transcriptome Analysis Reveals High Similarities between Adult Human Cardiac Stem Cells and Neural Crest-Derived Stem Cells. Biology 2020, 9, 435. [Google Scholar] [CrossRef]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young Blood Reverses Age-Related Impairments in Cognitive Function and Synaptic Plasticity in Mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef]
- Conboy, I.M.; Conboy, M.J.; Wagers, A.J.; Girma, E.R.; Weissman, I.L.; Rando, T.A. Rejuvenation of Aged Progenitor Cells by Exposure to a Young Systemic Environment. Nature 2005, 433, 760–764. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Schafer, M.J.; Noren Hooten, N.; Atkinson, E.J.; Evans, M.K.; Baker, D.J.; Quarles, E.K.; Robbins, P.D.; Ladiges, W.C.; LeBrasseur, N.K.; et al. Circulating Levels of Monocyte Chemoattractant Protein-1 as a Potential Measure of Biological Age in Mice and Frailty in Humans. Aging Cell 2018, 17, e12706. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, M.; Lenz, V.; Knop, D.; Klump, H.; Alt, M.; Aufderhorst, U.W.; Schipper, L.; Schwarzkopf, S.; Meller, L.; Steckel, N.; et al. Convalescent Plasma Treatment of Critically Ill Intensive Care COVID-19 Patients. Transfusion 2021, 61, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, A.; Cassar, J. Plasma-Derived Medicines: Access and Usage Issues. Blood Transfus. 2012, 10, 273–278. [Google Scholar] [CrossRef]
- Schmidt, K.E.; Höving, A.L.; Kiani Zahrani, S.; Trevlopoulou, K.; Kaltschmidt, B.; Knabbe, C.; Kaltschmidt, C. Serum-Induced Proliferation of Human Cardiac Stem Cells Is Modulated via TGFβRI/II and SMAD2/3. Int. J. Mol. Sci. 2024, 25, 959. [Google Scholar] [CrossRef] [PubMed]
- Höving, A.L.; Schmitz, J.; Schmidt, K.E.; Greiner, J.F.W.; Knabbe, C.; Kaltschmidt, B.; Grünberger, A.; Kaltschmidt, C. Human Blood Serum Induces P38-MAPK- and Hsp27-Dependent Migration Dynamics of Adult Human Cardiac Stem Cells: Single-Cell Analysis via a Microfluidic-Based Cultivation Platform. Biology 2021, 10, 708. [Google Scholar] [CrossRef]
- Kaltschmidt, C.; Greiner, J.F.W.; Kaltschmidt, B. The Transcription Factor NF-κB in Stem Cells and Development. Cells 2021, 10, 2042. [Google Scholar] [CrossRef] [PubMed]
- Schreck, R.; Meier, B.; Männel, D.N.; Dröge, W.; Baeuerle, P.A. Dithiocarbamates as Potent Inhibitors of Nuclear Factor Kappa B Activation in Intact Cells. J. Exp. Med. 1992, 175, 1181–1194. [Google Scholar] [CrossRef]
- Abete, P.; Napoli, C.; Santoro, G.; Ferrara, N.; Tritto, I.; Chiariello, M.; Rengo, F.; Ambrosio, G. Age-Related Decrease in Cardiac Tolerance to Oxidative Stress. J. Mol. Cell. Cardiol. 1999, 31, 227–236. [Google Scholar] [CrossRef]
- Abete, P.; Della-Morte, D.; Gargiulo, G.; Basile, C.; Langellotto, A.; Galizia, G.; Testa, G.; Canonico, V.; Bonaduce, D.; Cacciatore, F. Cognitive Impairment and Cardiovascular Diseases in the Elderly. A Heart–Brain Continuum Hypothesis. Ageing Res. Rev. 2014, 18, 41–52. [Google Scholar] [CrossRef]
- Picerno, A.; Stasi, A.; Franzin, R.; Curci, C.; di Bari, I.; Gesualdo, L.; Sallustio, F. Why Stem/Progenitor Cells Lose Their Regenerative Potential. World J. Stem Cells 2021, 13, 1714–1732. [Google Scholar] [CrossRef]
- Fossett, E.; Khan, W.S.; Pastides, P.; Adesida, A.B. The Effects of Ageing on Proliferation Potential, Differentiation Potential and Cell Surface Characterisation of Human Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2012, 7, 282–286. [Google Scholar] [CrossRef]
