
Dihydromyrcenol Modulates Involucrin Expression through the Akt Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
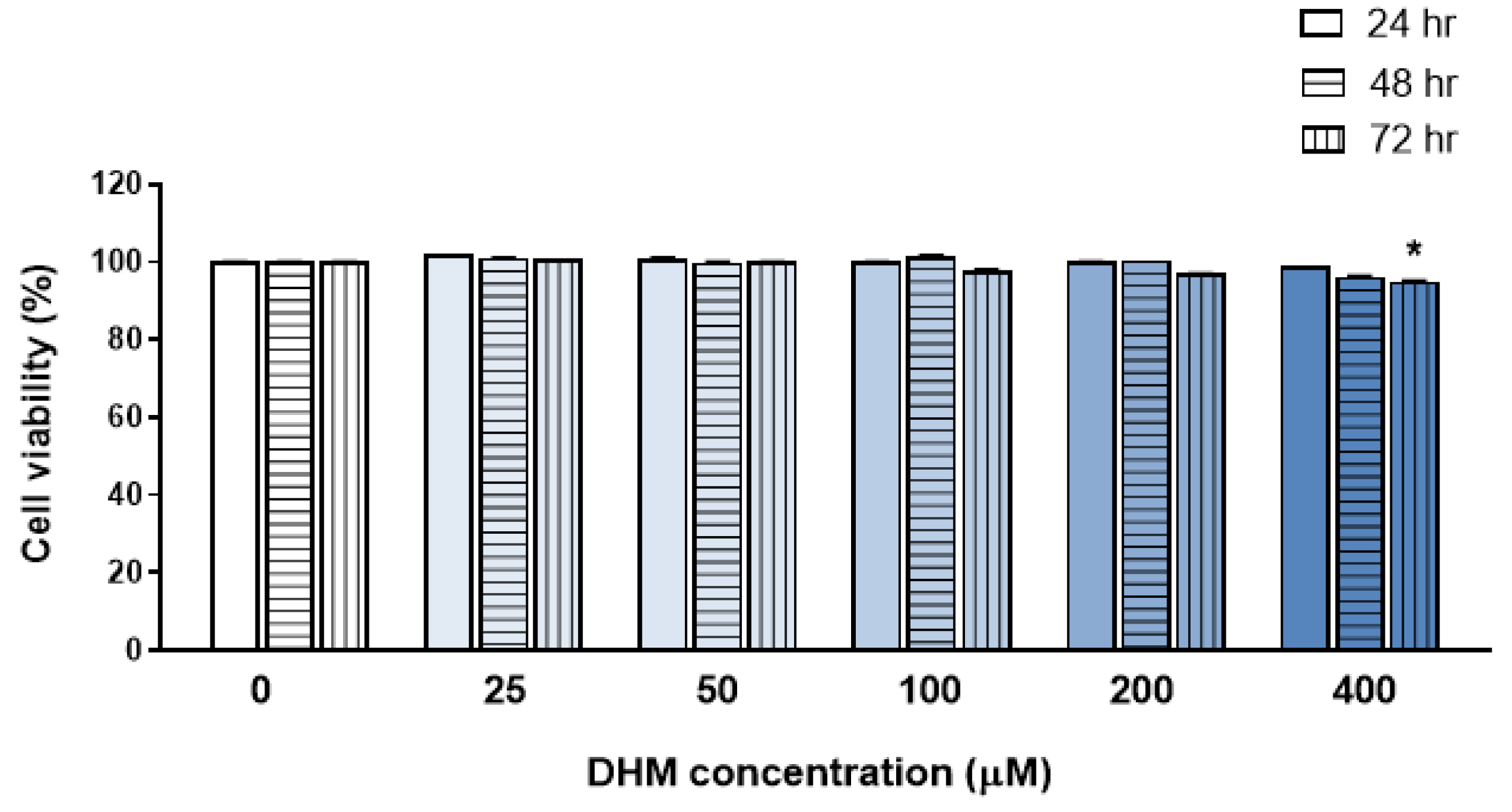
2.1. DHM Did Not Affect the Viability of HaCaT Cells at Concentrations up to 200 μM
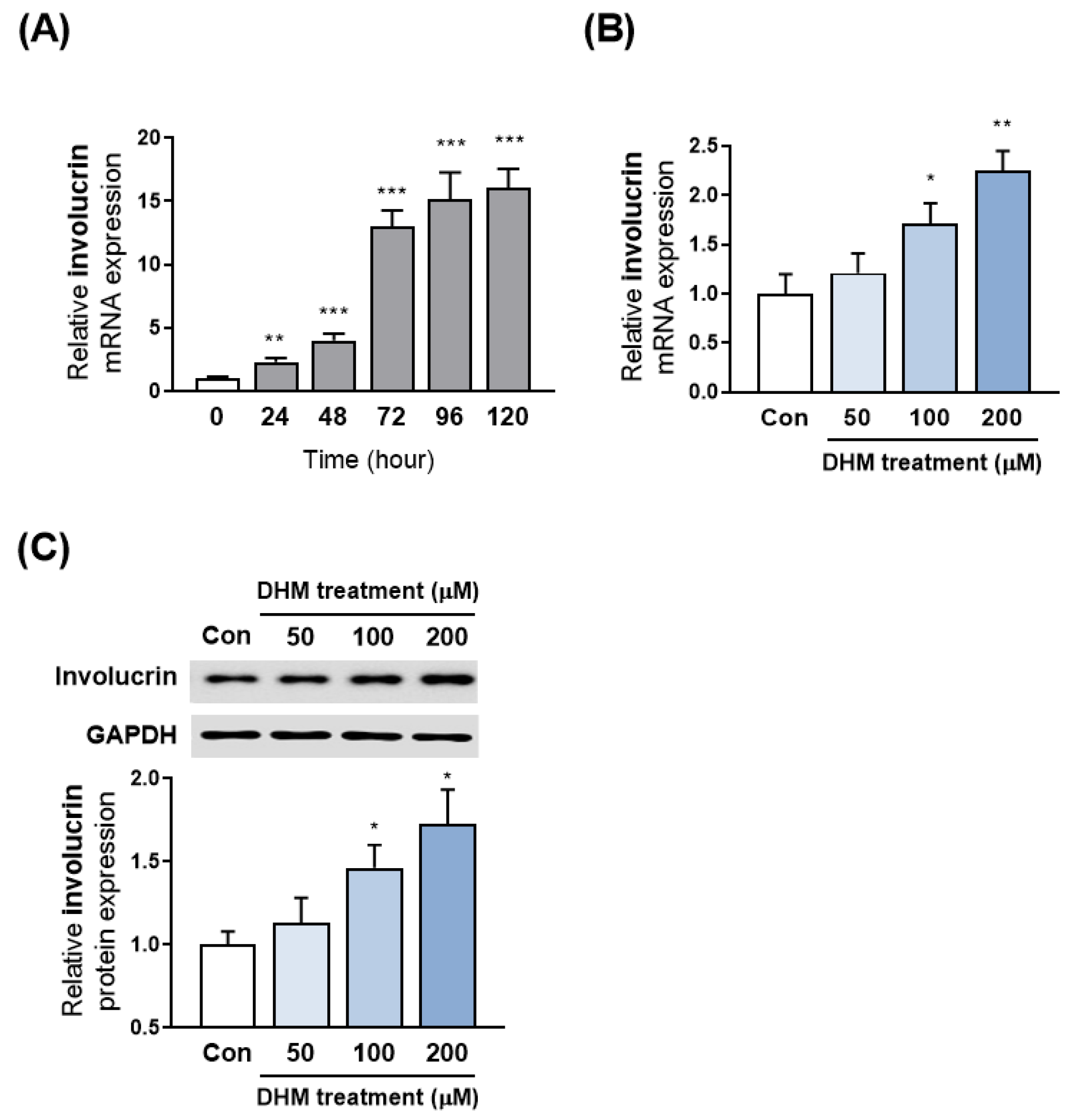
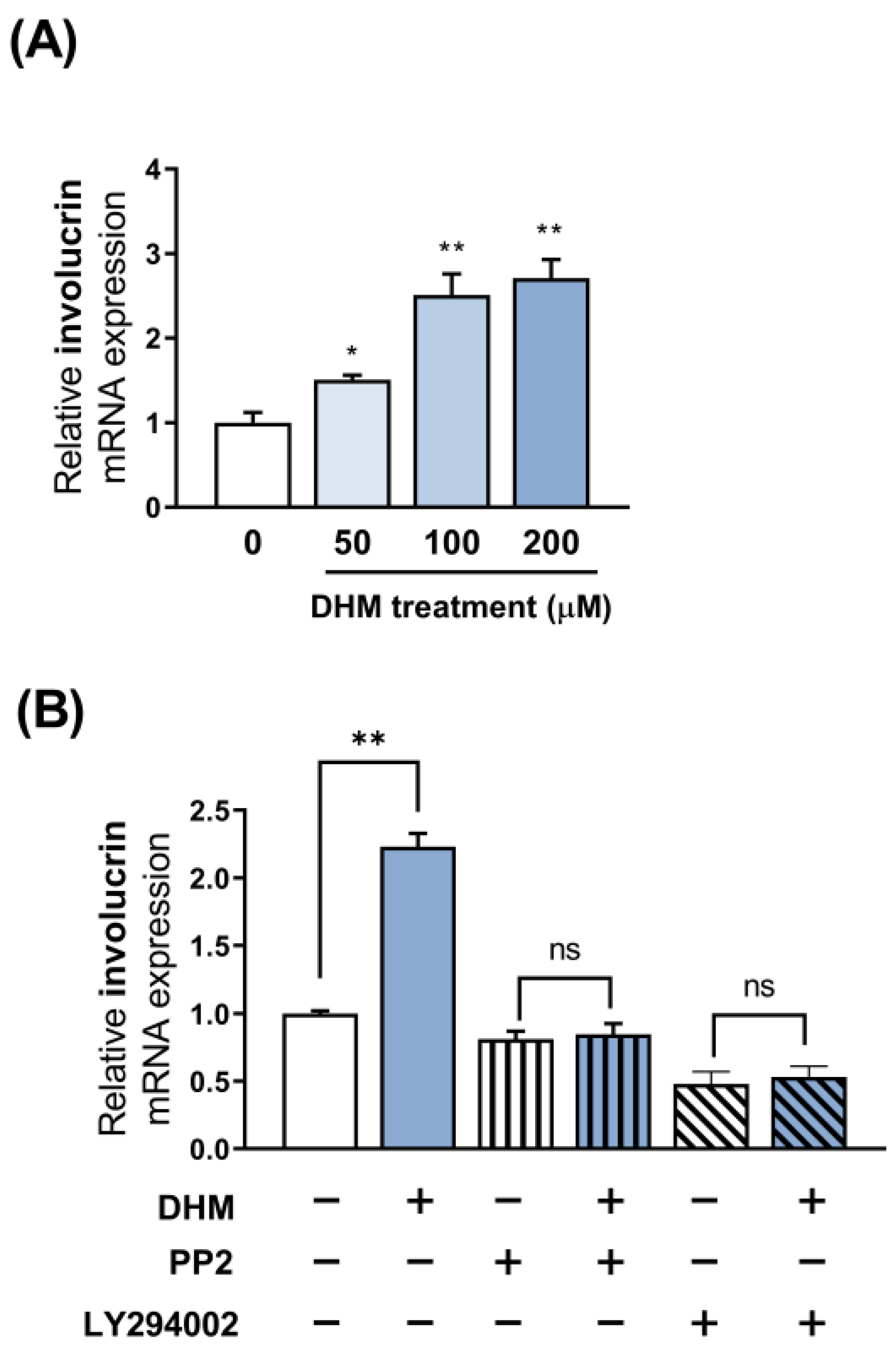
2.2. DHM Increased Involucrin Expression in HaCaT Cells
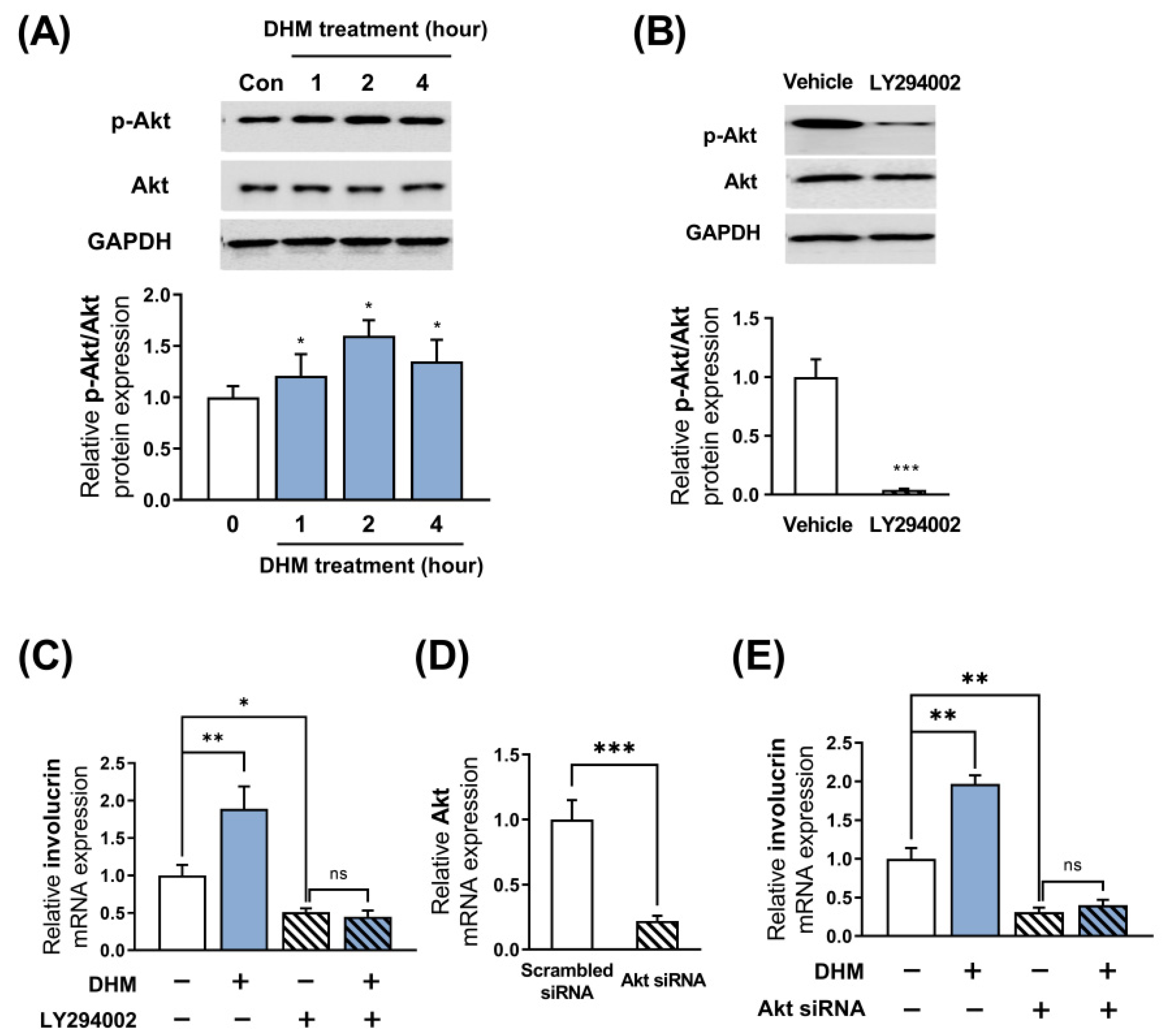
2.3. Akt Is Required for the DHM-Mediated Induction of Involucrin in HaCaT Cells
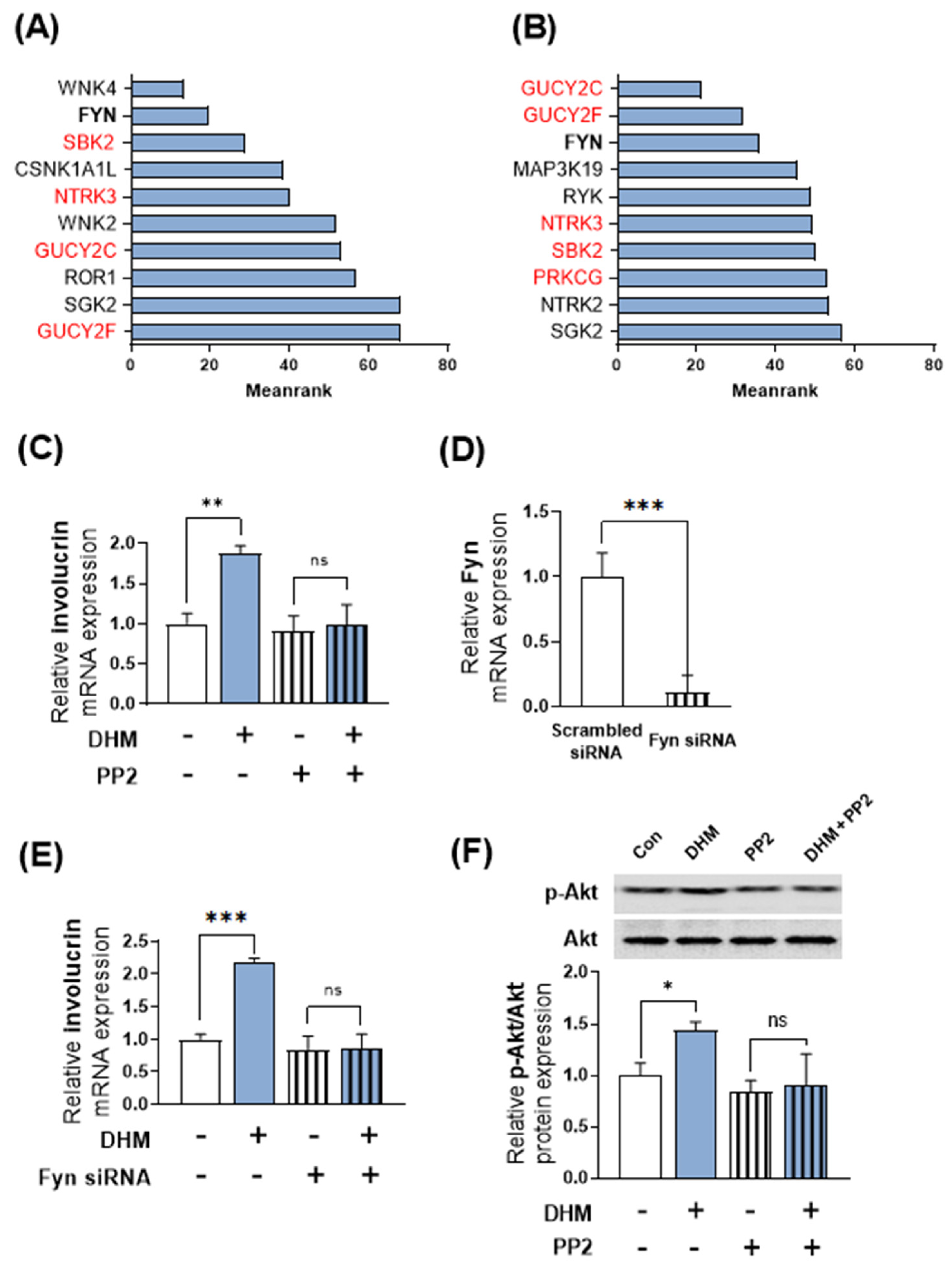
2.4. Fyn Serves as a Potential Upstream Regulator of DHM-Induced Akt Phosphorylation and Involucrin Expression in HaCaT Cells
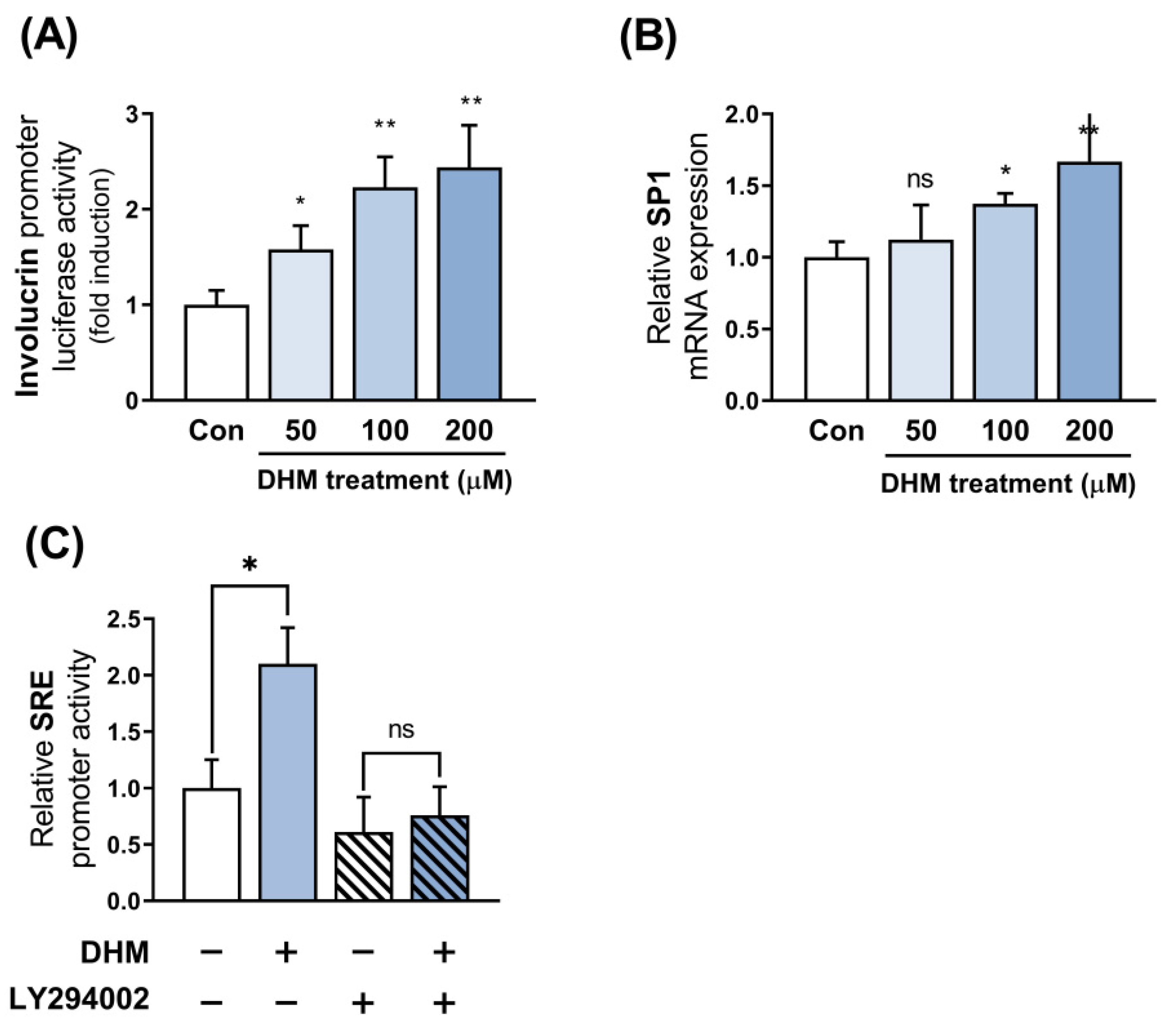
2.5. Sp1 Is Implicated as the Downstream Effector of Akt in Response to DHM
2.6. DHM Enhances Involucrin Expression in Primary Keratinocytes via the Fyn-Akt Pathway
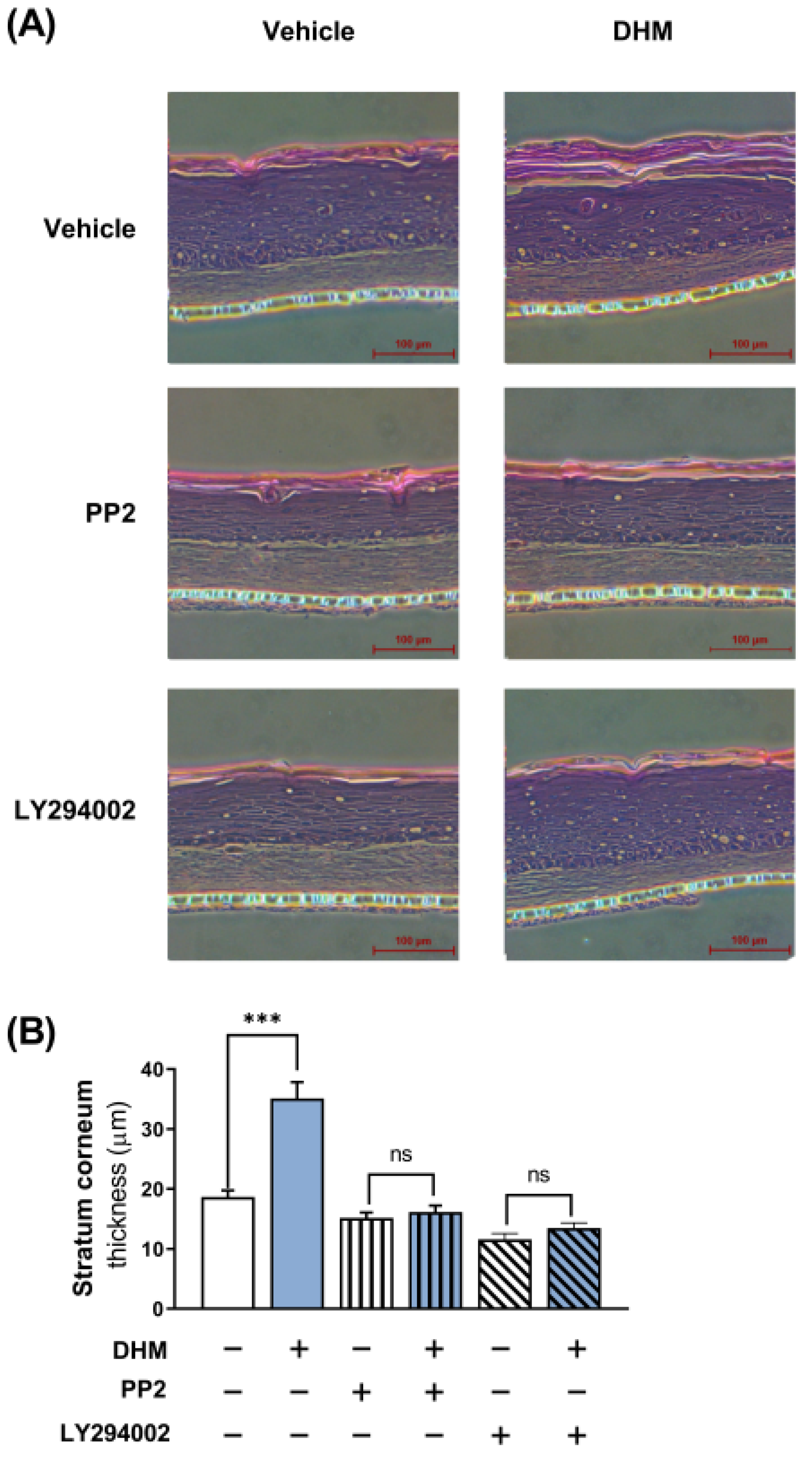
2.7. DHM Promoted Barrier Formation through the Fyn-Akt Pathway in a 3D Human Skin Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Assay
4.3. Transfection and Luciferase Assay
4.4. RT-qPCR
4.5. Western Blot
4.6. siRNA
4.7. RNA Sequencing and Bioinformatics
4.8. Reconstructed Human Skin Model
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef]
- Lim, K.-M. Skin epidermis and barrier function. Int. J. Mol. Sci. 2021, 22, 3035. [Google Scholar] [CrossRef]
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Brandner, J.M.; Jensen, J.M. The skin: An indispensable barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.A.; Irvine, A.D. The multifunctional role of filaggrin in allergic skin disease. J. Allergy Clin. Immunol. 2013, 131, 280–291. [Google Scholar] [CrossRef]
