
Myconoside and Calceolarioside E Restrain UV-Induced Skin Photoaging by Activating NRF2-Mediated Defense Mechanisms

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
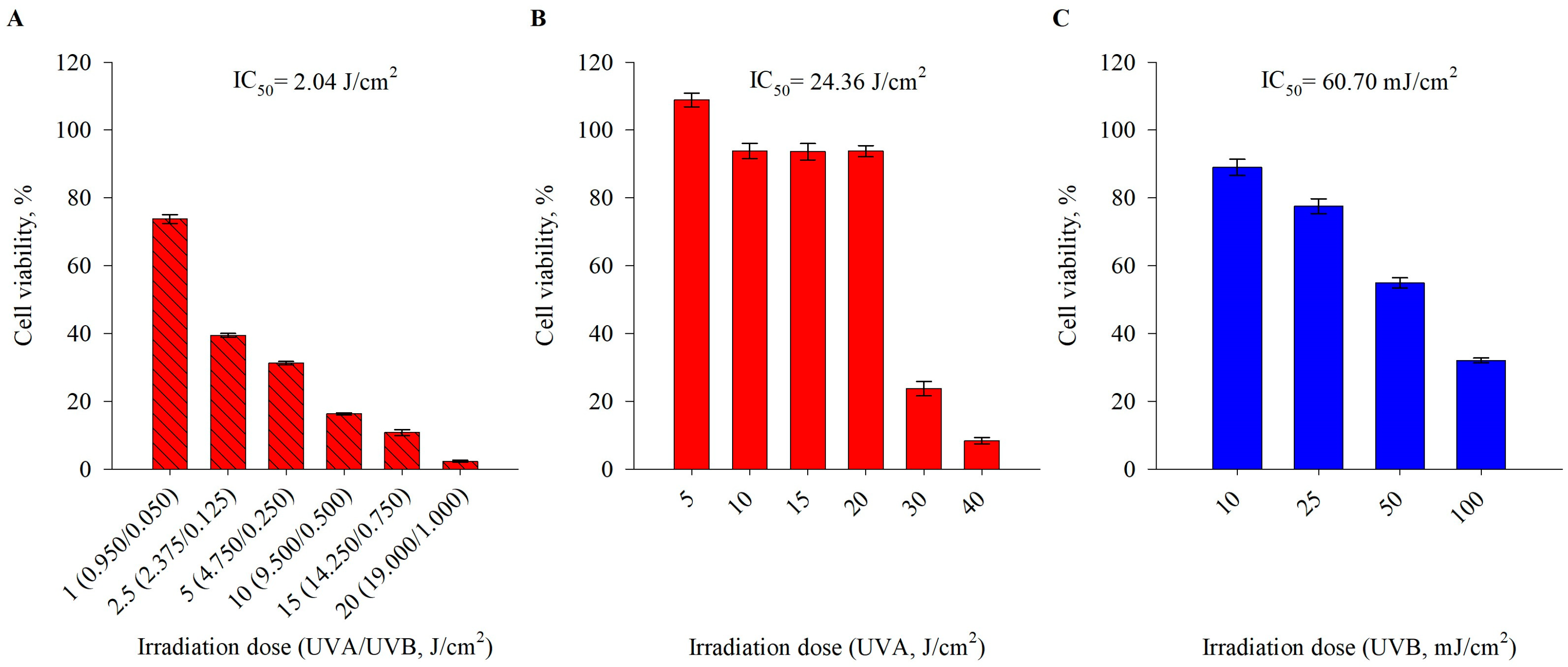
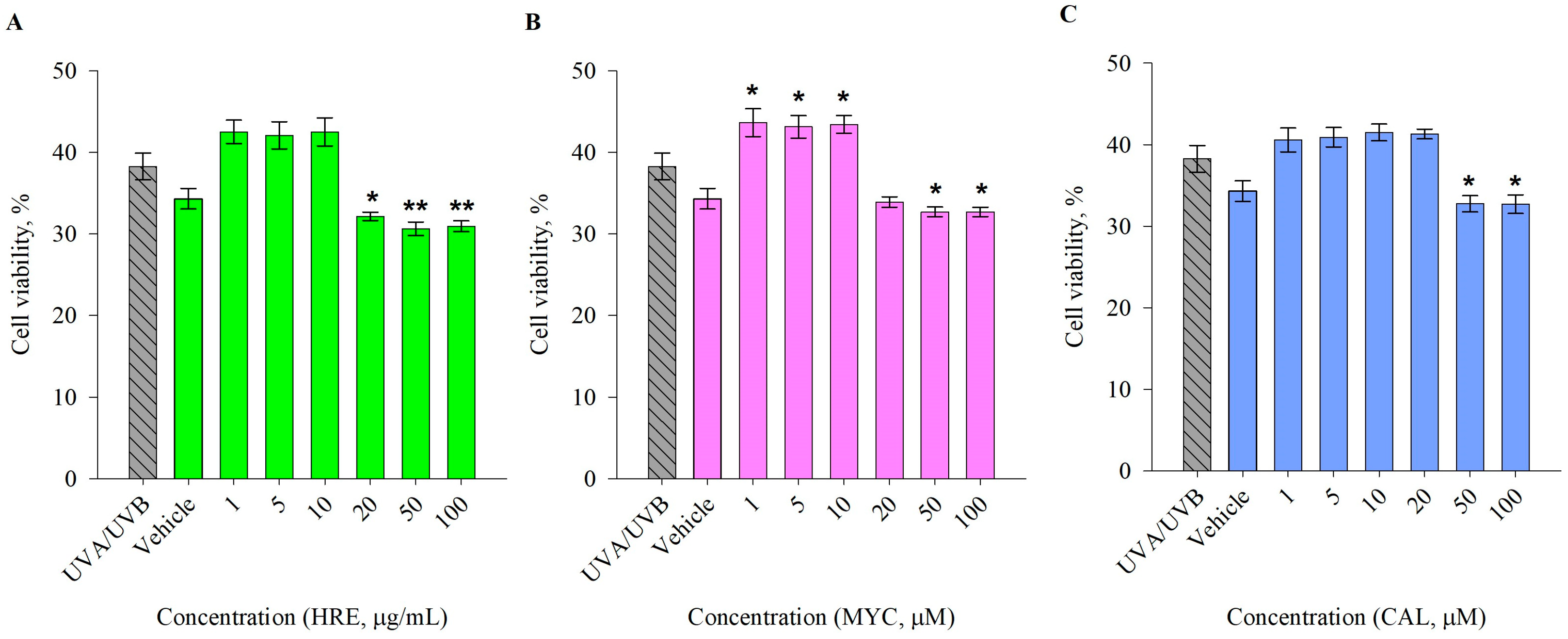
2.1. Pre-Treatment with H. rhodopensis Extract and Its Secondary Metabolites Induces Photoprotective Effects in UVA/UVB-Exposed Human Keratinocytes
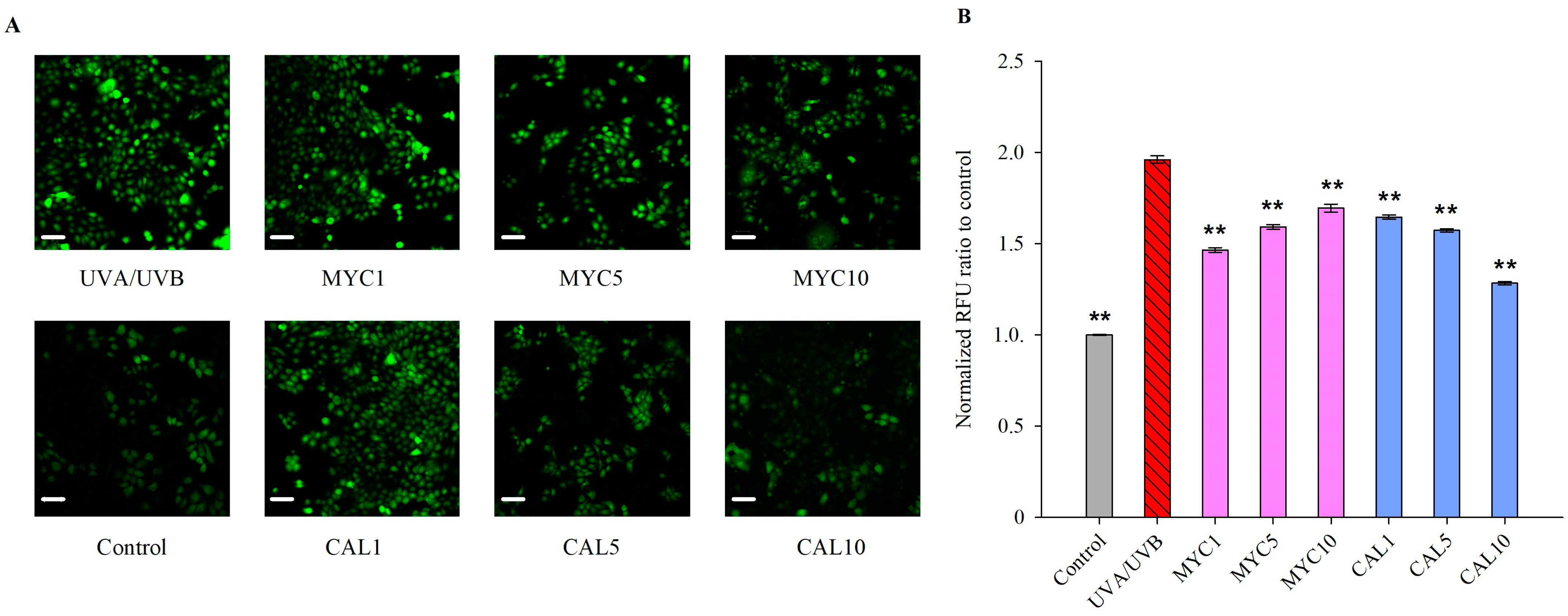
2.2. Myconoside and Calceolarioside E Alleviate UVA/UVB-Induced ROS Generation in Human Keratinocytes
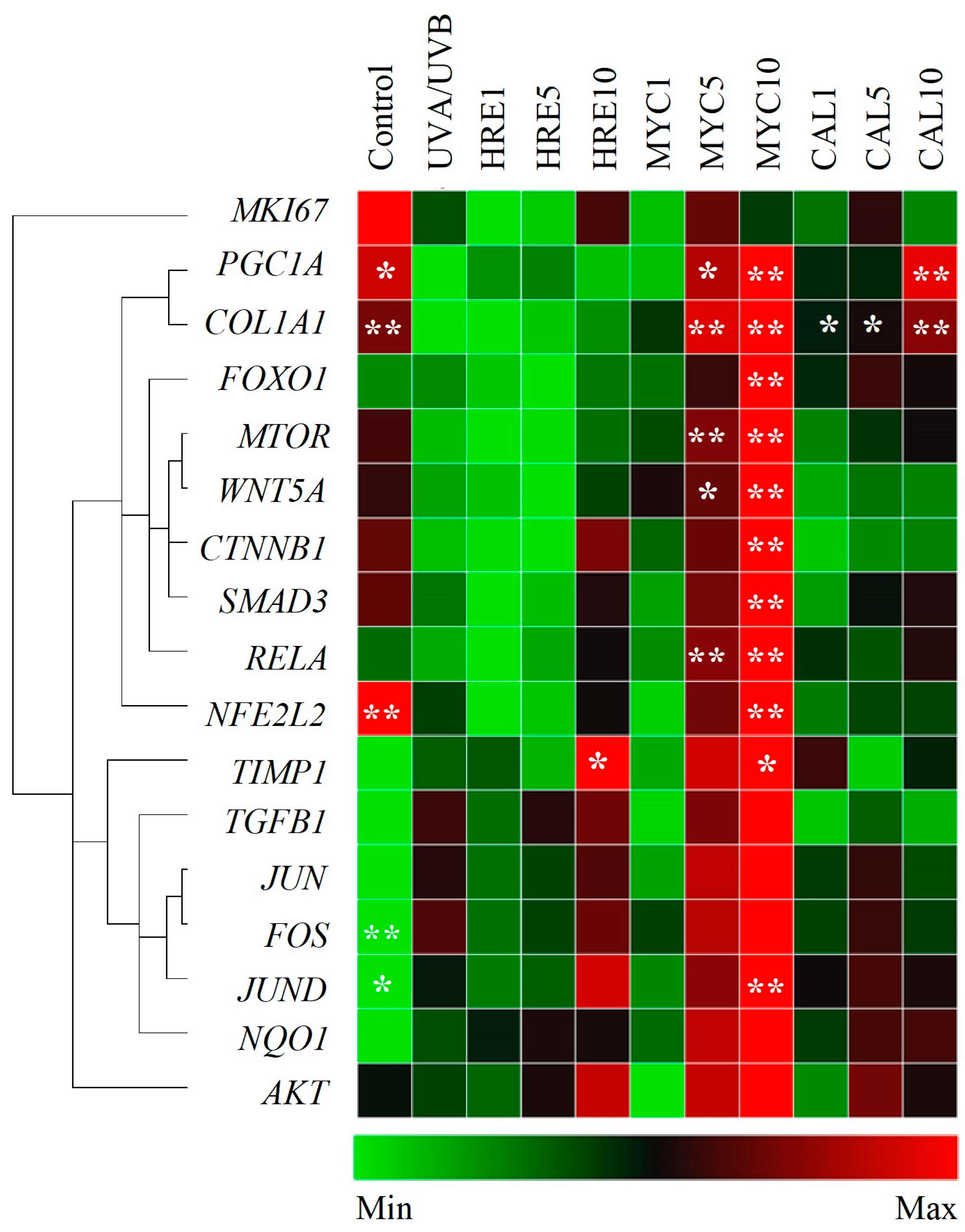
2.3. Myconoside and Calceolarioside E from H. rhodopensis Modulate the Gene Expression Profile of UVA/UVB-Irradiated Keratinocytes
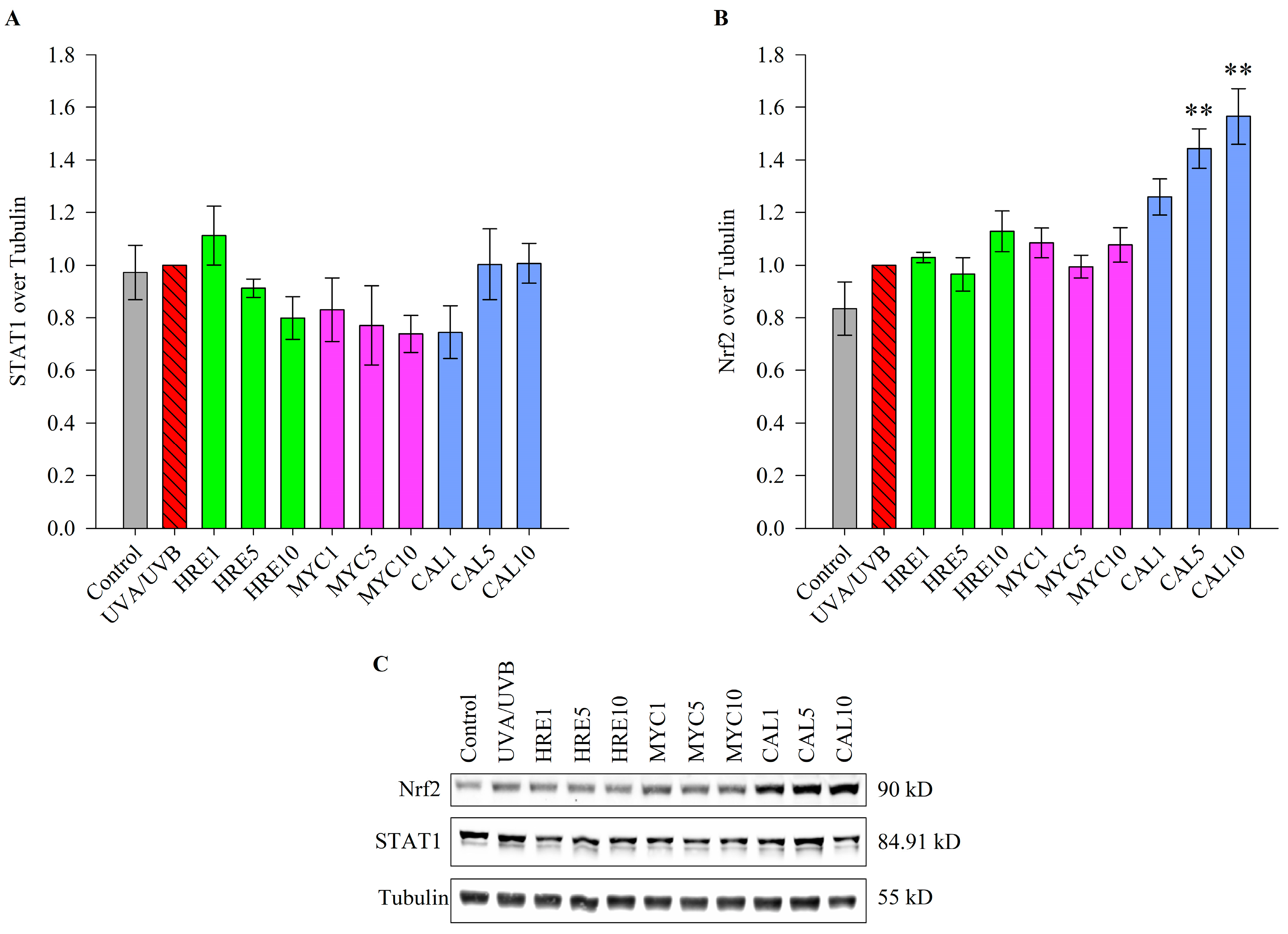
2.4. Calceolarioside E Activates the Transcription Factor NRF2 in UVR-Stimulated Keratinocytes
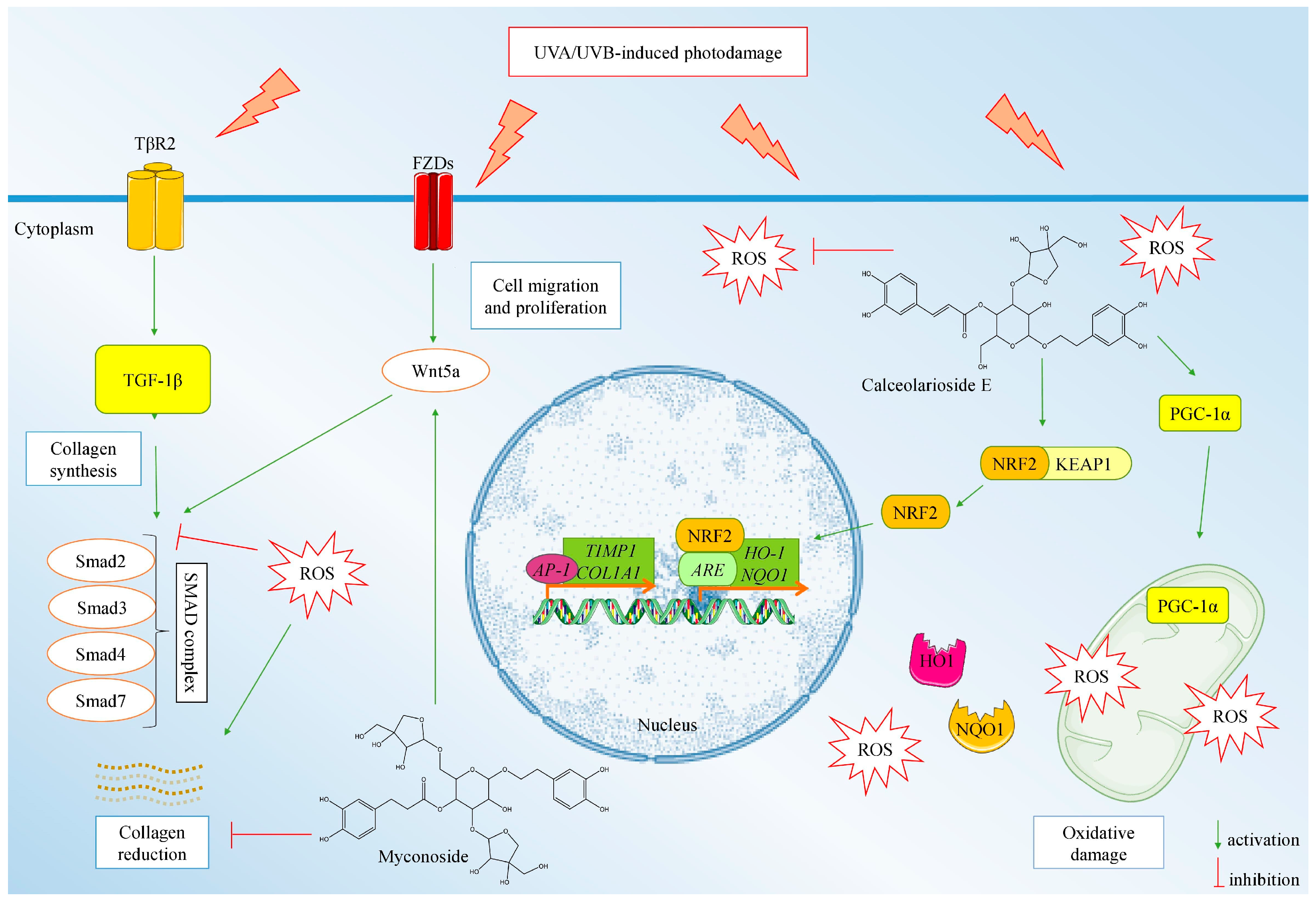
2.5. Proposed Model Mechanism of the Anti-Photoaging Activity of MYC and CAL Isolated from H. rhodopensis Extract
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture, Irradiation, and Treatment
4.3. Viability and Photoprotection Assays
4.4. Fluorescent Microcopy for UV-Induced ROS Formation
4.5. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.6. Western Blotting
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gegotek, A.; Bielawska, K.; Biernacki, M.; Dobrzyńska, I.; Skrzydlewska, E. Time-dependent effect of rutin on skin fibroblasts membrane disruption following UV radiation. Redox Biol. 2017, 12, 733–744. [Google Scholar] [CrossRef] [PubMed]
- de Assis, L.; Tonolli, P.; Moraes, M.; Baptista, M.; Castrucci, A. How does the skin sense sun light? An integrative view of lightsensing molecules. J. Photochem. Photobiol. C-Photochem. Rev. 2021, 47, 100403. [Google Scholar] [CrossRef]
- Fitsiou, E.; Pulido, T.; Campisi, J.; Alimirah, F.; Demaria, M. Cellular senescence and the senescence-associated secretory phenotype as drivers of skin photoaging. J. Investig. Dermatol. 2021, 141, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Kahremany, S.; Hofmann, L.; Gruzman, A.; Dinkova-Kostova, A.T.; Cohen, G. NRF2 in dermatological disorders: Pharmacological activation for protection against cutaneous photodamage and photodermatosis. Free. Radic. Biol. Med. 2022, 188, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.T.; Ganju, P.; Ramkumar, A.; Grover, R.; Sgokhale, R. Multifaceted pathways protect human skin from UV radiation. Nat. Chem. Biol. 2014, 10, 542–551. [Google Scholar] [CrossRef]
- Bustamante, M.; Hernandez-Ferre, C.; Tewari, A.; Sarria, Y.; Harrison, G.I.; Puigdecanet, E.; Nonell, L.; Kang, W.; Friedlander, M.R.; Estivill, X.; et al. Dose and time effects of solar-simulated ultraviolet radiation on the in vivo human skin transcriptome. Br. J. Dermatol. 2020, 182, 1458–1468. [Google Scholar] [CrossRef]
- Garg, C.; Sharma, H.; Garg, M. Skin photo-protection with phytochemicals against photo-oxidative stress, photo-carcinogenesis, signal transduction pathways and extracellular matrix remodelling—An overview. Ageing Res. Rev. 2020, 62, 101127. [Google Scholar] [CrossRef]
- Kim, J.-M.; Chung, K.-S.; Yoon, Y.-S.; Jang, S.-Y.; Heo, S.-W.; Park, G.; Jang, Y.P.; Ahn, S.H.; Shin, Y.K.; Lee, S.H.; et al. Dieckol isolated from Eisenia bicyclis ameliorates wrinkling and improves skin hydration via MAPK/AP-1 and TGF-β/Smad signalling pathways in UVB-irradiated hairless mice. Mar. Drugs 2022, 20, 779. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Dańczak-Pazdrowska, A.; Gornowicz-Porowska, J.; Polańska, A.; Krajka-Kuźniak, V.; Stawny, M.; Gostyńska, A.; Rubiś, B.; Nourredine, S.; Ashiqueali, S.; Schneider, A.; et al. Cellular senescence in skin-related research: Targeted signalling pathways and naturally occurring therapeutic agents. Aging Cell 2023, 22, e13845. [Google Scholar] [CrossRef]
- Akhmetshina, A.; Palumbo, K.; Dees, C.; Bergmann, C.; Venalis, P.; Zerr, P.; Horn, A.; Kireva, T.; Beyer, C.; Zwerina, J.; et al. Activation of canonical Wnt signalling is required for TGF-β-mediated fibrosis. Nat. Commun. 2012, 3, 735. [Google Scholar] [CrossRef] [PubMed]
- Noh, D.; Choi, J.G.; Huh, E.; Oh, M.S. Tectorigenin, a flavonoid-based compound of leopard lily rhizome, attenuates UV-B-induced apoptosis and collagen degradation by inhibiting oxidative stress in human keratinocytes. Nutrients 2018, 10, 1998. [Google Scholar] [CrossRef]
- Lee, H.; Kong, G.; Park, J.; Park, J. The potential inhibitory effect of ginsenoside Rh2 on mitophagy in UV-irradiated human dermal fibroblasts. J. Ginseng Res. 2022, 46, 64–65. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-K.; Kwon, O.-Y.; Lee, S.-H. Kaempferide prevents photoaging of ultraviolet-B irradiated NIH-3T3 cells and mouse skin via regulating the ROS-mediated signalling. Antioxidants 2023, 12, 11. [Google Scholar] [CrossRef]
- Kurinna, S.; Schäfer, M.; Ostano, P.; Karouzakis, E.; Chiorino, G.; Bloch, W.; Bachmann, A.; Gay, S.; Garrod, D.; Lefort, K.; et al. A novel Nrf2-miR-29-desmocollin-2 axis regulates desmosome function in keratinocytes. Nat. Commun. 2014, 5, 5099. [Google Scholar] [CrossRef]
- Hao, D.; Wen, X.; Liu, L.; Wang, L.; Zhou, X.; Li, Y.; Zeng, X.; He, G.; Jiang, X. Sanshool improves UVB-induced skin photodamage by targeting JAK2/STAT3-dependent autophagy. Cell Death Dis. 2019, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Lee, J.S.; Hong, S.; Lim, T.-G.; Byun, S. Quercetin directly targets JAK2 and PKCδ and prevents UV-induced photoaging in human skin. Int. J. Mol. Sci. 2019, 20, 5262. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, S.; Liu, Y.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. A novel glyceroglycolipid from brown algae Ishige okamurae improve photoaging and counteract inflammation in UVB-induced HaCaT cells. Chem.-Biol. Interact. 2022, 351, 109737. [Google Scholar] [CrossRef]
- Li, C.; Fu, Y.; Dai, H.; Wang, Q.; Gao, R.; Zhang, Y. Recent progress in preventive effect of collagen peptides on photoaging skin and action mechanism. Food Sci. Hum. Wellness 2022, 11, 218–229. [Google Scholar] [CrossRef]
- Moon, N.R.; Kang, S.; Park, S. Consumption of ellagic acid and dihydromyricetin synergistically protects against UV-B induced photoaging, possibly by activating both TGF-β1 and wnt signalling pathways. J. Photochem. Photobiol. B-Biol. 2018, 178, 92–100. [Google Scholar] [CrossRef]
- Xue, N.; Liu, Y.; Jin, J.; Ji, M.; Chen, X. Chlorogenic acid prevents UVA-induced skin photoaging through regulating collagen metabolism and apoptosis in human dermal fibroblasts. Int. J. Mol. Sci. 2022, 23, 6941. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, S.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. Trehalose against UVB-induced skin photoaging by suppressing MMP expression and enhancing procollagen I synthesis in HaCaT cells. J. Func. Food 2020, 74, 104198. [Google Scholar] [CrossRef]
- Tao, S.; Park, S.L.; de la Vega, M.R.; Zhang, D.D.; Wondrak, G.T. Systemic administration of the apocarotenoid bixin protects skin against solar UV-induced damage through activation of NRF2. Free Radic. Biol. Med. 2015, 89, 690–700. [Google Scholar] [CrossRef]
- Tao, S.; Zheng, Y.; Lau, A.; Jaramillo, M.C.; Chau, B.T.; Lantz, R.C.; Wong, P.K.; Wondrak, G.T.; Zhang, D.D. Tanshinone I activates the Nrf2-dependent antioxidant response and protects against As(III)-induced lung inflammation in vitro and in vivo. Antiox. Redox Singal. 2013, 19, 1647–1661. [Google Scholar] [CrossRef]
- Li, Q.; Bai, D.; Qin, L.; Shao, M.; Zhang, S.; Yan, C.; Yu, G.; Hao, J. Protective effect of d-tetramannuronic acid tetrasodium salt on UVA-induced photo-aging in HaCaT cells. Biomed. Pharmacother. 2020, 126, 110094. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.L.; Kensler, T.W.; et al. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef]
- Vasileva, L.V.; Savova, M.S.; Amirova, K.M.; Dinkova-Kostova, A.T.; Georgiev, M.I. Obesity and NRF2-mediated cytoprotection: Where is the missing link? Pharmacol. Res. 2020, 156, 104760. [Google Scholar] [CrossRef] [PubMed]
- Amirova, K.M.; Dimitrova, P.A.; Marchev, A.S.; Krustanova, S.V.; Simova, S.D.; Alipieva, K.I.; Georgiev, M.I. Biotechnologically-produced myconoside and calceolarioside E induce Nrf2 expression in neutrophils. Int. J. Mol. Sci. 2021, 22, 1759. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-L.; Lee, C.-L.; Korivi, M.; Liao, J.-W.; Rajendran, P.; Wu, J.-J.; Hseu, Y.-C. Zerumbone protects human skin keratinocytes against UVA-irradiated damages through NRF2 induction. Biochem. Pharmacol. 2018, 148, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Kim, K.C.; Kang, K.A.; Fernando, P.D.S.M.; Herath, H.M.U.L.; Hyun, J.W. Phloroglucinol attenuates ultraviolet B-induced 8-oxoguanine formation in human HaCaT keratinocytes through Akt and Erk-mediated NRF2/Ogg1 signalling pathways. Biomol. Ther. 2021, 29, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, X.; Lei, X.; Lei, L.; Zhao, J.; Zeng, K.; Ming, J. Premna microphylla Turcz pectin protected UVB-induced skin aging in BALB/c-nu mice via NRF2 pathway. Int. J. Biol. Macromol. 2022, 215, 12–22. [Google Scholar] [CrossRef]
- Tanveer, M.A.; Rashid, H.; Nazir, L.A.; Archoo, S.; Shahid, N.H.; Ragni, G.S.; Umar, A.; Tasduq, S.A. Trigonelline, a plant derived alkaloid prevents ultraviolet-B-induced oxidative DNA damage in primary human dermal fibroblasts and BALB/c mice via modulation of phosphoinositide 3-kinase-Akt-Nrf2 signalling axis. Exp. Gerontol. 2023, 171, 112028. [Google Scholar] [CrossRef]
- Gravel, S.-P.; Khalifa, Y.B.; McGuirk, S.; St-Louis, C.; Laurin, K.M.; Lavallée, É.; Benas, D.; Desbouis, S.; Amaral, F.; D’Amours, D.; et al. PGC-1s shape epidermal physiology by modulating keratinocyte proliferation and terminal differentiation. iScience 2023, 26, 106314. [Google Scholar] [CrossRef] [PubMed]
- Correia-Melo, C.; Marques, F.D.; Anderson, R.; Hewitt, G.; Hewitt, R.; Cole, J.; Carroll, B.M.; Miwa, S.; Birch, J.; Merz, D.; et al. Mitochondria are required for pro-ageing features of the senescent phenotype. Embo J. 2016, 35, 724–742. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Sun, Q.; Hu, A.; Hou, L.; Li, J.; Cai, X.; Yu, C.-P. Pharmaceuticals and personal care products in a mesoscale subtropical watershed and their application as sewage markers. J. Hazard. Mater. 2014, 280, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Young, A.R.; Claveau, J.; Rossi, A.B. Ultraviolet radiation and the skin: Photobiology and sunscreen photoprotection. J. Am. Acad. Dermatol. 2017, 76, S100–S109. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Mohammed, Y.; Prow, T.W. Advances and controversies in studying sunscreen delivery and toxicity. Adv. Drug Deliv. Rev. 2020, 153, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.M.; Martins, S.S.; Barbosa, G.L.F.; e Silva, E.G.N.; Fonseca, M.J.V.; de Freitas, L.A.P. Natural component and solid lipid microparticles of solar filter in sunscreen: Photoprotective and photostability effect enhancement. J. Drug Deliv. Sci. Technol. 2023, 88, 104860. [Google Scholar] [CrossRef]
- Tomazelli, L.C.; Ramos, M.M.A.; Sause, R.; Candido, T.M.; Sarruf, F.D.; Pinto, C.A.S.O.; de Oliveira, C.A.; Rosado, C.; Velasco, M.V.R.; Baby, A.R. SPF enhancement provided by rutin in a multifunctional sunscreen. Int. J. Pharm. 2018, 552, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Chaiprasongsuk, A.; Onkoksoong, T.; Pleumsamran, T.; Limsaengurai, S.; Panich, U. Photoprotection by dietary phenolics against melanogenesis induced by UVA through Nrf2-dependent antioxidant responses. Redox Biol. 2016, 8, 79–90. [Google Scholar] [CrossRef]
- Peres, D.D.; Sarruf, F.D.; de Oliveira, C.A.; Velasco, M.V.R.; Baby, A.R. Ferulic acid photoprotective properties in association with UV filters: Multifunctional sunscreen with improved SPF and UVA-PF. J. Photochem. Photobiol. B-Biol. 2018, 185, 46–49. [Google Scholar] [CrossRef]
- Rosado, C.; Tokunaga, V.K.; Sauce, R.; de Oliveira, C.A.; Sarruf, F.D.; Parise-Filho, R.; Mauricio, E.; de Almeida, T.S.; Velasco, M.V.R.; Baby, A.R. Another reason for using caffeine in dermocosmetics: Sunscreen adjuvant. Front. Physiol. 2019, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Chaiprasongsuk, A.; Panich, U. Role of phytochemicals in skin photoprotection via regulation of Nrf2. Front. Pharmacol. 2022, 13, 823881. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.; Lyall, R.; Petrov, V.; Batels, D. Systems biology of resurrection plants. Cell. Mol. Life Sci. 2021, 78, 6365–6394. [Google Scholar] [CrossRef]
- Dell’Acqua, G.; Schweikert, K. Skin benefits of a myconoside-rich extract from resurrection plant Haberlea rhodopensis. Int. J. Cosmet. Sci. 2012, 34, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Spyridopoulou, K.; Kyriakou, S.; Nomikou, A.; Roupas, A.; Ermogenous, A.; Karamanoli, K.; Moyankova, D.; Djilianov, D.; Galanis, A.; Panayiotidis, M.I.; et al. Chemical profiling, antiproliferative and antimigratory capacity of Haberlea rhodopensis extracts in an in vitro platform of various human cancer cell lines. Antioxidants 2022, 11, 2305. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chang, C.-T.; Gowrisankar, Y.V.; Chen, X.-Z.; Lin, H.-C.; Yen, H.-R.; Yang, H.-L. Zerumbone exhibits antiphotoaging and dermatoprotective properties in ultraviolet A-irradiated human skin fibroblast cells via the activation of NRF2/ARE defensive pathway. Oxidative Med. Cell. Longev. 2019, 2019, 4098674. [Google Scholar] [CrossRef]
- Yosefzon, Y.; Soteriou, D.; Feldman, A.; Kostic, L.; Koren, E.; Brown, S.; Ankawa, R.; Sedov, E.; Glaser, F.; Fuchs, Y. Caspase-3 regulates YAP-dependent cell proliferation and organ size. Mol. Cell 2018, 70, 573–587. [Google Scholar] [CrossRef]
- Katayoshi, T.; Nakajo, T.; Tsuji-Naito, K. Restoring NAD+ by NAMPT is essential for the SIRT1/p53-mediated survival of UVA- and UVB-irradiated epidermal keratinocytes. J. Photochem. Photobiol. B-Biol. 2021, 221, 112238. [Google Scholar] [CrossRef]
- Oh, J.H.; Karadeniz, F.; Lee, J.I.; Park, S.Y.; Seo, Y.; Kong, C.-S. Anticatabolic and anti-inflammatory effects of myricetin 3-O-β-d-galactopyranoside in UVA-irradiated dermal cells via repression of MAPK/AP-1 and activation of TGFβ/Smad. Molecules 2020, 25, 1331. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, H.; Yang, Y.; Zhang, S.; Wang, J.; Zhang, D.; Yu, H. Metformin attenuates UVA-induced skin photoaging by suppressing mitophagy and the PI3K/AKT/mTOR pathway. Int. J. Mol. Sci. 2022, 23, 6960. [Google Scholar] [CrossRef] [PubMed]
- Charachit, N.; Sukhamwang, A.; Dejkriengkraikul, P.; Yodkeeree, S. Hyperoside and quercitrin in Houttuynia cordata extract attenuate UVB-induced human keratinocyte cell damage and oxidative stress via modulation of MAPKs and Akt signaling pathway. Antioxidants 2022, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Jiratchayamaethasakul, C.; Lee, S.-H. Protocatechuic aldehyde attenuates UVA-induced photoaging in human dermal fibroblast cells by suppressing MAPKs/AP-1 and NF-κB signaling pathways. Int. J. Mol. Sci. 2020, 21, 4619. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Bai, D.; Qin, L.; Shao, M.; Liu, X.; Zhang, S.; Yan, C.; Yu, G.; Hao, J. Protective effect of l-hexaguluroic acid hexasodium salt on UVA-induced photo-aging in HaCaT cells. Int. J. Mol. Sci. 2020, 21, 1201. [Google Scholar] [CrossRef]
- Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.-S. Syringaresinol inhibits UVA-induced MMP-1expression by suppression of MAPK/AP-1 signaling in HaCaT keratinocytes and human dermal fibroblasts. Int. J. Mol. Sci. 2020, 21, 3981. [Google Scholar] [CrossRef]
- Koycheva, I.K.; Vasileva, L.V.; Amirova, K.M.; Marchev, A.S.; Balcheva-Sivenova, Z.P.; Georgiev, M.I. Biotechnologically produced Lavandula angustifolia Mill. Extract rich in rosmarinic acid resolves psoriasis-related inflammation through Janus kinase/signal transducer and activator of transcription signaling. Front. Pharmacol. 2021, 12, 680168. [Google Scholar] [CrossRef]
- Sánchez-Marzo, N.; Pérez-Sánchez, A.; Barrajón-Catalán, E.; Castillo, J.; Herranz-López, M.; Micol, V. Rosemary diterpenes and flavanone aglycones provide improved genoprotection against UV-Induced DNA damage in a human skin cell model. Antioxidants 2020, 9, 255. [Google Scholar] [CrossRef]
- Martínez-Gutiérrez, A.; Fernández-Duran, I.; Marazuela-Duque, A.; Simonet, N.G.; Yousef, I.; Martínez-Rovira, I.; Martínez-Hoyos, J.; Vaquero, A. Shikimic acid protects skin cells from UV-induced senescence through activation of the NAD+-dependent deacetylase SIRT1. Aging 2021, 13, 12308–12333. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.; Heo, S.-J.; Dias, M.D.; Madusanka, D.D.; Han, E.-J.; Kim, M.-J.; Sanjeewa, K.; Lee, K.; Ahn, G. (-)-Loliolide isolated from Sargassum horneri abate UVB-induced oxidative damage in human dermal fibroblasts and subside ECM degradation. Mar. Drugs 2021, 19, 435. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Li, H.; Hwang, E.; Park, B.; Xiao, Y.-K.; Liu, S.; Fang, J.; Kim, Y.-J.; Yi, T.-H.; Cai, C. Chemical distance measurement and system pharmacology approach uncover the novel protective effects of biotransformed ginsenoside C-Mc against UVB-irradiated photoaging. Oxidative Med. Cell. Longev. 2022, 2022, 4691576. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-W.; Hung, Y.-C.; Lin, T.-Y.; Fang, J.-Y.; Yang, P.-M.; Chen, M.-H.; Pan, T.-L. Comparison of the biological impact of UVA and UVB upon the skin with functional proteomics and immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.; Kanimozhi, G.; Madahavan, N.R.; Agilan, B.; Ganesan, M.; Prasad, N.R.; Rathinaraj, P. Alpha-pinene attenuates UVA-induced photoaging through inhibition of matrix metalloproteinases expression in mouse skin. Life Sci. 2019, 217, 110–118. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoykova, I.D.; Koycheva, I.K.; Binev, B.K.; Mihaylova, L.V.; Benina, M.Y.; Alipieva, K.I.; Georgiev, M.I. Myconoside and Calceolarioside E Restrain UV-Induced Skin Photoaging by Activating NRF2-Mediated Defense Mechanisms. Int. J. Mol. Sci. 2024, 25, 2441. https://doi.org/10.3390/ijms25042441
Stoykova ID, Koycheva IK, Binev BK, Mihaylova LV, Benina MY, Alipieva KI, Georgiev MI. Myconoside and Calceolarioside E Restrain UV-Induced Skin Photoaging by Activating NRF2-Mediated Defense Mechanisms. International Journal of Molecular Sciences. 2024; 25(4):2441. https://doi.org/10.3390/ijms25042441
Chicago/Turabian StyleStoykova, Iva D., Ivanka K. Koycheva, Biser K. Binev, Liliya V. Mihaylova, Maria Y. Benina, Kalina I. Alipieva, and Milen I. Georgiev. 2024. "Myconoside and Calceolarioside E Restrain UV-Induced Skin Photoaging by Activating NRF2-Mediated Defense Mechanisms" International Journal of Molecular Sciences 25, no. 4: 2441. https://doi.org/10.3390/ijms25042441