- Fuentes, T.; Kearns-Jonker, M. Endogenous Cardiac Stem Cells for the Treatment of Heart Failure. Stem Cells Cloning 2013, 6, 1–12. [Google Scholar] [CrossRef]
- Freudlsperger, C.; Bian, Y.; Contag, S.; Burnett, J.; Coupar, J.; Yang, X.; Chen, Z.; Van Waes, C. TGF-β and NF-κB Signal Pathway Cross-Talk Is Mediated through TAK1 and SMAD7 in a Subset of Head and Neck Cancers. Oncogene 2013, 32, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Widera, D.; Mikenberg, I.; Elvers, M.; Kaltschmidt, C.; Kaltschmidt, B. Tumor Necrosis Factor α Triggers Proliferation of Adult Neural Stem Cells via IKK/NF-κB Signaling. BMC Neurosci. 2006, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Carrero, R.; Cerrada, I.; Lledó, E.; Dopazo, J.; García-García, F.; Rubio, M.-P.; Trigueros, C.; Dorronsoro, A.; Ruiz-Sauri, A.; Montero, J.A.; et al. IL1β Induces Mesenchymal Stem Cells Migration and Leucocyte Chemotaxis Through NF-κB. Stem Cell Rev. Rep. 2012, 8, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Pilastro, M.R.; DiMarco, R.; Scifo, C.; Renis, M.; Mazzarino, M.C.; Messina, A. Cell Death in Human Acute Myelogenous Leukemic Cells Induced by Pyrrolidinedithiocarbamate. Apoptosis 2003, 8, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Erl, W.; Weber, C.; Hansson, G.K. Pyrrolidine Dithiocarbamate-Induced Apoptosis Depends on Cell Type, Density, and the Presence of Cu2+ and Zn2+. Am. J. Physiol.-Cell Physiol. 2000, 278, C1116–C1125. [Google Scholar] [CrossRef] [PubMed]
- Bessho, R.; Matsubara, K.; Kubota, M.; Kuwakado, K.; Hirota, H.; Wakazono, Y.; Lin, Y.W.; Okuda, A.; Kawai, M.; Nishikomori, R.; et al. Pyrrolidine Dithiocarbamate, a Potent Inhibitor of Nuclear Factor κB (NF-κB) Activation, Prevents Apoptosis in Human Promyelocytic Leukemia HL-60 Cells and Thymocytes. Biochem. Pharmacol. 1994, 48, 1883–1889. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.; Chae, S.; Park, N.; Bang, B.; Cho, S.; Kim, J.; Kim, H.; Kim, H.; Lee, Z.; Kim, H. Pyrrolidine Dithiocarbamate Inhibits Serum-Induced NF-κB Activation and Induces Apoptosis in ROS 17/2.8 Osteoblasts. Int. Immunopharmacol. 2001, 1, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Monks, N.R.; Biswas, D.K.; Pardee, A.B. Blocking Anti-Apoptosis as a Strategy for Cancer Chemotherapy: NF-κB as a Target. J. Cell. Biochem. 2004, 92, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Baichwal, V.R.; Baeuerle, P.A. Apoptosis: Activate NF-κB or Die? Curr. Biol. 1997, 7, R94–R96. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-Y.; Liu, D.-J.; Liu, M.-X. The Protective Effect of NF-κB Signaling Pathway Inhibitor PDTC on Mice with Chronic Atrophic Gastritis. Scand. J. Gastroenterol. 2021, 56, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Tahata, S.; Yuan, B.; Kikuchi, H.; Takagi, N.; Hirano, T.; Toyoda, H. Cytotoxic Effects of Pyrrolidine Dithiocarbamate in Small-Cell Lung Cancer Cells, Alone and in Combination with Cisplatin. Int. J. Oncol. 2014, 45, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Pyrrolidine Dithiocarbamate Inhibits NF-κB Activation and Enhances TNF-Induced Apoptosis in Human Breast Cancer Cells. Available online: https://www.springermedizin.de/pyrrolidine-dithiocarbamate-inhibits-nf-%CE%BAb-activation-and-enhanc/9028714 (accessed on 2 November 2023).
- Cuzzocrea, S.; Chatterjee, P.K.; Mazzon, E.; Dugo, L.; Serraino, I.; Britti, D.; Mazzullo, G.; Caputi, A.P.; Thiemermann, C. Pyrrolidine Dithiocarbamate Attenuates the Development of Acute and Chronic Inflammation. Br. J. Pharmacol. 2002, 135, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.F.; Ye, X.; Malik, A.B. Inhibition of NF-κB Activation by Pyrrolidine Dithiocarbamate Prevents In Vivo Expression of Proinflammatory Genes. Circulation 1999, 100, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Chabicovsky, M.; Prieschl-Grassauer, E.; Seipelt, J.; Muster, T.; Szolar, O.H.J.; Hebar, A.; Doblhoff-Dier, O. Pre-Clinical Safety Evaluation of Pyrrolidine Dithiocarbamate. Basic Clin. Pharmacol. Toxicol. 2010, 107, 758–767. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Greiner, J.F.W.; Kadhim, H.M.; Kaltschmidt, C. Subunit-Specific Role of NF-κB in Cancer. Biomedicines 2018, 6, 44. [Google Scholar] [CrossRef]
- Induced Neural Stem Cells Regulate Microglial Activation through Akt-Mediated Upregulation of CXCR4 and Crry in a Mouse Model of Closed Head Injury. Available online: https://www.researchsquare.com (accessed on 9 November 2023).
- Kumar, A.; Kumar, A.; Michael, P.; Brabant, D.; Parissenti, A.M.; Ramana, C.V.; Xu, X.; Parrillo, J.E. Human Serum from Patients with Septic Shock Activates Transcription Factors STAT1, IRF1, and NF-κB and Induces Apoptosis in Human Cardiac Myocytes*. J. Biol. Chem. 2005, 280, 42619–42626. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Moniuszko-Jakoniuk, J. Interactions between Cadmium and Zinc in the Organism. Food Chem. Toxicol. 2001, 39, 967–980. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.H.; Lee, J.; Ahn, Y.S. Zinc-Induced NF-κB Inhibition Can Be Modulated by Changes in the Intracellular Metallothionein Level. Toxicol. Appl. Pharmacol. 2003, 190, 189–196. [Google Scholar] [CrossRef]
- Lanke, K.; Krenn, B.M.; Melchers, W.J.G.; Seipelt, J.; van Kuppeveld, F.J.M. PDTC Inhibits Picornavirus Polyprotein Processing and RNA Replication by Transporting Zinc Ions into Cells. J. Gen. Virol. 2007, 88, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Nobel, C.S.I.; Kimland, M.; Lind, B.; Orrenius, S.; Slater, A.F.G. Dithiocarbamates Induce Apoptosis in Thymocytes by Raising the Intracellular Level of Redox-Active Copper (∗). J. Biol. Chem. 1995, 270, 26202–26208. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.-B.; Wang, T.-X.; Liu, Z.-W.; Yan, J.-Y.; Zhang, K. Zinc Finger Protein A20 Regulates the Development and Progression of Osteoarthritis by Affecting the Activity of NF-κB P65. Immunopharmacol. Immunotoxicol. 2021, 43, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M. Zinc Fingers and Other Metal-Binding Domains. Elements for Interactions between Macromolecules. J. Biol. Chem. 1990, 265, 6513–6516. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kim, J.H.; Xu, J.; Hsu, C.Y.; Ahn, Y.S. Pyrrolidine Dithiocarbamate Induces Bovine Cerebral Endothelial Cell Death by Increasing the Intracellular Zinc Level. J. Neurochem. 1999, 72, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Seve, M.; Chimienti, F.; Favier, A. Role of intracellular zinc in programmed cell death. Pathol. Biol. 2002, 50, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.B.; Costello, L.C. The Important Role of the Apoptotic Effects of Zinc in the Development of Cancers. J. Cell. Biochem. 2009, 106, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, Y.; Yang, Y.; Qiu, Y.; Wang, Z.; Li, X.; Zhang, W. Emerging Roles of Activating Transcription Factor (ATF) Family Members in Tumourigenesis and Immunity: Implications in Cancer Immunotherapy. Genes Dis. 2022, 9, 981–999. [Google Scholar] [CrossRef]
- Bakin, A.V.; Stourman, N.V.; Sekhar, K.R.; Rinehart, C.; Yan, X.; Meredith, M.J.; Arteaga, C.L.; Freeman, M.L. Smad3–ATF3 Signaling Mediates TGF-β Suppression of Genes Encoding Phase II Detoxifying Proteins. Free Radic. Biol. Med. 2005, 38, 375–387. [Google Scholar] [CrossRef]
- Kim, E.-S.; Sohn, Y.-W.; Moon, A. TGF-β-Induced Transcriptional Activation of MMP-2 Is Mediated by Activating Transcription Factor (ATF)2 in Human Breast Epithelial Cells. Cancer Lett. 2007, 252, 147–156. [Google Scholar] [CrossRef]
- Mallano, T.; Palumbo-Zerr, K.; Zerr, P.; Ramming, A.; Zeller, B.; Beyer, C.; Dees, C.; Huang, J.; Hai, T.; Distler, O.; et al. Activating Transcription Factor 3 Regulates Canonical TGFβ Signalling in Systemic Sclerosis. Ann. Rheum. Dis. 2016, 75, 586–592. [Google Scholar] [CrossRef]
- Potchinsky, M.B.; Weston, W.M.; Lloyd, M.R.; Greene, R.M. TGF-β Signaling in Murine Embryonic Palate Cells Involves Phosphorylation of the CREB Transcription Factor. Exp. Cell Res. 1997, 231, 96–103. [Google Scholar] [CrossRef]
- Koga, Y.; Hisada, T.; Ishizuka, T.; Utsugi, M.; Ono, A.; Yatomi, M.; Kamide, Y.; Aoki-Saito, H.; Tsurumaki, H.; Dobashi, K.; et al. CREB Regulates TNF-α-Induced GM-CSF Secretion via P38 MAPK in Human Lung Fibroblasts. Allergol. Int. 2016, 65, 406–413. [Google Scholar] [CrossRef]
- Gustin, J.A.; Pincheira, R.; Mayo, L.D.; Ozes, O.N.; Kessler, K.M.; Baerwald, M.R.; Korgaonkar, C.K.; Donner, D.B. Tumor Necrosis Factor Activates CRE-Binding Protein through a P38 MAPK/MSK1 Signaling Pathway in Endothelial Cells. Am. J. Physiol.-Cell Physiol. 2004, 286, C547–C555. [Google Scholar] [CrossRef]
- Iezaki, T.; Ozaki, K.; Fukasawa, K.; Inoue, M.; Kitajima, S.; Muneta, T.; Takeda, S.; Fujita, H.; Onishi, Y.; Horie, T.; et al. ATF3 Deficiency in Chondrocytes Alleviates Osteoarthritis Development. J. Pathol. 2016, 239, 426–437. [Google Scholar] [CrossRef]
- Du, W.; Thanos, D.; Maniatis, T. Mechanisms of Transcriptional Synergism between Distinct Virus-Inducible Enhancer Elements. Cell 1993, 74, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.; Emi, M.; Tanabe, K.; Murakami, S. Role of the Unfolded Protein Response in Cell Death. Apoptosis 2006, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Y.; Geng, X.; Li, H.; Wang, X.; Zhang, Y.; Zhao, H. Zinc Attenuates Arsenic Overdose-Induced Brain Damage via PERK/ATF6 and TLR/MyD88/NF-κB Pathways. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2024, 276, 109806. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-P.; Sun, H.-F.; Jiang, H.-L.; Li, L.-D.; Hu, X.; Xu, X.-E.; Jin, W. Loss of COX5B Inhibits Proliferation and Promotes Senescence via Mitochondrial Dysfunction in Breast Cancer. Oncotarget 2015, 6, 43363. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-K.; Noh, E.-K.; Gwon, G.-D.; Kim, J.Y.; Jo, J.-C.; Choi, Y.; Koh, S.; Baek, J.H.; Min, Y.J.; Kim, H. LIGHT (TNFSF14) Increases the Survival and Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells. PLoS ONE 2016, 11, e0166589. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-D.; Lu, M.-Y. Increased Expression of TNFRSF14 Indicates Good Prognosis and Inhibits Bladder Cancer Proliferation by Promoting Apoptosis. Mol. Med. Rep. 2018, 18, 3403–3410. [Google Scholar] [CrossRef] [PubMed]
- Slotta, C.; Schlüter, T.; Ruiz-Perera, L.M.; Kadhim, H.M.; Tertel, T.; Henkel, E.; Hübner, W.; Greiner, J.F.W.; Huser, T.; Kaltschmidt, B.; et al. CRISPR/Cas9-Mediated Knockout of c-REL in HeLa Cells Results in Profound Defects of the Cell Cycle. PLoS ONE 2017, 12, e0182373. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Fornes, O.; Stigliani, A.; Gheorghe, M.; Castro-Mondragon, J.A.; van der Lee, R.; Bessy, A.; Chèneby, J.; Kulkarni, S.R.; Tan, G.; et al. JASPAR 2018: Update of the Open-Access Database of Transcription Factor Binding Profiles and Its Web Framework. Nucleic Acids Res. 2018, 46, D260–D266. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promotor | Name | Score | Start | End | +/− | Predicted Sequence |
|---|---|---|---|---|---|---|
| ATF6B_1 | MA0101.1.REL | 11.31 | 1342 | 1351 | + | TGGGGTTTCC |
| MA0105.3.NFKB1 | 12.59 | 1342 | 1352 | + | TGGGGTTTCCT | |
| MA0107.1.RELA | 11.98 | 1342 | 1351 | + | TGGGGTTTCC | |
| COX5B_1 | MA0105.3.NFKB1 | 10.71 | 1499 | 1509 | − | GGGCTTTTCCT |
| MA0101.1.REL | 9.85 | 1500 | 1509 | − | GGGCTTTTCC | |
| TNFRSF14_1 | MA0105.2.NFKB1 | 13.66 | 68 | 78 | − | GGGGGTCTCCC |
| MA0105.3.NFKB1 | 11.37 | 259 | 269 | + | TGGGGTTCCCT | |
| TNFRSF14_2 | MA0105.1.NFKB1 | 12.80 | 948 | 957 | + | GGGACATTCC |
| MA0101.1.REL | 11.13 | 948 | 957 | + | GGGACATTCC | |
| MA0105.2.NFKB1 | 13.67 | 2224 | 2234 | − | GGGGAGCCCCC | |
| MA0105.3.NFKB1 | 12.15 | 1984 | 1994 | + | GGGCTTTTCCC | |
| MA0105.3.NFKB1 | 11.93 | 1817 | 1827 | − | TGGGCTTCCCT | |
| MA0101.1.REL | 10.33 | 1070 | 1079 | + | GGGGCTTTTC | |
| MA0778.1.NFKB2 | 13.93 | 2224 | 2236 | + | GGGGGCTCCCCCT | |
| MA0105.2.NFKB1 | 13.10 | 2225 | 2235 | + | GGGGCTCCCCC | |
| MA0105.3.NFKB1 | 11.22 | 1070 | 1080 | + | GGGGCTTTTCC | |
| MA0778.1.NFKB2 | 13.45 | 2224 | 2236 | − | AGGGGGAGCCCCC | |
| TNFRSF14_3 | MA0105.1.NFKB1 | 13.53 | 1613 | 1622 | − | GGGGCTTCCC |
| MA0105.2.NFKB1 | 14.66 | 2265 | 2275 | + | GGGGAGTCCCC | |
| MA0105.3.NFKB1 | 13.57 | 1612 | 1622 | - | GGGGCTTCCCA | |
| MA0105.2.NFKB1 | 13.66 | 986 | 996 | − | GGGGGTCTCCC | |
| MA0105.2.NFKB1 | 13.13 | 2265 | 2275 | − | GGGGACTCCCC | |
| MA0105.1.NFKB1 | 11.38 | 2265 | 2274 | + | GGGGAGTCCC | |
| MA0105.1.NFKB1 | 11.11 | 2265 | 2274 | − | GGGACTCCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, K.E.; Höving, A.L.; Nowak, K.; an Mey, N.; Kiani Zahrani, S.; Nemeita, B.; Riedel, L.; Majewski, A.; Kaltschmidt, B.; Knabbe, C.; et al. Serum Induces the Subunit-Specific Activation of NF-κB in Proliferating Human Cardiac Stem Cells. Int. J. Mol. Sci. 2024, 25, 3593. https://doi.org/10.3390/ijms25073593
Schmidt KE, Höving AL, Nowak K, an Mey N, Kiani Zahrani S, Nemeita B, Riedel L, Majewski A, Kaltschmidt B, Knabbe C, et al. Serum Induces the Subunit-Specific Activation of NF-κB in Proliferating Human Cardiac Stem Cells. International Journal of Molecular Sciences. 2024; 25(7):3593. https://doi.org/10.3390/ijms25073593
Chicago/Turabian StyleSchmidt, Kazuko E., Anna L. Höving, Katja Nowak, Nike an Mey, Sina Kiani Zahrani, Britta Nemeita, Lena Riedel, Agnes Majewski, Barbara Kaltschmidt, Cornelius Knabbe, and et al. 2024. "Serum Induces the Subunit-Specific Activation of NF-κB in Proliferating Human Cardiac Stem Cells" International Journal of Molecular Sciences 25, no. 7: 3593. https://doi.org/10.3390/ijms25073593