- Razia, S.; Park, H.; Shin, E.; Shim, K.-S.; Cho, E.; Kim, S.-Y. Effects of Aloe vera flower extract and its active constituent isoorientin on skin moisturization via regulating involucrin expression: In vitro and molecular docking studies. Molecules 2021, 26, 2626. [Google Scholar] [CrossRef]
- Gueniche, A.; Valois, A.; Kerob, D.; Rasmont, V.; Nielsen, M. A combination of Vitreoscilla filiformis extract and Vichy volcanic mineralizing water strengthens the skin defenses and skin barrier. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 16–25. [Google Scholar] [CrossRef]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, H.-S.; Jeon, Y.-J.; Jee, Y.; Kim, K.-N.; Lee, K.; Fernando, I.P.S.; Ahn, G. Sargassum horneri (Turner) C. Agardh ethanol extract attenuates fine dust-induced inflammatory responses and impaired skin barrier functions in HaCaT keratinocytes. J. Ethnopharmacol. 2021, 273, 114003. [Google Scholar] [CrossRef]
- Bhaskar, P.T.; Hay, N. The two TORCs and AKT. Dev. Cell 2007, 12, 487–502. [Google Scholar] [CrossRef]
- Martelli, A.M.; Tabellini, G.; Bressanin, D.; Ognibene, A.; Goto, K.; Cocco, L.; Evangelisti, C. The emerging multiple roles of nuclear Akt. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2012, 1823, 2168–2178. [Google Scholar] [CrossRef]
- O’Shaughnessy, R.F.; Welti, J.C.; Cooke, J.C.; Avilion, A.A.; Monks, B.; Birnbaum, M.J.; Byrne, C. AKT-dependent HspB1 (Hsp27) activity in epidermal differentiation. J. Biol. Chem. 2007, 282, 17297–17305. [Google Scholar] [CrossRef] [PubMed]
- Sully, K.; Akinduro, O.; Philpott, M.; Naeem, A.; Harwood, C.; Reeve, V.; O’shaughnessy, R.; Byrne, C. The mTOR inhibitor rapamycin opposes carcinogenic changes to epidermal Akt1/PKBα isoform signaling. Oncogene 2013, 32, 3254–3262. [Google Scholar] [CrossRef]
- O’Shaughnessy, R.F.; Welti, J.C.; Sully, K.; Byrne, C. Akt-dependent Pp2a activity is required for epidermal barrier formation during late embryonic development. Development 2009, 136, 3423–3431. [Google Scholar] [CrossRef]
- Kim, I.W.; Jeong, H.S.; Kwon, N.S.; Baek, K.J.; Yun, H.Y.; Kim, D.S. LGI 3 promotes human keratinocyte differentiation via the Akt pathway. Exp. Dermatol. 2018, 27, 1224–1229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ishikawa, M.; Doyle, A.; Nakamura, T.; He, B.; Yamada, Y. Pannexin 3 regulates skin development via Epiprofin. Sci. Rep. 2021, 11, 1779. [Google Scholar] [CrossRef]
- Sevilla, L.M.; Nachat, R.; Groot, K.R.; Klement, J.F.; Uitto, J.; Djian, P.; Määttä, A.; Watt, F.M. Mice deficient in involucrin, envoplakin, and periplakin have a defective epidermal barrier. J. Cell Biol. 2007, 179, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake-Mudiyanselage, S.; Jensen, J.-M.; Proksch, E.; Aschauer, H.; Schmook, F.P.; Meingassner, J.G. Expression of epidermal keratins and the cornified envelope protein involucrin is influenced by permeability barrier disruption. J. Investig. Dermatol. 1998, 111, 517–523. [Google Scholar] [CrossRef]
- Proksch, E.; Fölster-Holst, R.; Jensen, J.-M. Skin barrier function, epidermal proliferation and differentiation in eczema. J. Dermatol. Sci. 2006, 43, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-E.; Kook, J.-K.; Min, B.-M. βig-h3 induces keratinocyte differentiation via modulation of involucrin and transglutaminase expression through the integrin α3β1 and the phosphatidylinositol 3-kinase/Akt signaling pathway. J. Biol. Chem. 2005, 280, 21629–21637. [Google Scholar] [CrossRef]
- Tseng, Y.-H.; Chang, C.-S.; Liu, T.-Y.; Kao, S.-Y.; Chang, K.-W.; Lin, S.-C. Areca nut extract treatment down-regulates involucrin in normal human oral keratinocyte through P13K/AKT activation. Oral Oncol. 2007, 43, 670–679. [Google Scholar] [CrossRef]
- Mercurio, L.; Albanesi, C.; Madonna, S. Recent updates on the involvement of PI3K/AKT/mTOR molecular cascade in the pathogenesis of hyperproliferative skin disorders. Front. Med. 2021, 8, 665647. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; He, Y.; Li, C.; Zhang, X.; Xu, H.; Wang, B. Nicastrin mutations in familial acne inversa impact keratinocyte proliferation and differentiation through the Notch and phosphoinositide 3-kinase/AKT signalling pathways. Br. J. Dermatol. 2016, 174, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Calautti, E.; Li, J.; Saoncella, S.; Brissette, J.L.; Goetinck, P.F. Phosphoinositide 3-kinase signaling to Akt promotes keratinocyte differentiation versus death. J. Biol. Chem. 2005, 280, 32856–32865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, X. The role of PI3K/AKT/FOXO signaling in psoriasis. Arch. Dermatol. Res. 2019, 311, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Rogerson, C.; O’Shaughnessy, R.F. Protein kinases involved in epidermal barrier formation: The AKT family and other animals. Exp. Dermatol. 2018, 27, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Hong, I.-K.; Byun, H.-J.; Lee, J.; Jin, Y.-J.; Wang, S.-J.; Jeoung, D.-I.; Kim, Y.-M.; Lee, H. The tetraspanin CD81 protein increases melanoma cell motility by up-regulating metalloproteinase MT1-MMP expression through the pro-oncogenic Akt-dependent Sp1 activation signaling pathways. J. Biol. Chem. 2014, 289, 15691–15704. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-N.; Tsai, C.-L.; Yi, J.-S.; Kao, H.-K.; Huang, Y.; Wang, C.-I.; Lee, Y.-S.; Chang, K.-P. Activin A regulates the epidermal growth factor receptor promoter by activating the PI3K/SP1 pathway in oral squamous cell carcinoma cells. Sci. Rep. 2019, 9, 5197. [Google Scholar] [CrossRef]
- Bos, J.D.; Meinardi, M.M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. Viewp. 2000, 9, 165–169. [Google Scholar] [CrossRef]
- Ly, P.T.; Stewart, C.; Pallen, C.J. PTPα is required for laminin-2-induced Fyn–Akt signaling to drive oligodendrocyte differentiation. J. Cell Sci. 2018, 131, jcs212076. [Google Scholar] [CrossRef]
- An, L.; Li, W.; Cheng, G. Fyn arrests swainsonine-induced apoptosis in 293T cells via Akt and its phosphorylation. Genet. Mol. Res. 2015, 14, 5304–5309. [Google Scholar] [CrossRef]
- Cabral-Dias, R.; Lucarelli, S.; Zak, K.; Rahmani, S.; Judge, G.; Abousawan, J.; DiGiovanni, L.F.; Vural, D.; Anderson, K.E.; Sugiyama, M.G. Fyn and TOM1L1 are recruited to clathrin-coated pits and regulate Akt signaling. J. Cell Biol. 2022, 221, e201808181. [Google Scholar] [CrossRef]
