
Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Follicular Growth and Oocyte Maturation
2.1. Follicular Growth
2.2. The Role of Granulosa Cells in Mature Oocytes
3. “Bidirectional Regulation” of Granulosa Cells and Oocytes
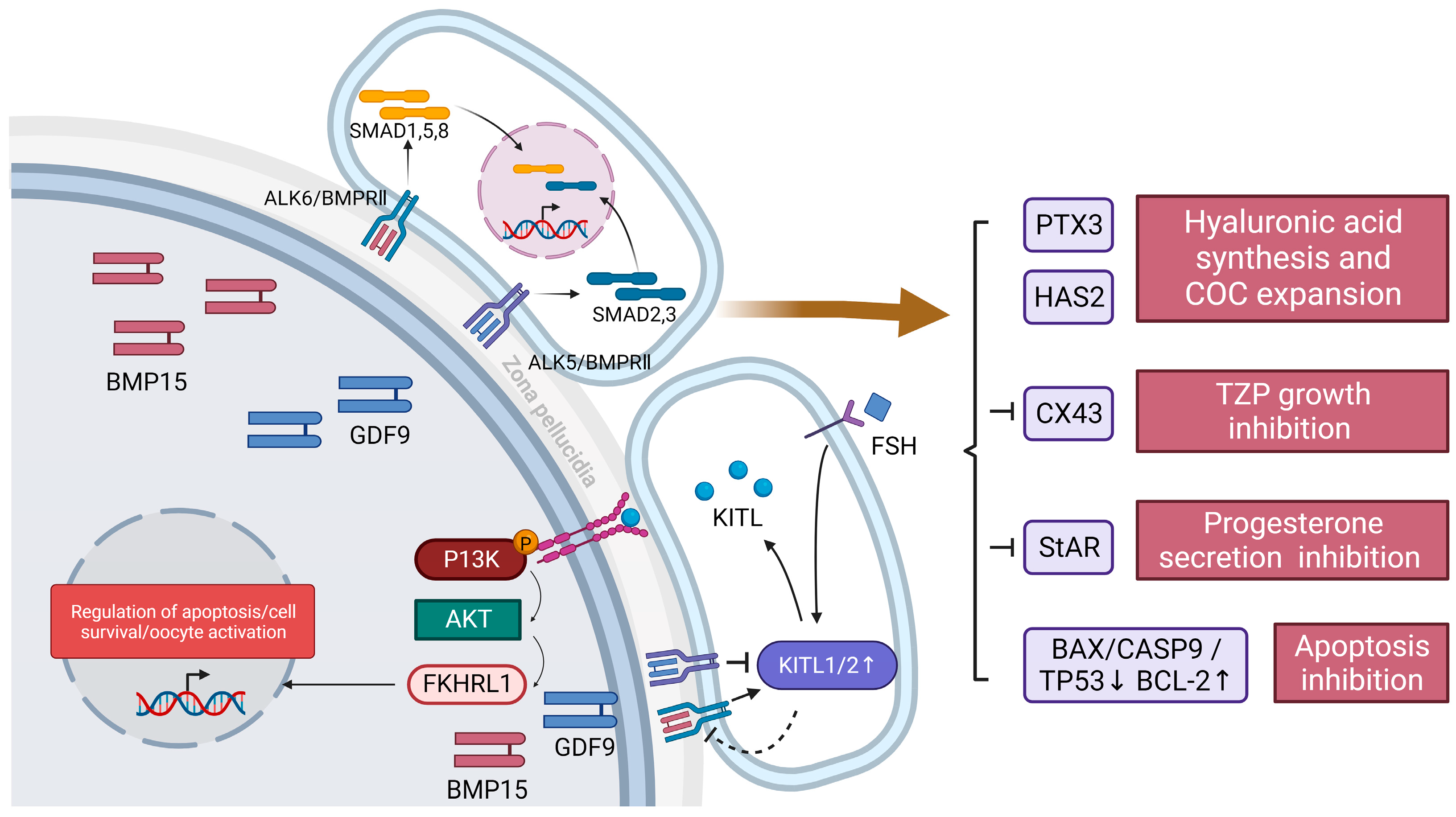
3.1. Paracrine Pathway
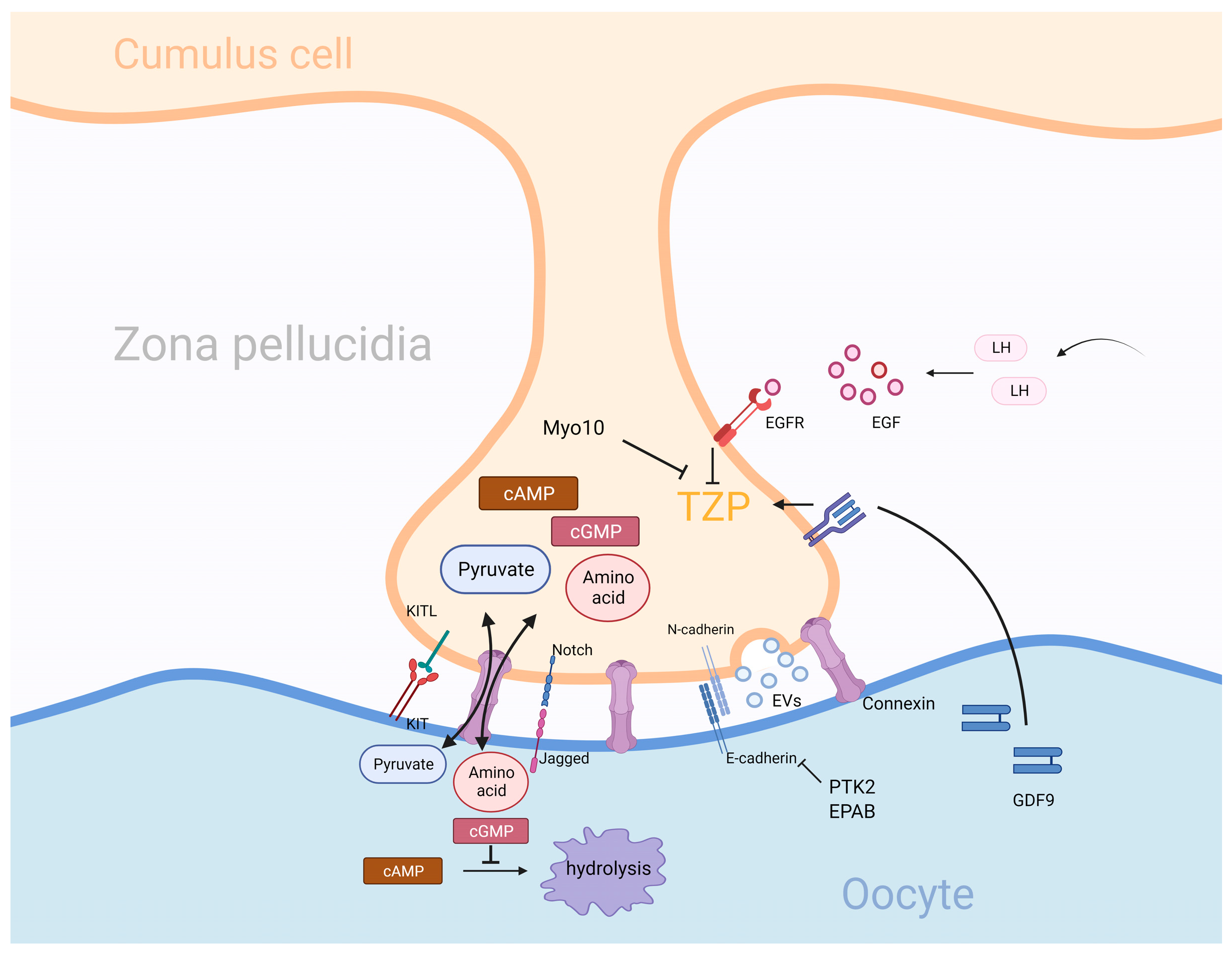
3.2. Gap Junction
3.3. Adhesion Connection and Extracellular Vesicles (EVs)
4. Glucose and Lipid Metabolism in Granulosa Cells and Oocytes in Healthy Women
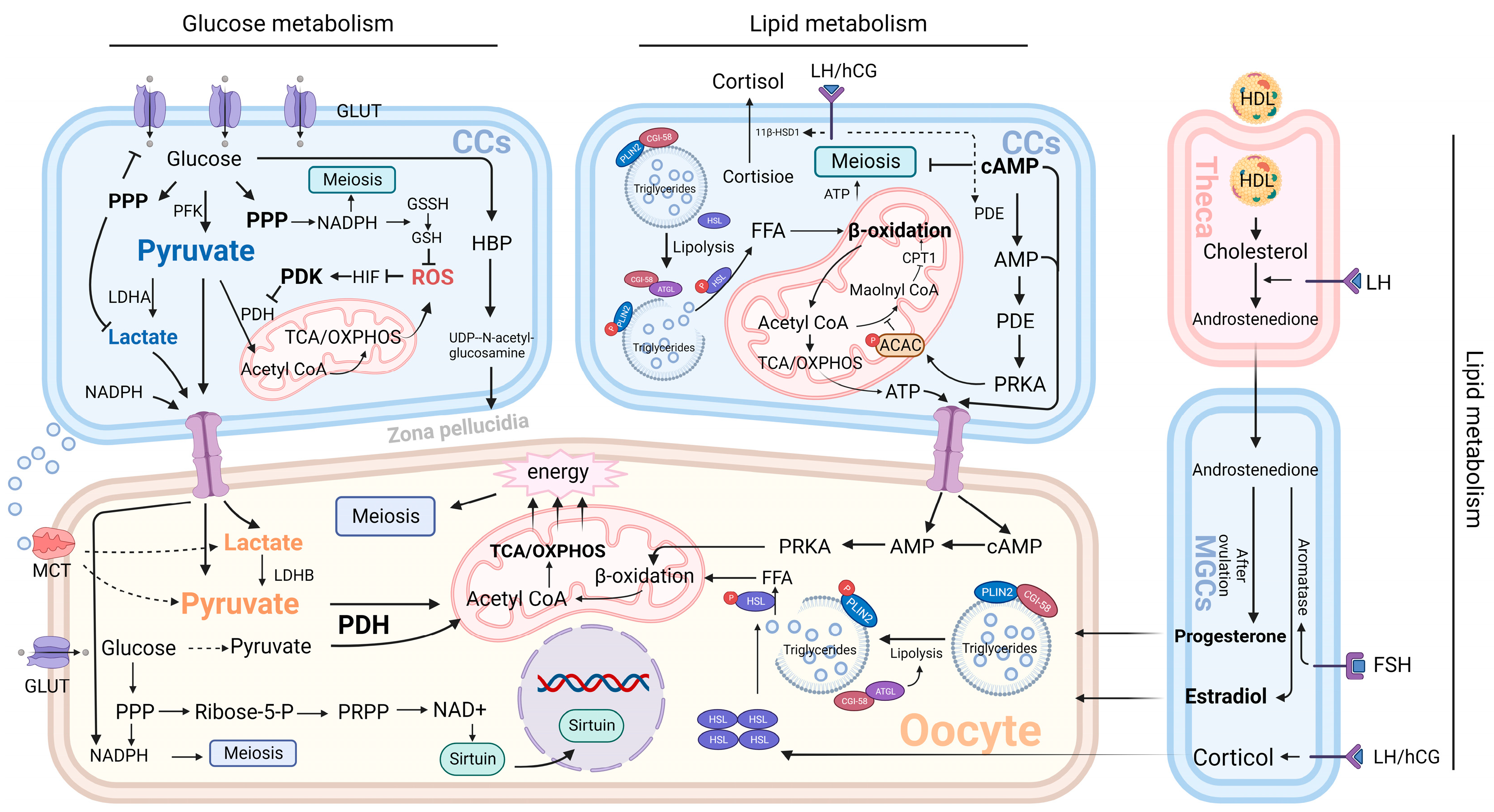
4.1. Characteristics of Energy Metabolism in Granulosa Cells and Oocytes
4.2. Effect of Granulosa Cell Glucose Metabolism on Oocytes
4.2.1. Glycolysis
4.2.2. Pentose Phosphate Pathway
4.2.3. Hexosamine Biosynthetic Pathway
4.3. Effect of Granulosa Cell Lipid Metabolism on Oocytes
4.3.1. Fatty Acid Metabolism
4.3.2. Steroid Metabolism
5. Relationship between Abnormal Glucose and Lipid Metabolism of Granulosa Cells and PCOS
5.1. Overview of PCOS
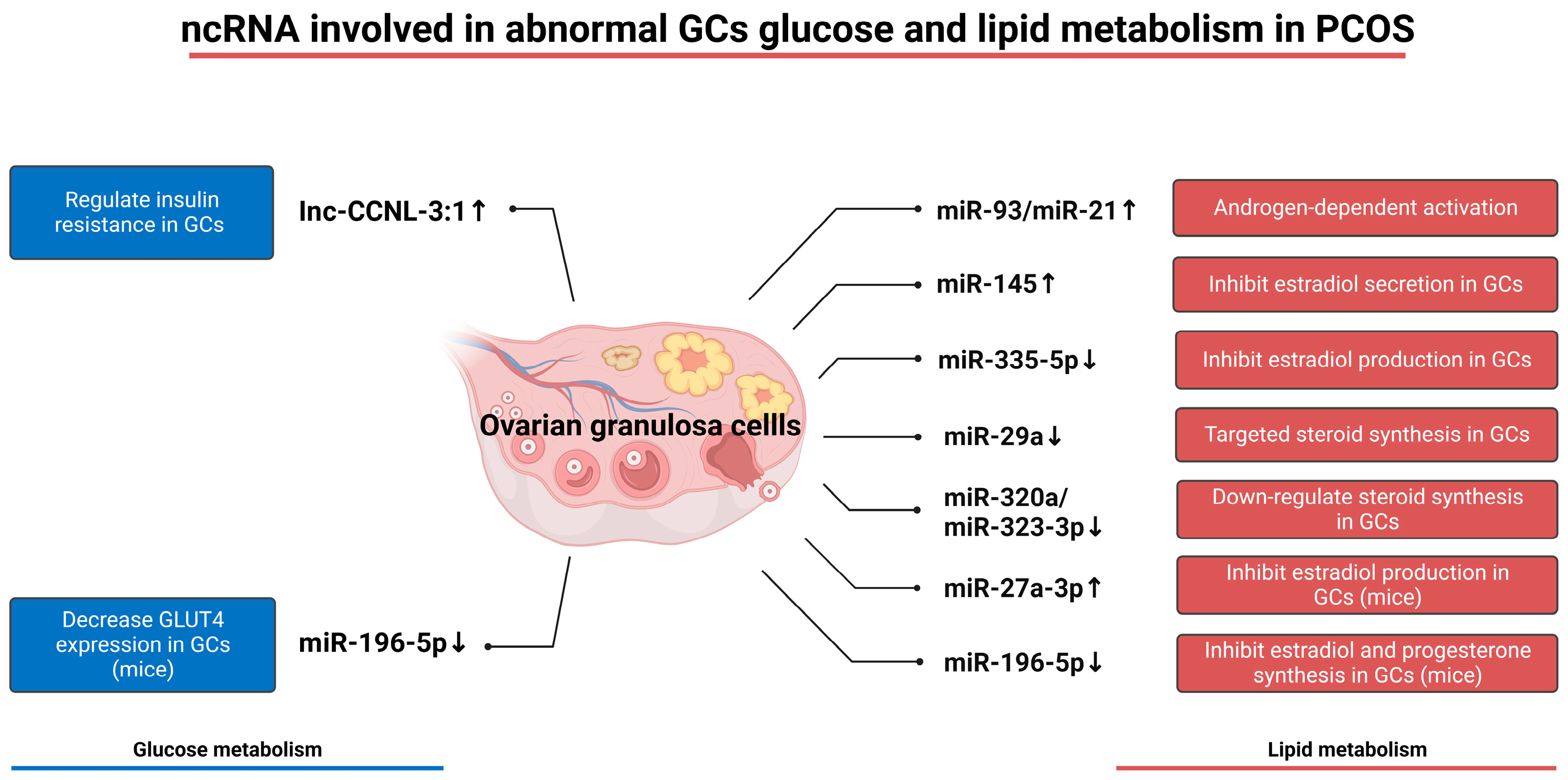
5.2. Abnormal Glucose Metabolism of Granulosa Cells in PCOS
5.3. Abnormal Lipid Metabolism of Granulosa Cells in PCOS
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ndefo, U.A.; Eaton, A.; Green, M.R. Polycystic ovary syndrome: A review of treatment options with a focus on pharmacological approaches. Pharm. Ther. 2013, 38, 336–355. [Google Scholar]
- Gougeon, A. Regulation of ovarian follicular development in primates: Facts and hypotheses. Endocr. Rev. 1996, 17, 121–155. [Google Scholar] [CrossRef] [PubMed]
- Uyar, A.; Torrealday, S.; Seli, E. Cumulus and granulosa cell markers of oocyte and embryo quality. Fertil. Steril. 2013, 99, 979–997. [Google Scholar] [CrossRef] [PubMed]
- Diaz, F.J.; O’Brien, M.J.; Wigglesworth, K.; Eppig, J.J. The preantral granulosa cell to cumulus cell transition in the mouse ovary: Development of competence to undergo expansion. Dev. Biol. 2006, 299, 91–104. [Google Scholar] [CrossRef]
- Da Silva-Buttkus, P.; Jayasooriya, G.S.; Mora, J.M.; Mobberley, M.; Ryder, T.A.; Baithun, M.; Stark, J.; Franks, S.; Hardy, K. Effect of cell shape and packing density on granulosa cell proliferation and formation of multiple layers during early follicle development in the ovary. J. Cell Sci. 2008, 121, 3890–3900. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.S.; Sabouni, R.; Cayton Vaught, K.C.; Owen, C.M.; Albertini, D.F.; Segars, J.H. Biomechanics and mechanical signaling in the ovary: A systematic review. J. Assist. Reprod. Genet. 2018, 35, 1135–1148. [Google Scholar] [CrossRef]
- Diaz, F.J.; Wigglesworth, K.; Eppig, J.J. Oocytes are required for the preantral granulosa cell to cumulus cell transition in mice. Dev. Biol. 2007, 305, 300–311. [Google Scholar] [CrossRef]
- Madgwick, S.; Jones, K.T. How eggs arrest at metaphase II: MPF stabilisation plus APC/C inhibition equals Cytostatic Factor. Cell Div. 2007, 2, 4. [Google Scholar] [CrossRef]
- Salustri, A.; Garlanda, C.; Hirsch, E.; De Acetis, M.; Maccagno, A.; Bottazzi, B.; Doni, A.; Bastone, A.; Mantovani, G.; Beck Peccoz, P.; et al. PTX3 plays a key role in the organization of the cumulus oophorus extracellular matrix and in in vivo fertilization. Development 2004, 131, 1577–1586. [Google Scholar] [CrossRef]
- Salustri, A.; Yanagishita, M.; Underhill, C.B.; Laurent, T.C.; Hascall, V.C. Localization and synthesis of hyaluronic acid in the cumulus cells and mural granulosa cells of the preovulatory follicle. Dev. Biol. 1992, 151, 541–551. [Google Scholar] [CrossRef]
- Ikawa, M.; Inoue, N.; Benham, A.M.; Okabe, M. Fertilization: A sperm’s journey to and interaction with the oocyte. J. Clin. Investig. 2010, 120, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Niswender, G.D.; Juengel, J.L.; Silva, P.J.; Rollyson, M.K.; McIntush, E.W. Mechanisms controlling the function and life span of the corpus luteum. Physiol. Rev. 2000, 80, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Erickson, G.F.; Hofeditz, C.; Unger, M.; Allen, W.R.; Dulbecco, R. A monoclonal antibody to a mammary cell line recognizes two distinct subtypes of ovarian granulosa cells. Endocrinology 1985, 117, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Havelock, J.C.; Rainey, W.E.; Carr, B.R. Ovarian granulosa cell lines. Mol. Cell. Endocrinol. 2004, 228, 67–78. [Google Scholar] [CrossRef]
- Sugiura, K.; Pendola, F.L.; Eppig, J.J. Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells: Energy metabolism. Dev. Biol. 2005, 279, 20–30. [Google Scholar] [CrossRef]
- Thomas, R.E.; Armstrong, D.T.; Gilchrist, R.B. Bovine cumulus cell-oocyte gap junctional communication during in vitro maturation in response to manipulation of cell-specific cyclic adenosine 3′,5′-monophosophate levels. Biol. Reprod. 2004, 70, 548–556. [Google Scholar] [CrossRef]
- Hillier, S.G.; Tetsuka, M. Role of androgens in follicle maturation and atresia. Baillieres Clin. Obstet. Gynaecol. 1997, 11, 249–260. [Google Scholar] [CrossRef]
- Plancha, C.E.; Sanfins, A.; Rodrigues, P.; Albertini, D. Cell polarity during folliculogenesis and oogenesis. Reprod. Biomed. Online 2005, 10, 478–484. [Google Scholar] [CrossRef]
- Saeed-Zidane, M.; Linden, L.; Salilew-Wondim, D.; Held, E.; Neuhoff, C.; Tholen, E.; Hoelker, M.; Schellander, K.; Tesfaye, D. Cellular and exosome mediated molecular defense mechanism in bovine granulosa cells exposed to oxidative stress. PLoS ONE 2017, 12, e0187569. [Google Scholar] [CrossRef]
- Mora, J.M.; Fenwick, M.A.; Castle, L.; Baithun, M.; Ryder, T.A.; Mobberley, M.; Carzaniga, R.; Franks, S.; Hardy, K. Characterization and significance of adhesion and junction-related proteins in mouse ovarian follicles. Biol. Reprod. 2012, 86, 153. [Google Scholar] [CrossRef]
- Picton, H.; Briggs, D.; Gosden, R. The molecular basis of oocyte growth and development. Mol. Cell. Endocrinol. 1998, 145, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; O’Brien, M.; Wigglesworth, K. Mammalian oocyte growth and development in vitro. Mol. Reprod. Dev. 1996, 44, 260–273. [Google Scholar] [CrossRef]
- Kolesarova, A.; Sirotkin, A.V.; Mellen, M.; Roychoudhury, S. Possible intracellular regulators of female sexual maturation. Physiol. Res. 2015, 64, 379–386. [Google Scholar] [CrossRef]
- Xie, H.L.; Wang, Y.B.; Jiao, G.Z.; Kong, D.L.; Li, Q.; Li, H.; Zheng, L.L.; Tan, J.H. Effects of glucose metabolism during in vitro maturation on cytoplasmic maturation of mouse oocytes. Sci. Rep. 2016, 6, 20764. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, F.; Smitz, J. Molecular control of oogenesis. Biochim. Biophys. Acta 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed]
- Juengel, J.L.; McNatty, K.P. The role of proteins of the transforming growth factor-beta superfamily in the intraovarian regulation of follicular development. Hum. Reprod. Update 2005, 11, 143–160. [Google Scholar] [CrossRef]
- de Castro, F.C.; Cruz, M.H.; Leal, C.L. Role of Growth Differentiation Factor 9 and Bone Morphogenetic Protein 15 in Ovarian Function and Their Importance in Mammalian Female Fertility—A Review. Asian Australas. J. Anim. Sci. 2016, 29, 1065–1074. [Google Scholar] [CrossRef]
- Persani, L.; Rossetti, R.; Di Pasquale, E.; Cacciatore, C.; Fabre, S. The fundamental role of bone morphogenetic protein 15 in ovarian function and its involvement in female fertility disorders. Hum. Reprod. Update 2014, 20, 869–883. [Google Scholar] [CrossRef]
- Dube, J.L.; Wang, P.; Elvin, J.; Lyons, K.M.; Celeste, A.J.; Matzuk, M.M. The bone morphogenetic protein 15 gene is X-linked and expressed in oocytes. Mol. Endocrinol. 1998, 12, 1809–1817. [Google Scholar] [CrossRef]
- Peng, J.; Li, Q.; Wigglesworth, K.; Rangarajan, A.; Kattamuri, C.; Peterson, R.T.; Eppig, J.J.; Thompson, T.B.; Matzuk, M.M. Growth differentiation factor 9:bone morphogenetic protein 15 heterodimers are potent regulators of ovarian functions. Proc. Natl. Acad. Sci. USA 2013, 110, E776–E785. [Google Scholar] [CrossRef]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef]
- Mazerbourg, S.; Klein, C.; Roh, J.; Kaivo-Oja, N.; Mottershead, D.G.; Korchynskyi, O.; Ritvos, O.; Hsueh, A.J. Growth differentiation factor-9 signaling is mediated by the type I receptor, activin receptor-like kinase 5. Mol. Endocrinol. 2004, 18, 653–665. [Google Scholar] [CrossRef]
- Mazerbourg, S.; Hsueh, A.J. Genomic analyses facilitate identification of receptors and signalling pathways for growth differentiation factor 9 and related orphan bone morphogenetic protein/growth differentiation factor ligands. Hum. Reprod. Update 2006, 12, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Mottershead, D.G.; Ritter, L.J.; Gilchrist, R.B. Signalling pathways mediating specific synergistic interactions between GDF9 and BMP15. Mol. Hum. Reprod. 2012, 18, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, C.; Jiang, H.; Gao, Y.; Xu, M.; Wang, J.; Liu, S.; Fu, Y.; Sun, X.; Xu, J.; et al. Regulatory Role of miRNA-375 in Expression of BMP15/GDF9 Receptors and its Effect on Proliferation and Apoptosis of Bovine Cumulus Cells. Cell Physiol. Biochem. 2017, 41, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Qiao, J.; Leung, P.C. Oocyte-somatic cell interactions in the human ovary-novel role of bone morphogenetic proteins and growth differentiation factors. Hum. Reprod. Update 2016, 23, 1–18. [Google Scholar] [CrossRef]
- Chang, H.M.; Cheng, J.C.; Leung, P.C. Theca-derived BMP4 and BMP7 down-regulate connexin43 expression and decrease gap junction intercellular communication activity in immortalized human granulosa cells. J. Clin. Endocrinol. Metab. 2013, 98, E437–E445. [Google Scholar] [CrossRef]
- Chang, H.M.; Cheng, J.C.; Taylor, E.; Leung, P.C. Oocyte-derived BMP15 but not GDF9 down-regulates connexin43 expression and decreases gap junction intercellular communication activity in immortalized human granulosa cells. Mol. Hum. Reprod. 2014, 20, 373–383. [Google Scholar] [CrossRef]
- El-Hayek, S.; Yang, Q.; Abbassi, L.; FitzHarris, G.; Clarke, H.J. Mammalian Oocytes Locally Remodel Follicular Architecture to Provide the Foundation for Germline-Soma Communication. Curr. Biol. 2018, 28, 1124–1131.e1123. [Google Scholar] [CrossRef]
- Orisaka, M.; Jiang, J.Y.; Orisaka, S.; Kotsuji, F.; Tsang, B.K. Growth differentiation factor 9 promotes rat preantral follicle growth by up-regulating follicular androgen biosynthesis. Endocrinology 2009, 150, 2740–2748. [Google Scholar] [CrossRef] [PubMed]
- Hussein, T.S.; Froiland, D.A.; Amato, F.; Thompson, J.G.; Gilchrist, R.B. Oocytes prevent cumulus cell apoptosis by maintaining a morphogenic paracrine gradient of bone morphogenetic proteins. J. Cell Sci. 2005, 118, 5257–5268. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Hirose, M.; Inoue, K.; Hiura, H.; Miki, H.; Ogonuki, N.; Sugimoto, M.; Abe, K.; Kanatsu-Shinohara, M.; Kono, T.; et al. Large-scale production of growing oocytes in vitro from neonatal mouse ovaries. Int. J. Dev. Biol. 2009, 53, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Han, C.S.; Zhang, X.S.; Yuan, J.X.; Hu, Z.Y.; Liu, Y.X. Signal transduction of stem cell factor in promoting early follicle development. Mol. Cell. Endocrinol. 2005, 229, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.H.; Ethier, J.F.; Shimasaki, S.; Vanderhyden, B.C. Follicle-stimulating hormone regulates oocyte growth by modulation of expression of oocyte and granulosa cell factors. Endocrinology 2005, 146, 941–949. [Google Scholar] [CrossRef]
- Joyce, I.M.; Clark, A.T.; Pendola, F.L.; Eppig, J.J. Comparison of recombinant growth differentiation factor-9 and oocyte regulation of KIT ligand messenger ribonucleic acid expression in mouse ovarian follicles. Biol. Reprod. 2000, 63, 1669–1675. [Google Scholar] [CrossRef]
- Anderson, E.; Wilkinson, R.F.; Lee, G.; Meller, S. A correlative microscopical analysis of differentiating ovarian follicles of mammals. J. Morphol. 1978, 156, 339–366. [Google Scholar] [CrossRef]
- Baena, V.; Terasaki, M. Three-dimensional organization of transzonal projections and other cytoplasmic extensions in the mouse ovarian follicle. Sci. Rep. 2019, 9, 1262. [Google Scholar] [CrossRef]
- Macaulay, A.D.; Gilbert, I.; Caballero, J.; Barreto, R.; Fournier, E.; Tossou, P.; Sirard, M.A.; Clarke, H.J.; Khandjian, E.W.; Richard, F.J.; et al. The gametic synapse: RNA transfer to the bovine oocyte. Biol. Reprod. 2014, 91, 90. [Google Scholar] [CrossRef]
- Willecke, K.; Eiberger, J.; Degen, J.; Eckardt, D.; Romualdi, A.; Guldenagel, M.; Deutsch, U.; Sohl, G. Structural and functional diversity of connexin genes in the mouse and human genome. Biol. Chem. 2002, 383, 725–737. [Google Scholar] [CrossRef]
- Kidder, G.M.; Vanderhyden, B.C. Bidirectional communication between oocytes and follicle cells: Ensuring oocyte developmental competence. Can. J. Physiol. Pharmacol. 2010, 88, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Chiquoine, A.D. The development of the zona pellucida of the mammalian ovum. Am. J. Anat. 1960, 106, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Fushii, M.; Yamada, R.; Lee, J.; Miyano, T. Reestablishment of transzonal projections and growth of bovine oocytes in vitro. J. Reprod. Dev. 2021, 67, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Granados-Aparici, S.; Volodarsky-Perel, A.; Yang, Q.; Anam, S.; Tulandi, T.; Buckett, W.; Son, W.Y.; Younes, G.; Chung, J.T.; Jin, S.; et al. MYO10 promotes transzonal projection-dependent germ line-somatic contact during mammalian folliculogenesisdagger. Biol. Reprod. 2022, 107, 474–487. [Google Scholar] [CrossRef]
- Clarke, H.J. Transzonal projections: Essential structures mediating intercellular communication in the mammalian ovarian follicle. Mol. Reprod. Dev. 2022, 89, 509–525. [Google Scholar] [CrossRef]
- Conti, M.; Hsieh, M.; Park, J.Y.; Su, Y.Q. Role of the epidermal growth factor network in ovarian follicles. Mol. Endocrinol. 2006, 20, 715–723. [Google Scholar] [CrossRef]
- Jaffe, L.A.; Egbert, J.R. Regulation of Mammalian Oocyte Meiosis by Intercellular Communication Within the Ovarian Follicle. Annu. Rev. Physiol. 2017, 79, 237–260. [Google Scholar] [CrossRef]
- Corbett, H.E.; Dube, C.D.; Slow, S.; Lever, M.; Trasler, J.M.; Baltz, J.M. Uptake of betaine into mouse cumulus–oocyte complexes via the SLC7A6 isoform of y + L transporter. Biol. Reprod. 2014, 90, 81. [Google Scholar] [CrossRef]
- Wigglesworth, K.; Lee, K.B.; O’Brien, M.J.; Peng, J.; Matzuk, M.M.; Eppig, J.J. Bidirectional communication between oocytes and ovarian follicular somatic cells is required for meiotic arrest of mammalian oocytes. Proc. Natl. Acad. Sci. USA 2013, 110, E3723–E3729. [Google Scholar] [CrossRef]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell 2018, 72, 1021–1034.e1024. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Risal, S.; Gorre, N.; Busayavalasa, K.; Li, X.; Shen, Y.; Bosbach, B.; Brannstrom, M.; Liu, K. Somatic cells initiate primordial follicle activation and govern the development of dormant oocytes in mice. Curr. Biol. 2014, 24, 2501–2508. [Google Scholar] [CrossRef] [PubMed]
- Vanorny, D.A.; Prasasya, R.D.; Chalpe, A.J.; Kilen, S.M.; Mayo, K.E. Notch signaling regulates ovarian follicle formation and coordinates follicular growth. Mol. Endocrinol. 2014, 28, 499–511. [Google Scholar] [CrossRef]
- Masoudi, M.; Yamini, N.; Salehi, F.; Aflatoonian, R.; Kutenaee, M.A.; Esfandiyari, S.; Hosseini, E.; Asadi, E.; Najarkolaei, A.H.; Mortezaee, K.; et al. Notch signaling pathway in cumulus cells reflecting zygote and embryo quality in polycystic ovary syndrome. Arch. Gynecol. Obstet. 2021, 304, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Machtinger, R.; Laurent, L.C.; Baccarelli, A.A. Extracellular vesicles: Roles in gamete maturation, fertilization and embryo implantation. Hum. Reprod. Update 2016, 22, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, C. Exosome-mediated communication in the ovarian follicle. J. Assist. Reprod. Genet. 2016, 33, 303–311. [Google Scholar] [CrossRef]
- Macaulay, A.D.; Gilbert, I.; Scantland, S.; Fournier, E.; Ashkar, F.; Bastien, A.; Saadi, H.A.; Gagne, D.; Sirard, M.A.; Khandjian, E.W.; et al. Cumulus Cell Transcripts Transit to the Bovine Oocyte in Preparation for Maturation. Biol. Reprod. 2016, 94, 16. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Veeramachaneni, D.N.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-secreted vesicles in equine ovarian follicular fluid contain miRNAs and proteins: A possible new form of cell communication within the ovarian follicle. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef]
- Hung, W.T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Carnevale, E.M.; Winger, Q.A.; Bouma, G.J. Regulation of ACVR1 and ID2 by cell-secreted exosomes during follicle maturation in the mare. Reprod. Biol. Endocrinol. 2014, 12, 44. [Google Scholar] [CrossRef]
- Sohel, M.M.; Hoelker, M.; Noferesti, S.S.; Salilew-Wondim, D.; Tholen, E.; Looft, C.; Rings, F.; Uddin, M.J.; Spencer, T.E.; Schellander, K.; et al. Exosomal and Non-Exosomal Transport of Extra-Cellular microRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLoS ONE 2013, 8, e78505. [Google Scholar] [CrossRef] [PubMed]
- Kansaku, K.; Itami, N.; Kawahara-Miki, R.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Differential effects of mitochondrial inhibitors on porcine granulosa cells and oocytes. Theriogenology 2017, 103, 98–103. [Google Scholar] [CrossRef]
- Yu, Y.; Dumollard, R.; Rossbach, A.; Lai, F.A.; Swann, K. Redistribution of mitochondria leads to bursts of ATP production during spontaneous mouse oocyte maturation. J. Cell. Physiol. 2010, 224, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Iwata, H.; Goto, H.; Shiratuki, S.; Tanaka, H.; Monji, Y.; Kuwayama, T. Effect of maternal age on the developmental competence and progression of nuclear maturation in bovine oocytes. Mol. Reprod. Dev. 2010, 77, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Davy, P.M.; Gardner, L.H.; Mathews, J.; Yamazaki, Y.; Allsopp, R.C. Hypoxia Inducible Factor 1 Alpha Is Expressed in Germ Cells throughout the Murine Life Cycle. PLoS ONE 2016, 11, e0154309. [Google Scholar] [CrossRef] [PubMed]
- Warzych, E.; Lipinska, P. Energy metabolism of follicular environment during oocyte growth and maturation. J. Reprod. Dev. 2020, 66, 1–7. [Google Scholar] [CrossRef]
- Cetica, P.; Pintos, L.; Dalvit, G.; Beconi, M. Activity of key enzymes involved in glucose and triglyceride catabolism during bovine oocyte maturation in vitro. Reproduction 2002, 124, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chi, M.M.; Schedl, T.; Moley, K.H. An intercellular pathway for glucose transport into mouse oocytes. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1511–E1518. [Google Scholar] [CrossRef]
- Kuchiiwa, T.; Nio-Kobayashi, J.; Takahashi-Iwanaga, H.; Yajima, T.; Iwanaga, T. Cellular expression of monocarboxylate transporters in the female reproductive organ of mice: Implications for the genital lactate shuttle. Histochem. Cell Biol. 2011, 135, 351–360. [Google Scholar] [CrossRef]
- Xu, W.; Li, L.; Sun, J.; Zhu, S.; Yan, Z.; Gao, L.; Gao, C.; Cui, Y.; Mao, C. Putrescine delays postovulatory aging of mouse oocytes by upregulating PDK4 expression and improving mitochondrial activity. Aging 2018, 10, 4093–4106. [Google Scholar] [CrossRef]
- Bermejo-Alvarez, P.; Lonergan, P.; Rizos, D.; Gutierrez-Adan, A. Low oxygen tension during IVM improves bovine oocyte competence and enhances anaerobic glycolysis. Reprod. Biomed. Online 2010, 20, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Uhde, K.; van Tol, H.T.A.; Stout, T.A.E.; Roelen, B.A.J. Metabolomic profiles of bovine cumulus cells and cumulus–oocyte-complex-conditioned medium during maturation in vitro. Sci. Rep. 2018, 8, 9477. [Google Scholar] [CrossRef] [PubMed]
- Herubel, F.; El Mouatassim, S.; Guerin, P.; Frydman, R.; Menezo, Y. Genetic expression of monocarboxylate transporters during human and murine oocyte maturation and early embryonic development. Zygote 2002, 10, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Turhan, A.; Pereira, M.T.; Schuler, G.; Bleul, U.; Kowalewski, M.P. Hypoxia-inducible factor (HIF1alpha) inhibition modulates cumulus cell function and affects bovine oocyte maturation in vitrodagger. Biol. Reprod. 2021, 104, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Richani, D.; Dunning, K.R.; Thompson, J.G.; Gilchrist, R.B. Metabolic co-dependence of the oocyte and cumulus cells: Essential role in determining oocyte developmental competence. Hum. Reprod. Update 2021, 27, 27–47. [Google Scholar] [CrossRef]
- Purcell, S.H.; Chi, M.M.; Lanzendorf, S.; Moley, K.H. