
Genomic Insights into High-Altitude Adaptation: A Comparative Analysis of Roscoea alpina and R. purpurea in the Himalayas

Abstract
:1. Introduction
2. Results
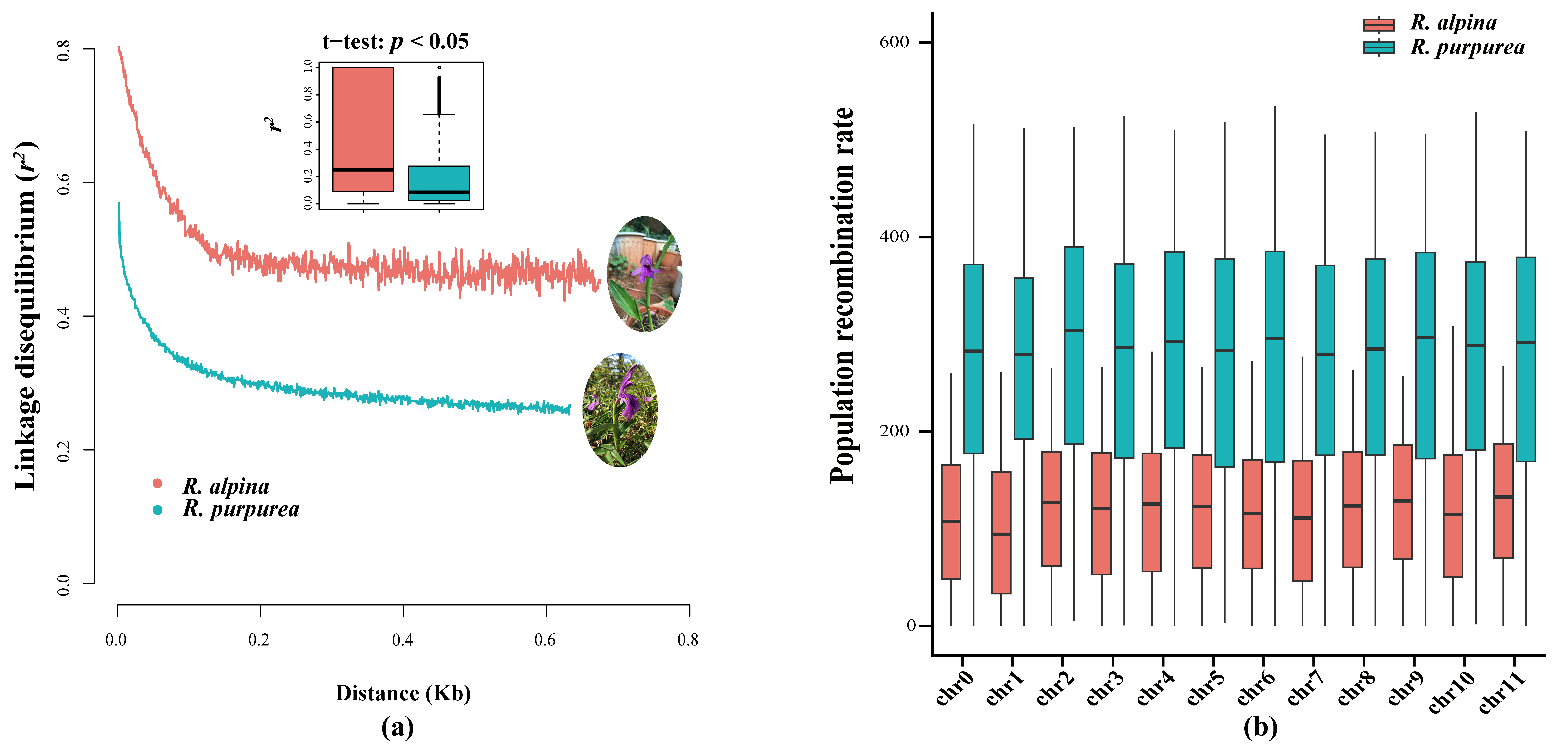
2.1. Genomic Feature Investigation
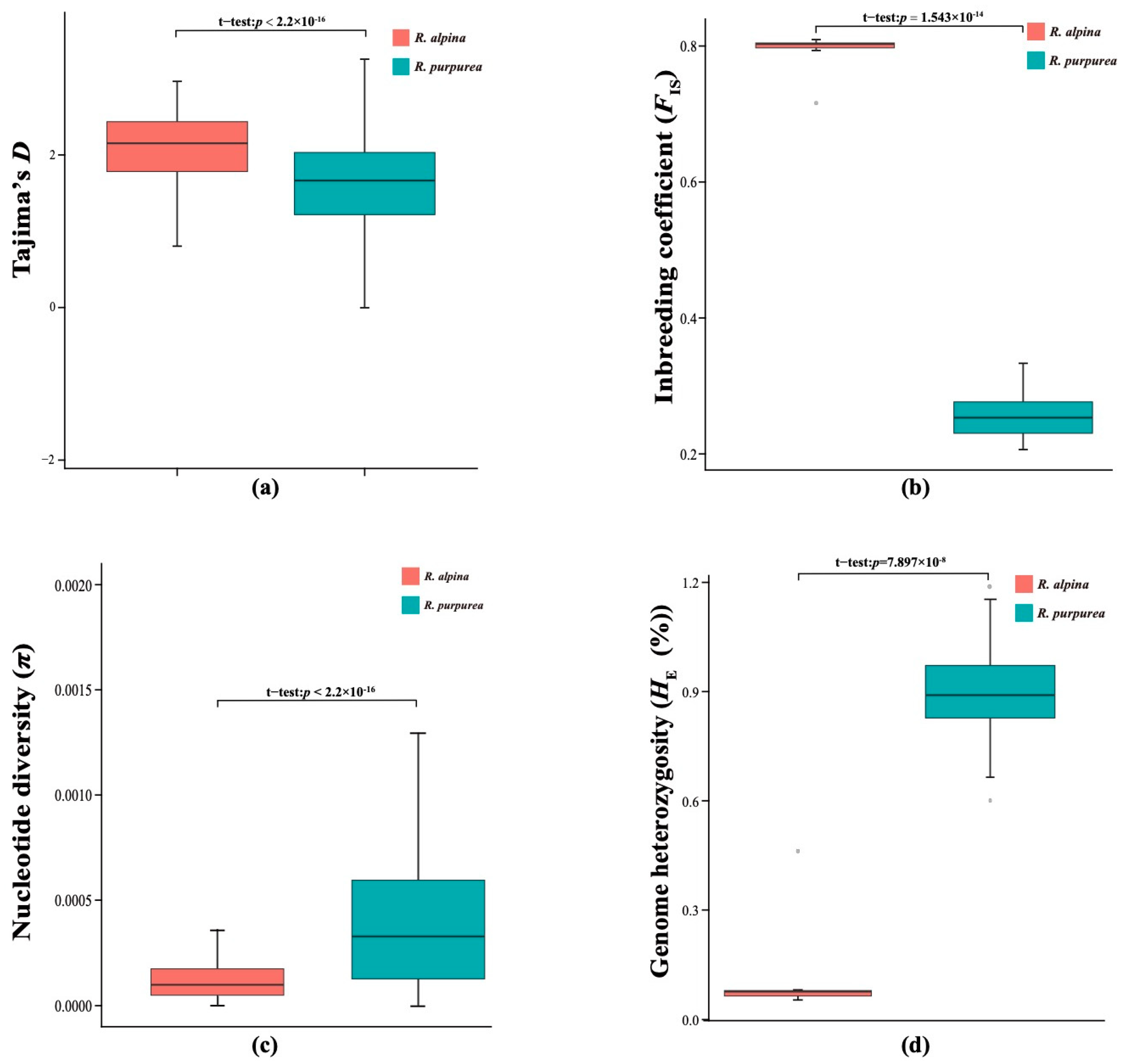
2.2. Difference in Demographic History
2.3. Candidate Genes Associated with High-Altitude Adaptation
3. Discussion
3.1. Genomic Features for Adaptive Evolution in the High Himalayas
3.2. Difference in Demographic History within the Himalayas
3.3. Candidate Genes Associated with High-Altitude Adaptation
4. Materials and Methods
4.1. Resequencing and Variant Discovery
4.2. Genomic Feature Investigation
4.3. Demographic Inferences
4.4. Identification of Candidate Genes Associated with High-Altitude Adaptation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, S.; Singh, R. High-altitude clear-sky direct solar ultraviolet irradiance at Leh and Hanle in the western Himalayas: Observations and model calculations. J. Geophys. Res. Atmos. 2004, 109, D19201. [Google Scholar] [CrossRef]
- Stres, B.; Sul, W.J.; Murovec, B.; Tiedje, J.M. Recently deglaciated high-altitude soils of the Himalaya: Diverse environments, heterogenous bacterial communities and long-range dust inputs from the upper troposphere. PLoS ONE 2013, 8, e76440. [Google Scholar] [CrossRef]
- Monge, C.; León-Velarde, F. Physiological adaptation to high altitude: Oxygen transport in mammals and birds. Physiol. Rev. 1991, 71, 1135–1172. [Google Scholar] [CrossRef]
- Storz, J.F.; Scott, G.R.; Cheviron, Z.A. Phenotypic plasticity and genetic adaptation to high-altitude hypoxia in vertebrates. J. Exp. Biol. 2010, 213, 4125–4136. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479485. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.R.; Zhang, Z.H.; Zhang, C.L.; Zhang, J.Q.; Ran, A. Ecophysiological responses of three tree species to a high-altitude environment in the southeastern Tibetan Plateau. Forests 2018, 9, 48. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glemin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Halbritter, A.H.; Fior, S.; Keller, I.; Billeter, R.; Edwards, P.J.; Holderegger, R.; Karrenberg, S.; Pluess, A.R.; Widmer, A.; Alexander, J.M. Trait differentiation and adaptation of plants along elevation gradients. J. Evol. Biol. 2018, 31, 784–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kuang, T.; Dong, W.; Qian, Z.; Zhang, H.; Landis, J.B.; Feng, T.; Li, L.; Sun, Y.; Huang, J.; et al. Genomic convergence underlying high-altitude adaptation in alpine plants. J. Integr. Plant Biol. 2023, 65, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, G.K.; Dalrymple, S.E.; Pritchard, H.W. Seed survival at low temperatures: A potential Sselecting factor influencing community level changes in high altitudes under climate change. Crit. Rev. Plant Sci. 2020, 39, 479–492. [Google Scholar] [CrossRef]
- Tyagi, A.; Yadav, A.; Tripathi, A.M.; Roy, S. High light intensity plays a major role in emergence of population level variation in Arabidopsis thaliana along an altitudinal gradient. Sci. Rep. 2016, 23, 26160. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Sun, Y.; Yang, G.; Li, L.; Sun, W.; Wang, Z.; Zhang, H.; Li, Y. Natural variation in stress response induced by low CO2 in Arabidopsis thaliana. Open Life Sci. 2020, 21, 923–938. [Google Scholar] [CrossRef] [PubMed]
- León, J.; Castillo, M.C.; Gayubas, B. The hypoxia-reoxygenation stress in plants. J. Exp. Bot. 2021, 72, 5841–5856. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.F.; Wang, Y.B.; Zhang, J.B.; Maddock, S.; Snook, M.; Peterson, T. A maize QTL for silk maysin levels contains duplicated Myb-homologous genes which jointly regulate flavone biosynthesis. Plant Mol. Biol. 2003, 52, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Casati, P.; Walbot, V. Differential accumulation of maysin and rhamnosylisoorientin in leaves of high-altitude landraces of maize after UV-B exposure. Plant Cell Environ. 2005, 28, 788–799. [Google Scholar] [CrossRef]
- Flood, P.J.; Hancock, A.M. The genomic basis of adaptation in plants. Curr. Opin. Plant Biol. 2017, 36, 88–94. [Google Scholar] [CrossRef]
- Tiffin, P.; Ross-Ibarra, J. Advances and limits of using population genetics to understand local adaptation. Trends Ecol. Evol. 2014, 29, 673–780. [Google Scholar] [CrossRef]
- Stapley, J.; Reger, J.; Feulner, P.G.D.; Smadja, C.; Galindo, J.; Ekblom, R.; Bennison, C.; Ball, A.D.; Beckerman, A.P.; Slate, J. Adaptation genomics: The next generation. Trends Ecol. Evol. 2010, 25, 705–712. [Google Scholar] [CrossRef]
- Ekblom, R.; Galindo, J. Applications of next generation sequencing in molecular ecology of non-model organisms. Heredity 2011, 107, 1–15. [Google Scholar] [CrossRef]
- Zaborowska, J.; Labiszak, B.; Perry, A.; Cavers, S.; Wachowiak, W. Candidate genes for the high-altitude adaptations of two mountain pine taxa. Int. J. Mol. Sci. 2021, 22, 3477. [Google Scholar] [CrossRef]
- Abbas, M.; Sharma, G.; Dambire, C.; Marquez, J.; Alonso-Blanco, C.; Proano, K.; Holdsworth, M.J. An oxygen-sensing mechanism for angiosperm adaptation to altitude. Nature 2022, 606, 565–569. [Google Scholar] [CrossRef]
- Wu, Z.Y. Origin and Evolution of the Flora of Tibet; Science Press: Beijing, China, 1988. [Google Scholar]
- Chang, H.T.; Kong, Y.C.; But, P.H. The origin and its affinity of the Nepalese flora. Acta Sci. Nat. Univ. Sunyatseni 1988, 2, 1–12. [Google Scholar]
- Singh, J.S.; Singh, S.P. Forest vegetation of the Himalaya. Bot. Rev. 1987, 53, 80–192. [Google Scholar] [CrossRef]
- Vetaas, O.R.; Grytnes, J.A. Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Glob. Ecol. Biogeogr. 2002, 11, 291–301. [Google Scholar] [CrossRef]
- Zhao, J.L.; Paudel, B.R.; Yu, X.Q.; Zhang, J.; Li, Q.J. Speciation along the elevation gradient: Divergence of Roscoea species within the south slope of the Himalayas. Mol. Phylogenet. Evol. 2021, 164, 107292. [Google Scholar] [CrossRef]
- Zhao, J.L.; Zhong, J.S.; Fan, Y.L.; Xia, Y.M.; Li, Q.J. A preliminary species-level phylogeny of the alpine ginger Roscoea: Implications for speciation. J. Syst. Evol. 2017, 55, 215–224. [Google Scholar] [CrossRef]
- Cowley, E. The Genus Roscoea; The Royal Botanic Gardens: Kew, UK, 2007. [Google Scholar]
- Zhao, J.L.; Xia, Y.M.; Cannon, C.H.; Kress, W.J.; Li, Q.J. Evolutionary diversification of alpine ginger reflects the early uplift of the Himalayan–Tibetan Plateau and rapid extrusion of Indochina. Gondwana Res. 2016, 32, 232–241. [Google Scholar] [CrossRef]
- Ngamriabsakul, C.; Newman, M.; Cronk, Q. Phylogeny and disjunction in Roscoea (Zingiberaceae). Edinb. J. Bot. 2000, 57, 39–61. [Google Scholar] [CrossRef]
- Paudel, B.R.; Dyer, A.G.; Garcia, J.E.; Shrestha, M. The effect of elevational gradient on alpine gingers (Roscoea alpina and R. purpurea) in the Himalayas. PeerJ 2019, 7, e7503. [Google Scholar] [CrossRef]
- Paudel, B.R.; Shrestha, M.; Burd, M.; Adhikari, S.; Sun, Y.S.; Li, Q.J. Coevolutionary elaboration of pollination-related traits in an alpine ginger (Roscoea purpurea) and a tabanid fly in the Nepalese Himalayas. New Phytol. 2016, 211, 1402–1411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Liu, B.G.; Li, Y.; Liu, Y.; He, Y.X.; Qian, Z.Q.; Li, J.X. Landscape genetics reveals that adaptive genetic divergence in Pinus bungeana (Pinaceae) is driven by environmental variables relating to ecological habitats. BMC Evol. Biol. 2019, 19, 160. [Google Scholar] [CrossRef] [PubMed]
- Hartfield, M.; Bataillon, T.; Glemin, S. The evolutionary interplay between adaptation and self-fertilization. Trends Genet. 2017, 33, 420–431. [Google Scholar] [CrossRef]
- Walter, G.M.; Clark, J.; Cristaudo, A.; Terranova, D.; Nevado, B.; Catara, S.; Paunov, M.; Velikova, V.; Filatov, D.; Cozzolino, S.; et al. Adaptive divergence generates distinct plastic responses in two closely related Senecio species. Evolution 2022, 76, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Fulda, M.S.; Browse, J.A. Arabidopsis contains nine long-chain acyl-coenzyme a synthetase genes that participate in fatty acid and glycerolipid metabolism. Plant Physiol. 2002, 129, 1710–1722. [Google Scholar] [CrossRef]
- Naested, H.; Holm, A.; Jenkins, T.; Nielsen, H.B.; Harris, C.A.; Beale, M.H.; Andersen, M.; Mant, A.; Scheller, H.; Camara, B.; et al. Arabidopsis VARIEGATED 3 encodes a chloroplast-targeted, zinc-finger protein required for chloroplast and palisade cell development. J. Cell Sci. 2004, 117, 4807–4818. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, X.; Liu, J.; Wang, B.H.; Liu, B.L.; Wang, Y.Z. Pod shattering resistance associated with domestication is mediated by a NAC gene in soybean. Nat. Commun. 2014, 5, 3352. [Google Scholar] [CrossRef]
- Ohnishi, T. Recent advances in brassinosteroid biosynthetic pathway: Insight into novel brassinosteroid shortcut pathway. J. Pestic. Sci. 2018, 43, 159–167. [Google Scholar] [CrossRef]
- Yang, S.Q.; Li, W.Q.; Miao, H.; Gan, P.F.; Qiao, L.; Chang, Y.L.; Shi, C.H.; Chen, K.M. REL2, a gene encoding an unknown function protein which contains DUF630 and DUF632 domains controls leaf rolling in rice. Rice 2016, 9, 37. [Google Scholar] [CrossRef]
- Zhou, B.; Zeng, L. Elucidating the role of highly homologous Nicotiana benthamiana ubiquitin E2 gene family members in plant immunity through an improved virus-induced gene silencing approach. Plant Methods 2017, 13, 59. [Google Scholar] [CrossRef]
- Toller, A.; Brownfield, L.; Neu, C.; Twell, D.; Schulze-Lefert, P. Dual function of Arabidopsis glucan synthase-like genes GSL8 and GSL10 in male gametophyte development and plant growth. Plant J. 2008, 54, 911–923. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef]
- Huang, W.; Pi, L.; Liang, W.; Xu, B.; Wang, H.; Cai, R.; Huang, H. The proteolytic function of the Arabidopsis 26S proteasome is required for specifying leaf adaxial identity. Plant Cell 2006, 18, 2479–2492. [Google Scholar] [CrossRef]
- Pagnussat, G.C.; Yu, H.J.; Ngo, Q.A.; Rajani, S.; Mayalagu, S.; Johnson, C.S.; Capron, A.; Xie, L.F.; Ye, D.; Sundaresan, V. Genetic and molecular identification of genes required for female gametophyte development and function in Arabidopsis. Development 2005, 132, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Sugano, S.; Andronis, C.; Ong, M.; Green, R.; Tobin, E. The protein kinase CK2 is involved in regulation of circadian rhythms in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 12362–12366. [Google Scholar] [CrossRef]
- Triwitayakorn, K.; Njiti, V.N.; Iqbal, M.J.; Yaegashi, S.; Town, C.; Lightfoot, D.A. Genomic analysis of a region encompassing QRfs1 and QRfs2: Genes that underlie soybean resistance to sudden death syndrome. Genome 2005, 48, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Montoya, D.; Brueggeman, R.; McClean, P.E.; Osorno, J.M. Computational identification of receptor-like kinases “RLK” and receptor-like proteins “RLP” in legumes. BMC Genom. 2020, 21, 459. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xue, N.; Fu, X.; Zhang, H.; Li, G. Arabidopsis thaliana FAR-RED ELONGATED HYPOCOTYLS3 (FHY3) and FAR-RED-IMPAIRED RESPONSE1 (FAR1) modulate starch synthesis in response to light and sugar. New Phytol. 2017, 213, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Li, B.; He, W.; Yin, X.; Liu, S.; Lian, L.; Zhang, Y.; Liang, W.; Zhang, P. Physiological analysis of the effect of altitudinal gradients on Leymus secalinus on the Qinghai-Tibetan Plateau. PLoS ONE 2018, 13, e0202881. [Google Scholar] [CrossRef] [PubMed]
- Pierrugues, O.; Brutesco, C.; Oshiro, J.; Gouy, M.; Deveaux, Y.; Carman, G.M.; Thuriaux, P.; Kazmaier, M. Lipid phosphate phosphatases in Arabidopsis. Regulation of the AtLPP1 gene in response to stress. J. Biol. Chem. 2001, 276, 20300–20308. [Google Scholar] [CrossRef]
- Wright, S.I.; Kalisz, S.; Slotte, T. Evolutionary consequences of self-fertilization in plants. Proc. Biol. Sci. 2013, 280, 20130133. [Google Scholar] [CrossRef]
- van Ginkel, M.; Flipphi, R.C.H. Why self-fertilizing plants still exist in wild populations: Diversity assurance through stress-induced male sterility may promote selective outcrossing and recombination. Agronomy 2020, 10, 349. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Li, Q.J. Autonomous selfing provides reproductive assurance in an alpine ginger Roscoea schneideriana (Zingiberaceae). Ann. Bot. 2008, 102, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Bliss, L.C. Adaptations of arctic and alpine plants to environmental conditions. Arctic 1962, 15, 117–144. [Google Scholar] [CrossRef]
- García-Camacho, R.; Totland, Ø. Pollen limitation in the alpine: A meta-analysis. Arct. Antarctic. Alp. Res. 2009, 41, 103–111. [Google Scholar] [CrossRef]
- Bingham, R.A.; Orthner, A.R. Efficient pollination of alpine plants. Nature 1998, 391, 238–239. [Google Scholar] [CrossRef]
- Golding, G.B.; Strobeck, C. Linkage disequilibrium in a finite population that is partially selfing. Genetics 1980, 94, 777–789. [Google Scholar] [CrossRef]
- Pollak, E. On the theory of partially inbreeding finite populations. I. Partial selfing. Genetics 1987, 117, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S.; Long, A.D. The lowdown on linkage disequilibrium. Plant Cell 2003, 15, 1502–1506. [Google Scholar] [CrossRef]
- Paudel, B.R.; Shrestha, M.; Dyer, A.G.; Li, Q.J. Ginger and the beetle: Evidence of primitive pollination system in a Himalayan endemic alpine ginger (Roscoea alpina, Zingiberaceae). PLoS ONE 2017, 12, e0180460. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Buckler, E.S.t. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef]
- Felsenstein, J. The effect of linkage on directional selection. Genetics 1965, 52, 349–363. [Google Scholar] [CrossRef]
- Nagylaki, T. Quasilinkage equilibrium and the evolution of two-locus systems. Proc. Natl. Acad. Sci. USA 1974, 71, 526–530. [Google Scholar] [CrossRef]
- Kimura, M. Attainment of quasi linkage equilibrium when gene frequencies are changing by natural selection. Genetics 1965, 52, 875–890. [Google Scholar] [CrossRef]
- Charlesworth, B.; Morgan, M.T.; Charlesworth, D. The effect of deleterious mutations on neutral molecular variation. Genetics 1993, 134, 1289–1303. [Google Scholar] [CrossRef]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Stajich, J.E.; Hahn, M.W. Disentangling the effects of demography and selection in human history. Mol. Biol. Evol. 2005, 22, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Kosiński, P.; Sękiewicz, K.; Walas, Ł.; Boratynski, A.; Dering, M. Spatial genetic structure of the endemic alpine plant Salix serpillifolia: Genetic swamping on nunataks due to secondary colonization? Alp. Bot. 2019, 129, 107–121. [Google Scholar] [CrossRef]
- Bingham, R.A.; Ranker, T.A. Genetic diversity in alpine and foothill populations of Campanula rotundifolia (Campanulaceae). Int. J. Plant Sci. 2000, 161, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. An Introduction to Conservation Genetics, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Catalan, J.; Ninot, J.M.; Aniz, M.M. The high mountain conservation in a changing world. In High Mountain Conservation in a Changing World. Advances in Global Change Research; Catalan, J., Ninot, J.M., Aniz, M.M., Eds.; Springer: Cham, Switzerland, 2017; Volume 62, pp. 3–36. [Google Scholar] [CrossRef]
- Rana, S.K.; Price, T.D.; Qian, H. Plant species richness across the Himalaya driven by evolutionary history and current climate. Ecosphere 2019, 10, e02945. [Google Scholar] [CrossRef]
- Chen, Y.X.; Xu, J.; Liu, J.; Li, T.G.; Xiong, Z.F.; Zhang, P.; Yan, H. Climatic and tectonic constraints on the Plio-Pleistocene evolution of the Indonesian Throughflow intermediate water recorded by benthic δ18O from IODP site U1482. Quat. Sci. Rev. 2022, 295, 107666. [Google Scholar] [CrossRef]
- Gossmann, T.I.; Woolfit, M.; Eyre-Walker, A. Quantifying the variation in the effective population size within a genome. Genetics 2011, 189, 1389–1402. [Google Scholar] [CrossRef] [PubMed]
- Becher, H.; Jackson, B.C.; Charlesworth, B. Patterns of genetic variability in genomic regions with low rates of recombination. Curr. Biol. 2020, 30, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.L.; Halligan, D.L.; Haddrill, P.R.; Charlesworth, B. The relation between recombination rate and patterns of molecular evolution and variation in Drosophila melanogaster. Mol. Biol. Evol. 2014, 31, 1010–1028. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.C.H.; Harder, L.D.; Worley, A.C. The comparative biology of pollination and mating in flowering plants. Phil. Trans. R. Soc. Lond. B 1996, 351, 1271–1280. [Google Scholar] [CrossRef]
- Burri, R. Evolution: Small populations, low recombination, big trouble? Curr. Biol. 2021, 31, R282–R284. [Google Scholar] [CrossRef]
- Blumthaler, M.; Ambach, W.; Ellinger, R. Increase in solar UV radiation with altitude. J. Photoch. Photobio. B 1997, 39, 130–134. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.J.; Zuo, H.; Zheng, W.K.; Zhang, S.S.; Huang, Y.; Pingcuo, G.S.; Ying, H.; Zhao, F.; Li, Y.R.; et al. Genomic basis of high-altitude adaptation in Tibetan Prunus fruit trees. Curr. Biol. 2021, 31, 3848–3860.e8. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Browning, S.R.; Browning, B.L. Rapid and accurate haplotype phasing and missing-data inference for whole-genome association studies by use of localized haplotype clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef]
- Chikhi, R.; Medvedev, P. Informed and automated k-mer size selection for genome assembly. Bioinformatics 2014, 30, 31–37. [Google Scholar] [CrossRef]
- Gao, F.; Ming, C.; Hu, W.; Li, H. New software for the fast estimation of population recombination rates (FastEPRR) in the genomic era. G3-Genes Genom. Genet. 2016, 6, 1563–1571. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Vurture, G.W.; Sedlazeck, F.J.; Nattestad, M.; Underwood, C.J.; Fang, H.; Gurtowski, J.; Schatz, M.C. GenomeScope: Fast reference-free genome profiling from short reads. Bioinformatics 2017, 33, 2202–2204. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qu, W.; Obrycki, J.J.; Meng, L.; Zhou, X.; Chu, D.; Li, B. Optimizing sample size for population genomic study in a global invasive lady beetle, Harmonia Axyridis. Insects 2020, 5, 290. [Google Scholar] [CrossRef]
- Qu, W.M.; Liang, N.; Wu, Z.K.; Zhao, Y.G.; Chu, D. Minimum sample sizes for invasion genomics: Empirical investigation in an invasive whitefly. Ecol. Evol. 2019, 10, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Nadachowska-Brzyska, K.; Burri, R.; Smeds, L.; Ellegren, H. PSMC analysis of effective population sizes in molecular ecology and its application to black-and-white Ficedula flycatchers. Mol. Ecol. 2016, 25, 1058–1072. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, L.; Zhang, Z.; Li, M.; Wang, D.; Zhang, X.; Xi, Z.; Keefover-Ring, K.; Smart, L.B.; DiFazio, S.P.; et al. Phylogenomics of the genus Populus reveals extensive interspecific gene flow and balancing selection. New Phytol. 2020, 225, 1370–1382. [Google Scholar] [CrossRef]
- Weissensteiner, M.H.; Bunikis, I.; Catalán, A.; Francoijs, K.J.; Knief, U.; Heim, W.; Peona, V.; Pophaly, S.D.; Sedlazeck, F.J.; Suh, A.; et al. Discovery and population genomics of structural variation in a songbird genus. Nat. Commun. 2020, 11, 3403. [Google Scholar] [CrossRef]
- Li, K.; Zhang, S.; Song, X.; Weyrich, A.; Wang, Y.; Liu, X.; Wan, N.; Liu, J.; Lövy, M.; Cui, H.; et al. Genome evolution of blind subterranean mole rats: Adaptive peripatric versus sympatric speciation. Proc. Natl. Acad. Sci. USA 2020, 117, 32499–32508. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, W.; Lenstra, J.A.; Zheng, Z.; Wu, X.; Yang, J.; Li, B.; Yang, Y.; Qiu, Q.; Liu, H.; et al. Evolutionary origin of genomic structural variations in domestic yaks. Nat. Commun. 2023, 14, 5617. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Stresses | Genes | Annotations | References |
|---|---|---|---|
| Light intensity | AAEs | Participates in fatty acid and glycerolipid metabolism | [37] |
| Light intensity | VAR3 | Part of a protein complex required for chlorophyll and carotenoid synthesis | [38] |
| Light intensity, biotic and abiotic stress | SHAT1-5 | Provides high pod-shatter resistance | [39] |
| Light intensity, biotic and abiotic stress | BRs | Brassinosteroid biosynthesis | [40] |
| Biotic and abiotic stress | REL2 | Controls leaf rolling | [41] |
| Biotic and abiotic stress | E2 | Involved in plant biotic and abiotic stress responses | [42] |
| Biotic and abiotic stress | CALS9 | Involved in sporophytic and gametophytic development | [43] |
| Biotic and abiotic stress | StEXPA3 | Likely plays a role in tuber development | [44] |
| Biotic and abiotic stress | RPN8a | Determines leaf polarity | [45] |
| Biotic and abiotic stress | MEE40 | May be involved in female gametophyte development | [46] |
| Circadian clock | CK2 | Influences the circadian clock | [47] |
| Pathogen reduction | RFS2 | Involved in the partial resistance to the spread of Fusarium virguliforme root infections | [48] |
| Pathogen reduction | RLK | Regulates plant development and defense responses | [49] |
| Pathogen reduction, hypobaric hypoxia | PER65 | Involved in the response to wounding, pathogen attack, and oxidative stress | https://www.uniprot.org/uniprotkb/Q9FJR1/entry (accessed on 22 February 2023). |
| Hypobaric hypoxia, light intensity | FAR1 | Modulates starch synthesis in response to light and sugar | [50] |
| Hypobaric hypoxia, low temperatures | POD | Carbohydrate or soluble sugar synthesis in plants of alpine regions | [51] |
| Molecular damage | AtLPP1 | Reported to be induced by genotoxic stress (gamma ray or UV-B) and elicitor treatments with mastoparan and harpin | [52] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-L.; Li, L.; Paudel, B.R.; Zhao, J.-L. Genomic Insights into High-Altitude Adaptation: A Comparative Analysis of Roscoea alpina and R. purpurea in the Himalayas. Int. J. Mol. Sci. 2024, 25, 2265. https://doi.org/10.3390/ijms25042265
Wang Y-L, Li L, Paudel BR, Zhao J-L. Genomic Insights into High-Altitude Adaptation: A Comparative Analysis of Roscoea alpina and R. purpurea in the Himalayas. International Journal of Molecular Sciences. 2024; 25(4):2265. https://doi.org/10.3390/ijms25042265
Chicago/Turabian StyleWang, Ya-Li, Li Li, Babu Ram Paudel, and Jian-Li Zhao. 2024. "Genomic Insights into High-Altitude Adaptation: A Comparative Analysis of Roscoea alpina and R. purpurea in the Himalayas" International Journal of Molecular Sciences 25, no. 4: 2265. https://doi.org/10.3390/ijms25042265