- Ninio-Many, L.; Grossman, H.; Shomron, N.; Chuderland, D.; Shalgi, R. microRNA-125a-3p reduces cell proliferation and migration by targeting Fyn. J. Cell Sci. 2013, 126, 2867–2876. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Thomas, S.M.; Rickles, R.J.; Taylor, J.A.; Brauer, A.W.; Seidel-Dugan, C.; Michael, W.M.; Dreyfuss, G.; Brugge, J.S. Identification of Src, Fyn, and Lyn SH3-binding proteins: Implications for a function of SH3 domains. Mol. Cell. Biol. 1994, 14, 4509–4521. [Google Scholar]
- Saito, Y.D.; Jensen, A.R.; Salgia, R.; Posadas, E.M. Fyn: A novel molecular target in cancer. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2010, 116, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Fyn, a Src family tyrosine kinase. Int. J. Biochem. Cell Biol. 1998, 30, 1159–1162. [Google Scholar] [CrossRef]
- He, Z.; Tang, F.; Ermakova, S.; Li, M.; Zhao, Q.; Cho, Y.Y.; Ma, W.Y.; Choi, H.S.; Bode, A.M.; Yang, C.S. Fyn is a novel target of (−)-epigallocatechin gallate in the inhibition of JB6 Cl41 cell transformation. Mol. Carcinog. Publ. Coop. Univ. Tex. MD Anderson Cancer Cent. 2008, 47, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Byun, S.; Kang, N.J.; Lim, S.H.; Heo, Y.-S.; Bode, A.M.; Bowden, G.T.; Lee, H.J.; Dong, Z. Myricetin suppresses UVB-induced skin cancer by targeting Fyn. Cancer Res. 2008, 68, 6021–6029. [Google Scholar] [CrossRef]
- Hwang, M.K.; Kang, N.J.; Heo, Y.-S.; Lee, K.W.; Lee, H.J. Fyn kinase is a direct molecular target of delphinidin for the inhibition of cyclooxygenase-2 expression induced by tumor necrosis factor-α. Biochem. Pharmacol. 2009, 77, 1213–1222. [Google Scholar] [CrossRef]
- Kang, N.J.; Lee, K.W.; Shin, B.J.; Jung, S.K.; Hwang, M.K.; Bode, A.M.; Heo, Y.-S.; Lee, H.J.; Dong, Z. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis 2009, 30, 321–330. [Google Scholar] [CrossRef]
- Shang, G.; Tang, X.; Gao, P.; Guo, F.; Liu, H.; Zhao, Z.; Chen, Q.; Jiang, T.; Zhang, N.; Li, H. Sulforaphane attenuation of experimental diabetic nephropathy involves GSK-3 beta/Fyn/Nrf2 signaling pathway. J. Nutr. Biochem. 2015, 26, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, G.; Crish, J.F.; Gopalakrishnan, R.; Bone, F.; Eckert, R.L. Involucrin expression in the corneal epithelium: An essential role for Sp1 transcription factors. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3109–3120. [Google Scholar] [CrossRef] [PubMed]
- Banks, E.B.; Crish, J.F.; Welter, J.F.; Eckert, R.L. Characterization of human involucrin promoter distal regulatory region transcriptional activator elements–a role for Sp1 and AP1 binding sites. Biochem. J. 1998, 331, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Eckert, R.L.; Crish, J.F.; Efimova, T.; Dashti, S.R.; Deucher, A.; Bone, F.; Adhikary, G.; Huang, G.; Gopalakrishnan, R.; Balasubramanian, S. Regulation of involucrin gene expression. J. Investig. Dermatol. 2004, 123, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Koch, P.J.; de Viragh, P.A.; Scharer, E.; Bundman, D.; Longley, M.A.; Bickenbach, J.; Kawachi, Y.; Suga, Y.; Zhou, Z.; Huber, M. Lessons from loricrin-deficient mice: Compensatory mechanisms maintaining skin barrier function in the absence of a major cornified envelope protein. J. Cell Biol. 2000, 151, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Mischke, D.; Korge, B.P.; Marenholz, I.; Volz, A.; Ziegler, A. Genes encoding structural proteins of epidermal cornification and S100 calcium-binding proteins form a gene complex (“epidermal differentiation complex”) on human chromosome 1q21. J. Investig. Dermatol. 1996, 106, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.M.; Albers, K.M.; Garlick, J.A.; Harrington, R.; Taichman, L.B. Tissue-and stratum-specific expression of the human involucrin promoter in transgenic mice. Proc. Natl. Acad. Sci. USA 1993, 90, 10270–10274. [Google Scholar] [CrossRef]
- Ng, D.C.; Shafaee, S.; Lee, D.; Bikle, D.D. Requirement of an AP-1 site in the calcium response region of the involucrin promoter. J. Biol. Chem. 2000, 275, 24080–24088. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Xie, Z.; London, A.B.; Yang, J.; Evangelista, J.E.; Lachmann, A.; Shu, I.; Torre, D.; Ma’ayan, A. KEA3: Improved kinase enrichment analysis via data integration. Nucleic Acids Res. 2021, 49, W304–W316. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Kang, W.; Choi, D.; Roh, J.; Park, T. Dihydromyrcenol Modulates Involucrin Expression through the Akt Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 2246. https://doi.org/10.3390/ijms25042246
Yang S, Kang W, Choi D, Roh J, Park T. Dihydromyrcenol Modulates Involucrin Expression through the Akt Signaling Pathway. International Journal of Molecular Sciences. 2024; 25(4):2246. https://doi.org/10.3390/ijms25042246
Chicago/Turabian StyleYang, Suhjin, Wesuk Kang, Dabin Choi, Jiyun Roh, and Taesun Park. 2024. "Dihydromyrcenol Modulates Involucrin Expression through the Akt Signaling Pathway" International Journal of Molecular Sciences 25, no. 4: 2246. https://doi.org/10.3390/ijms25042246