
Preconception Diet Interventions in Obese Outbred Mice and the Impact on Female Offspring Metabolic Health and Oocyte Quality
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Litter Characteristics
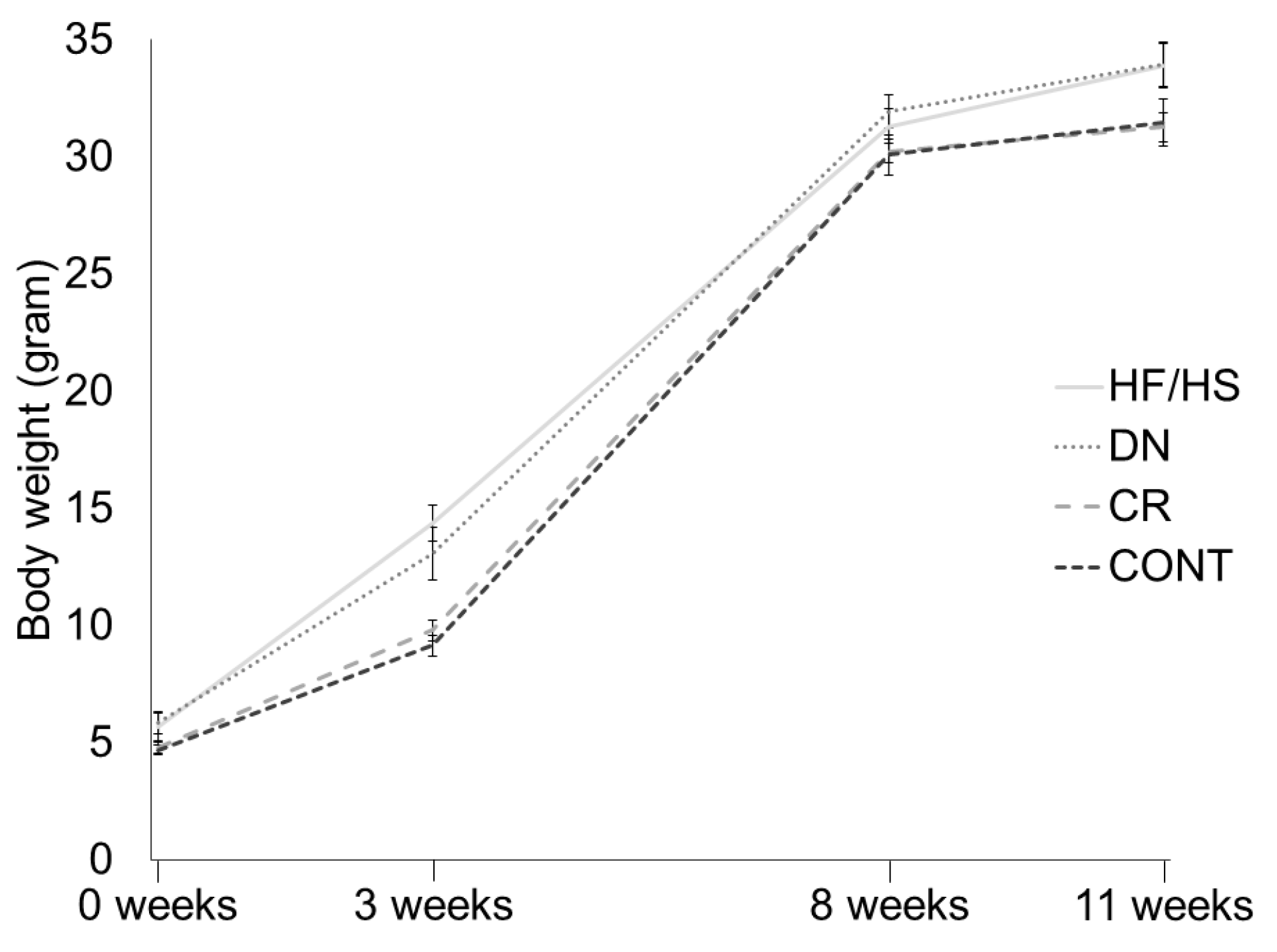
2.2. Offspring Body Weight
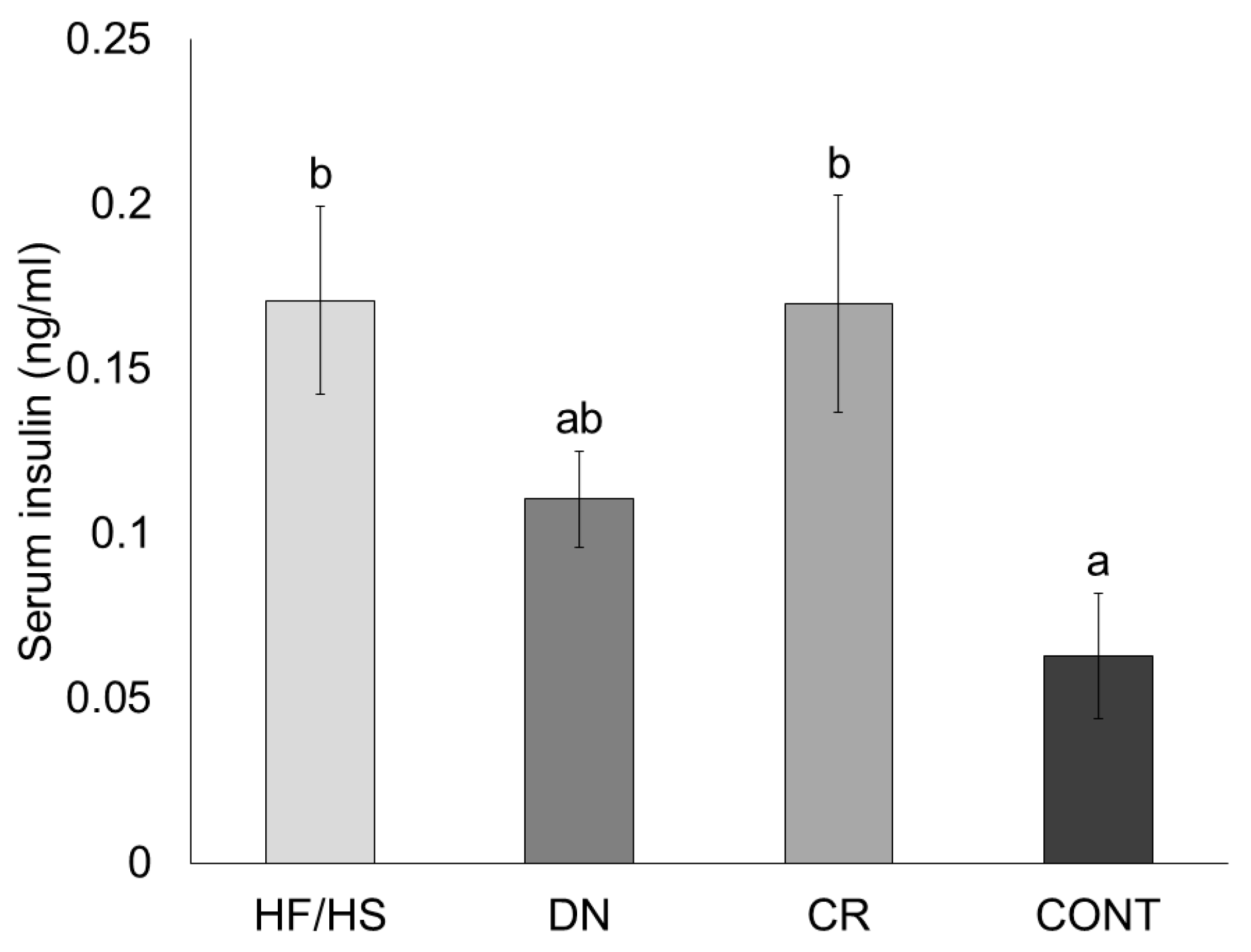
2.3. Blood Serum Profile
2.4. Oocyte Mitochondrial Ultrastructure
2.4.1. Mitochondrial Morphology
2.4.2. Mitochondrial Dimensions
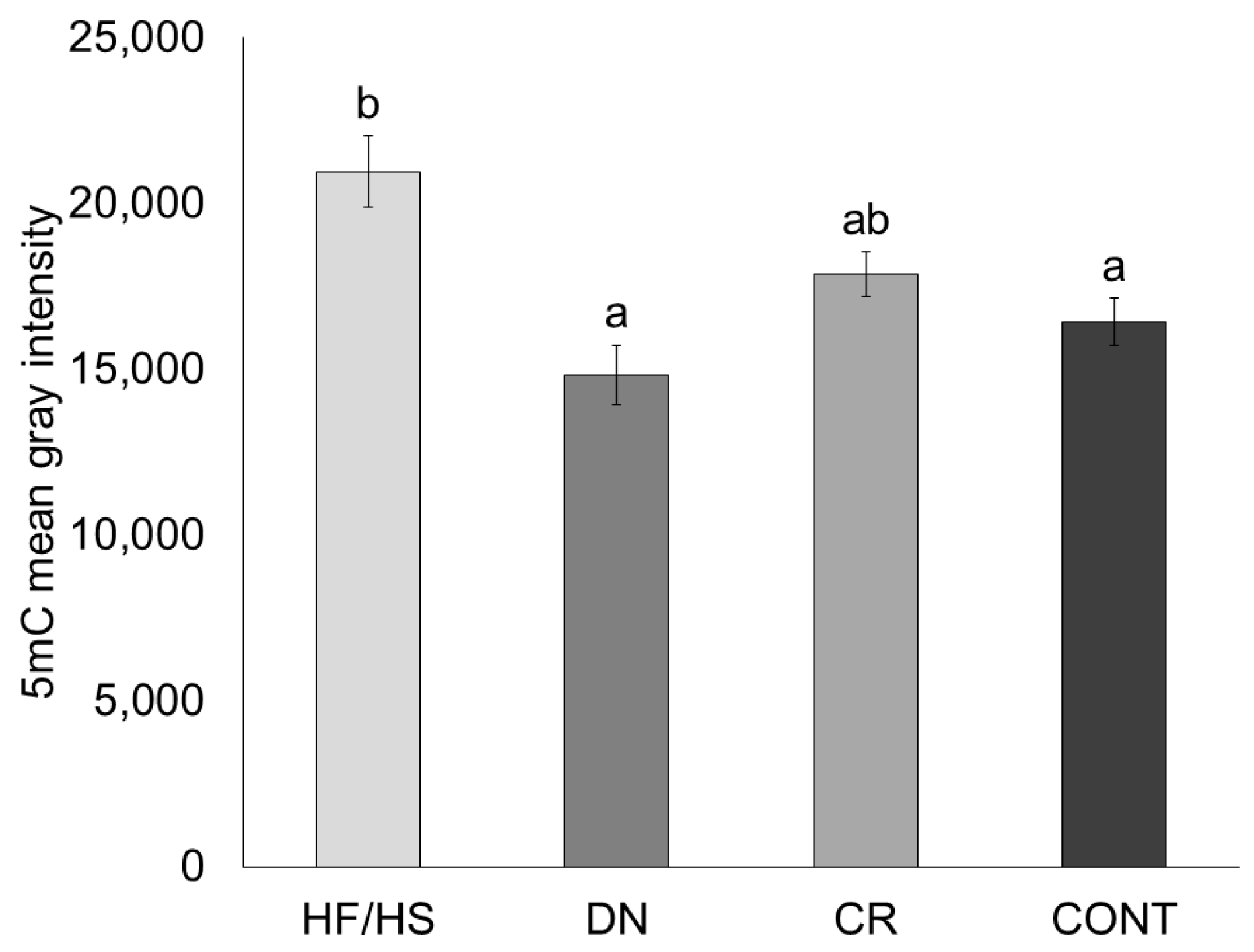
2.5. Oocyte Global DNA Methylation
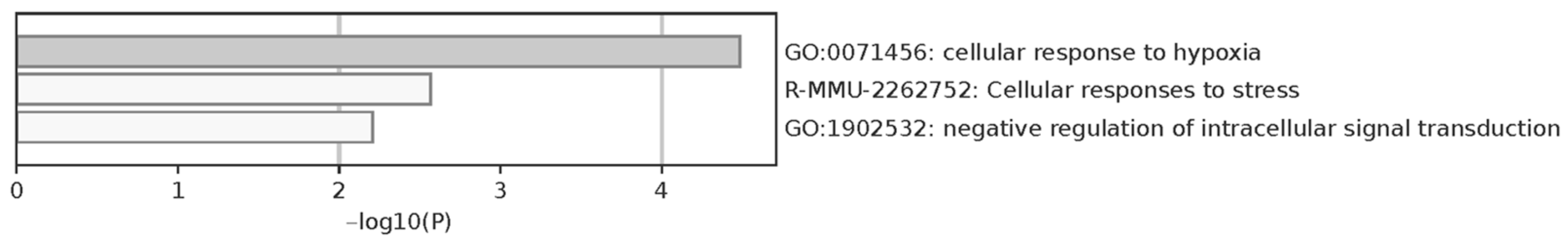
2.6. Oocyte Proteome Analysis
3. Discussion
4. Materials and Methods
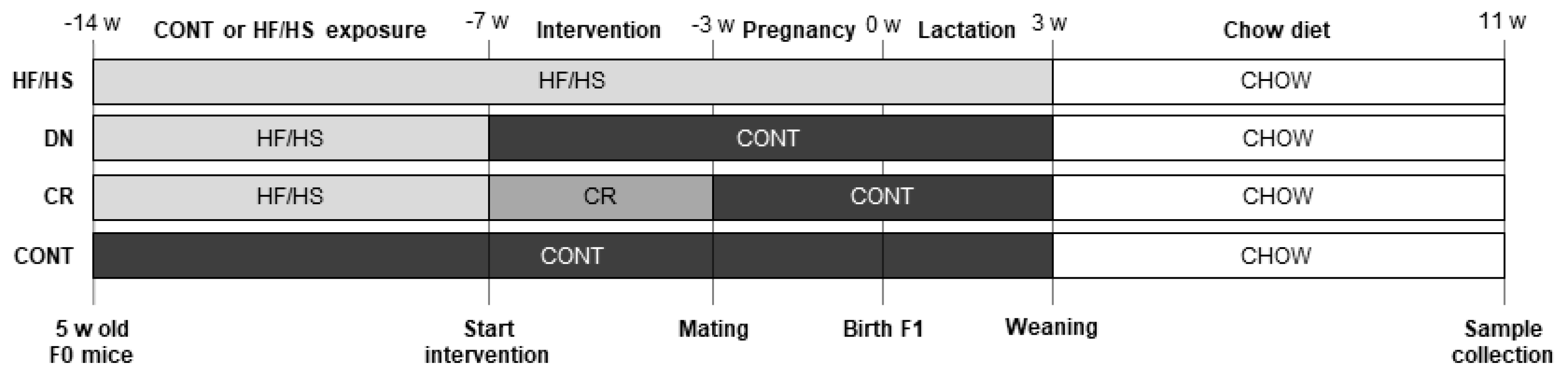
4.1. Animals, Diet, and Experimental Design
4.2. Serum Collection and Analysis
4.3. Oocyte Collection
4.4. Transmission Electron Microscopy (TEM)
4.5. 5-Methylcytosine Immunostaining
4.6. TIMS-TOF Shotgun Proteomics
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 17 November 2023).
- Bluher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Jin, X.; Qiu, T.; Li, L.; Yu, R.; Chen, X.; Li, C.; Proud, C.G.; Jiang, T. Pathophysiology of obesity and its associated diseases. Acta Pharm. Sin. B 2023, 13, 2403–2424. [Google Scholar] [CrossRef]
- Zain, M.M.; Norman, R.J. Impact of obesity on female fertility and fertility treatment. Womens Health 2008, 4, 183–194. [Google Scholar] [CrossRef]
- Bellver, J.; Busso, C.; Pellicer, A.; Remohi, J.; Simon, C. Obesity and assisted reproductive technology outcomes. Reprod. Biomed. Online 2006, 12, 562–568. [Google Scholar] [CrossRef]
- Luke, B.; Brown, M.B.; Stern, J.E.; Missmer, S.A.; Fujimoto, V.Y.; Leach, R.; Group, S.W. Female obesity adversely affects assisted reproductive technology (ART) pregnancy and live birth rates. Hum. Reprod. 2011, 26, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Valckx, S.D.; Arias-Alvarez, M.; De Pauw, I.; Fievez, V.; Vlaeminck, B.; Fransen, E.; Bols, P.E.; Leroy, J.L. Fatty acid composition of the follicular fluid of normal weight, overweight and obese women undergoing assisted reproductive treatment: A descriptive cross-sectional study. Reprod. Biol. Endocrinol. 2014, 12, 13. [Google Scholar] [CrossRef]
- Marei, W.F.A.; Smits, A.; Mohey-Elsaeed, O.; Pintelon, I.; Ginneberge, D.; Bols, P.E.J.; Moerloose, K.; Leroy, J. Differential effects of high fat diet-induced obesity on oocyte mitochondrial functions in inbred and outbred mice. Sci. Rep. 2020, 10, 9806. [Google Scholar] [CrossRef]
- Grindler, N.M.; Moley, K.H. Maternal obesity, infertility and mitochondrial dysfunction: Potential mechanisms emerging from mouse model systems. Mol. Hum. Reprod. 2013, 19, 486–494. [Google Scholar] [CrossRef]
- Igosheva, N.; Abramov, A.Y.; Poston, L.; Eckert, J.J.; Fleming, T.P.; Duchen, M.R.; McConnell, J. Maternal diet-induced obesity alters mitochondrial activity and redox status in mouse oocytes and zygotes. PLoS ONE 2010, 5, e10074. [Google Scholar] [CrossRef] [PubMed]
- Meulders, B.; Leroy, J.; Xhonneux, I.; Bols, P.E.J.; Marei, W.F.A. In vitro reduction of bovine oocyte ATP production with oligomycin affects embryo epigenome. Reproduction 2024, 167, e230271. [Google Scholar] [CrossRef]
- Messerschmidt, D.M.; Knowles, B.B.; Solter, D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev. 2014, 28, 812–828. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, T. DNA Methylation Reprogramming during Mammalian Development. Genes 2019, 10, 257. [Google Scholar] [CrossRef]
- Breton-Larrivee, M.; Elder, E.; McGraw, S. DNA methylation, environmental exposures and early embryo development. Anim. Reprod. 2019, 16, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.-J.; Zhu, C.-C.; Duan, X.; Liu, H.-L.; Wang, Q.; Sun, S.-C. Both diet and gene mutation induced obesity affect oocyte quality in mice. Sci. Rep. 2016, 6, 18858. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.-J.; Luo, S.-M.; Lin, F.; Liang, Q.-X.; Huang, L.; Wei, Y.-C.; Hou, Y.; Han, Z.-M.; Schatten, H.; Sun, Q.-Y. DNA methylation in oocytes and liver of female mice and their offspring: Effects of high-fat-diet-induced obesity. Environ. Health Perspect. 2014, 122, 159–164. [Google Scholar] [CrossRef]
- Marei, W.F.A.; Van Raemdonck, G.; Baggerman, G.; Bols, P.E.J.; Leroy, J. Proteomic changes in oocytes after in vitro maturation in lipotoxic conditions are different from those in cumulus cells. Sci. Rep. 2019, 9, 3673. [Google Scholar] [CrossRef]
- Nicholas, L.M.; Ozanne, S.E. Early life programming in mice by maternal overnutrition: Mechanistic insights and interventional approaches. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180116. [Google Scholar] [CrossRef]
- Samuelsson, A.M.; Matthews, P.A.; Argenton, M.; Christie, M.R.; McConnell, J.M.; Jansen, E.H.; Piersma, A.H.; Ozanne, S.E.; Twinn, D.F.; Remacle, C.; et al. Diet-induced obesity in female mice leads to offspring hyperphagia, adiposity, hypertension, and insulin resistance: A novel murine model of developmental programming. Hypertension 2008, 51, 383–392. [Google Scholar] [CrossRef]
- Volpato, A.M.; Schultz, A.; Magalhaes-da-Costa, E.; Correia, M.L.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Maternal high-fat diet programs for metabolic disturbances in offspring despite leptin sensitivity. Neuroendocrinology 2012, 96, 272–284. [Google Scholar] [CrossRef]
- Elahi, M.M.; Cagampang, F.R.; Mukhtar, D.; Anthony, F.W.; Ohri, S.K.; Hanson, M.A. Long-term maternal high-fat feeding from weaning through pregnancy and lactation predisposes offspring to hypertension, raised plasma lipids and fatty liver in mice. Br. J. Nutr. 2009, 102, 514–519. [Google Scholar] [CrossRef]
- Fleming, T.P.; Eckert, J.J.; Denisenko, O. The Role of Maternal Nutrition During the Periconceptional Period and Its Effect on Offspring Phenotype. Adv. Exp. Med. Biol. 2017, 1014, 87–105. [Google Scholar] [CrossRef]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; et al. Origins of lifetime health around the time of conception: Causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef]
- Steegers-Theunissen, R.P.; Twigt, J.; Pestinger, V.; Sinclair, K.D. The periconceptional period, reproduction and long-term health of offspring: The importance of one-carbon metabolism. Hum. Reprod. Update 2013, 19, 640–655. [Google Scholar] [CrossRef]
- Velazquez, M.A.; Fleming, T.P.; Watkins, A.J. Periconceptional environment and the developmental origins of disease. J. Endocrinol. 2019, 242, T33–T49. [Google Scholar] [CrossRef]
- Saben, J.L.; Boudoures, A.L.; Asghar, Z.; Thompson, A.; Drury, A.; Zhang, W.; Chi, M.; Cusumano, A.; Scheaffer, S.; Moley, K.H. Maternal Metabolic Syndrome Programs Mitochondrial Dysfunction via Germline Changes across Three Generations. Cell Rep. 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Andreas, E.; Reid, M.; Zhang, W.; Moley, K.H. The effect of maternal high-fat/high-sugar diet on offspring oocytes and early embryo development. Mol. Hum. Reprod. 2019, 25, 717–728. [Google Scholar] [CrossRef]
- Jungheim, E.S.; Moley, K.H. Current knowledge of obesity’s effects in the pre- and periconceptional periods and avenues for future research. Am. J. Obstet. Gynecol. 2010, 203, 525–530. [Google Scholar] [CrossRef]
- Lassi, Z.S.; Dean, S.V.; Mallick, D.; Bhutta, Z.A. Preconception care: Delivery strategies and packages for care. Reprod. Health 2014, 11 (Suppl. S3), S7. [Google Scholar] [CrossRef]
- Smits, A.; Marei, W.F.A.; De Neubourg, D.; Leroy, J. Diet normalization or caloric restriction as a preconception care strategy to improve metabolic health and oocyte quality in obese outbred mice. Reprod. Biol. Endocrinol. 2021, 19, 166. [Google Scholar] [CrossRef]
- Smits, A.; Marei, W.F.A.; Moorkens, K.; Bols, P.E.J.; De Neubourg, D.; Leroy, J. Obese outbred mice only partially benefit from diet normalization or calorie restriction as preconception care interventions to improve metabolic health and oocyte quality. Hum. Reprod. 2022, 37, 2867–2884. [Google Scholar] [CrossRef]
- den Harink, T.; Blom, N.A.; Gemke, R.; Groen, H.; Hoek, A.; Mol, B.W.J.; Painter, R.C.; Kuipers, I.M.; Roseboom, T.J.; van Deutekom, A.W. Preconception lifestyle intervention in women with obesity and echocardiographic indices of cardiovascular health in their children. Int. J. Obes. 2022, 46, 1262–1270. [Google Scholar] [CrossRef]
- Liu, Z.; Ding, Z.; Lynch, E.C.; McCauley, N.; Zhou, Y.; Zhang, K.K.; Xie, L. Pregestational diet transition to normal-fat diet avoids the deterioration of pancreatic beta-cell function in male offspring induced by maternal high-fat diet. J. Nutr. Biochem. 2020, 86, 108495. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, H.; Xu, H.; Li, J.; Golovko, M.; Cheng, H.; Lynch, E.C.; Liu, L.; McCauley, N.; Kennedy, L.; et al. Maternal diet intervention before pregnancy primes offspring lipid metabolism in liver. Lab. Investig. 2020, 100, 553–569. [Google Scholar] [CrossRef]
- Summerfield, M.; Zhou, Y.; Zhou, T.; Wu, C.; Alpini, G.; Zhang, K.K.; Xie, L. A long-term maternal diet transition from high-fat diet to normal fat diet during pre-pregnancy avoids adipose tissue inflammation in next generation. PLoS ONE 2018, 13, e0209053. [Google Scholar] [CrossRef]
- Panchenko, P.E.; Lacroix, M.C.; Jouin, M.; Voisin, S.; Badonnel, K.; Lemaire, M.; Meunier, N.; Safi-Stibler, S.; Persuy, M.A.; Jouneau, L.; et al. Effect of Maternal Obesity and Preconceptional Weight Loss on Male and Female Offspring Metabolism and Olfactory Performance in Mice. Nutrients 2019, 11, 948. [Google Scholar] [CrossRef]
- Zhu, Z.; Cao, F.; Li, X. Epigenetic Programming and Fetal Metabolic Programming. Front. Endocrinol. 2019, 10, 764. [Google Scholar] [CrossRef]
- Xhonneux, I.; Marei, W.F.A.; Meulders, B.; Andries, S.; Leroy, J. The impact of a maternal and offspring obesogenic diet on daughter’s oocyte mitochondrial ultrastructure and bioenergetic responses. Insights from an outbred mouse model. Front. Physiol. 2023, 14, 1288472. [Google Scholar] [CrossRef]
- Boudoures, A.L.; Saben, J.; Drury, A.; Scheaffer, S.; Modi, Z.; Zhang, W.; Moley, K.H. Obesity-exposed oocytes accumulate and transmit damaged mitochondria due to an inability to activate mitophagy. Dev. Biol. 2017, 426, 126–138. [Google Scholar] [CrossRef]
- Elias-Lopez, A.L.; Vazquez-Mena, O.; Sferruzzi-Perri, A.N. Mitochondrial dysfunction in the offspring of obese mothers and it’s transmission through damaged oocyte mitochondria: Integration of mechanisms. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166802. [Google Scholar] [CrossRef]
- Halestrap, A.P. The regulation of the matrix volume of mammalian mitochondria in vivo and in vitro and its role in the control of mitochondrial metabolism. Biochim. Biophys. Acta 1989, 973, 355–382. [Google Scholar] [CrossRef]
- Luzzo, K.M.; Wang, Q.; Purcell, S.H.; Chi, M.; Jimenez, P.T.; Grindler, N.; Schedl, T.; Moley, K.H. High fat diet induced developmental defects in the mouse: Oocyte meiotic aneuploidy and fetal growth retardation/brain defects. PLoS ONE 2012, 7, e49217. [Google Scholar] [CrossRef]
- Runkel, E.D.; Baumeister, R.; Schulze, E. Mitochondrial stress: Balancing friend and foe. Exp. Gerontol. 2014, 56, 194–201. [Google Scholar] [CrossRef]
- Brand, M.D. Uncoupling to survive? The role of mitochondrial inefficiency in ageing. Exp. Gerontol. 2000, 35, 811–820. [Google Scholar] [CrossRef]
- Seli, E.; Wang, T.; Horvath, T.L. Mitochondrial unfolded protein response: A stress response with implications for fertility and reproductive aging. Fertil. Steril. 2019, 111, 197–204. [Google Scholar] [CrossRef]
- Barlow, D.P.; Bartolomei, M.S. Genomic imprinting in mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a018382. [Google Scholar] [CrossRef]
- O’Doherty, A.M.; O’Gorman, A.; Al Naib, A.; Brennan, L.; Daly, E.; Duffy, P.; Fair, T. Negative energy balance affects imprint stability in oocytes recovered from postpartum dairy cows. Genomics 2014, 104, 177–185. [Google Scholar] [CrossRef]
- Temple, I.K.; Shield, J.P. Transient neonatal diabetes, a disorder of imprinting. J. Med. Genet. 2002, 39, 872–875. [Google Scholar] [CrossRef]
- Pan, Z.; Zhang, J.; Li, Q.; Li, Y.; Shi, F.; Xie, Z.; Liu, H. Current advances in epigenetic modification and alteration during mammalian ovarian folliculogenesis. J. Genet. Genomics 2012, 39, 111–123. [Google Scholar] [CrossRef]
- Hackett, J.A.; Surani, M.A. DNA methylation dynamics during the mammalian life cycle. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20110328. [Google Scholar] [CrossRef]
- Miura, Y.; Endo, T. Survival responses to oxidative stress and aging. Geriatr. Gerontol. Int. 2010, 10 (Suppl. S1), S1–S9. [Google Scholar] [CrossRef]
- Marei, W.F.A.; Leroy, J. Cellular Stress Responses in Oocytes: Molecular Changes and Clinical Implications. Adv. Exp. Med. Biol. 2022, 1387, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.J.; Liang, X.W.; Guo, L.; Liang, Q.X.; Luo, S.M.; Wang, Y.P.; Wei, Y.C.; Han, Z.M.; Schatten, H.; Sun, Q.Y. Maternal diabetes causes alterations of DNA methylation statuses of some imprinted genes in murine oocytes. Biol. Reprod. 2013, 88, 117. [Google Scholar] [CrossRef] [PubMed]
- Aksungar, F.B.; Sarikaya, M.; Coskun, A.; Serteser, M.; Unsal, I. Comparison of Intermittent Fasting Versus Caloric Restriction in Obese Subjects: A Two Year Follow-Up. J. Nutr. Health Aging 2017, 21, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Boudoures, A.L.; Chi, M.; Thompson, A.; Zhang, W.; Moley, K.H. The effects of voluntary exercise on oocyte quality in a diet-induced obese murine model. Reproduction 2016, 151, 261–270. [Google Scholar] [CrossRef]
- Dobbs, K.B.; Rodriguez, M.; Sudano, M.J.; Ortega, M.S.; Hansen, P.J. Dynamics of DNA methylation during early development of the preimplantation bovine embryo. PloS ONE 2013, 8, e66230. [Google Scholar] [CrossRef]
- Wu, X.; Hu, S.; Wang, L.; Li, Y.; Yu, H. Dynamic changes of histone acetylation and methylation in bovine oocytes, zygotes, and preimplantation embryos. J. Exp. Zool. B Mol. Dev. Evol. 2020, 334, 245–256. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HF/HS | DN | CR | CONT | |
|---|---|---|---|---|
| Litter size (n) | 12.5 ± 2.1 | 10.3 ± 1.7 | 16.5 ± 0.3 | 15.5 ± 0.9 |
| Female/male (%) | 60.3 ± 2.3% | 57.8 ± 9.8% | 56.7 ± 10.8% | 53.4 ± 3.9% |
| HF/HS | DN | CR | CONT | |
|---|---|---|---|---|
| Total | 1603 | 1414 | 774 | 1172 |
| Normal | 1399 (87.3%) b | 1232 (87.1%) b | 636 (82.2%) a | 1061 (90.5%) c |
| Spherical homogenous | 828 (51.7%) a | 830 (58.7%) c | 415 (53.6%) b | 668 (57.0%) c |
| Regular vacuoles | 572 (35.7%) b | 402 (28.4%) a | 221 (28.6%) a | 393 (33.5%) b |
| Abnormal | 204 (12.7%) b | 182 (12.9%) b | 138 (17.8%) c | 111 (9.5%) a |
| Spherical loose IM | 79 (4.9%) a,b | 77 (5.5%) b,c | 58 (7.5%) c | 41 (3.5%) a |
| Non-spherical loose IM | 8 (0.5%) | 1 (0.1%) | 0 (0%) | 3 (0.3%) |
| Dumbbell | 13 (0.8%) | 8 (0.6%) | 6 (0.8%) | 5 (0.4%) |
| Elongation | 20 (1.3%) a | 26 (1.8%) a,b | 15 (1.9%) a,b | 33 (2.8%) b |
| Degeneration | 46 (2.9%) b | 46 (3.3%) b | 44 (5.7%) c | 15 (1.3%) a |
| Rose petal | 13 (0.8%) a | 19 (1.3%) a,b | 14 (1.8%) b | 9 (0.8%) a |
| Increased electron density | 28 (1.8%) b | 6 (0.4%) a | 2 (0.3%) a | 6 (0.5%) a |
| HF/HS | DN | CR | CONT | |
|---|---|---|---|---|
| Area (pixels2) | 0.104 ± 0.002 b | 0.099 ± 0.002 a | 0.098 ± 0.002 ab | 0.092 ± 0.001 a |
| Width (pixels) | 0.334 ± 0.003 bc | 0.327 ± 0.004 ab | 0.341 ± 0.004 c | 0.304 ± 0.003 a |
| Length (pixels) | 0.399 ± 0.004 c | 0.356 ± 0.004 a | 0.381 ± 0.005 b | 0.367 ± 0.004 ab |
| Roundness | 0.742 ± 0.004 a | 0.729 ± 0.005 a | 0.771 ± 0.007 b | 0.723 ± 0.005 a |
| Mean gray intensity | 187.2 ± 1.2 c | 174.7 ± 1.1 a | 184.1 ± 1.1 b | 182.1 ± 1.1 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meulders, B.; Marei, W.F.A.; Xhonneux, I.; Loier, L.; Smits, A.; Leroy, J.L.M.R. Preconception Diet Interventions in Obese Outbred Mice and the Impact on Female Offspring Metabolic Health and Oocyte Quality. Int. J. Mol. Sci. 2024, 25, 2236. https://doi.org/10.3390/ijms25042236
Meulders B, Marei WFA, Xhonneux I, Loier L, Smits A, Leroy JLMR. Preconception Diet Interventions in Obese Outbred Mice and the Impact on Female Offspring Metabolic Health and Oocyte Quality. International Journal of Molecular Sciences. 2024; 25(4):2236. https://doi.org/10.3390/ijms25042236
Chicago/Turabian StyleMeulders, Ben, Waleed F. A. Marei, Inne Xhonneux, Lien Loier, Anouk Smits, and Jo L. M. R. Leroy. 2024. "Preconception Diet Interventions in Obese Outbred Mice and the Impact on Female Offspring Metabolic Health and Oocyte Quality" International Journal of Molecular Sciences 25, no. 4: 2236. https://doi.org/10.3390/ijms25042236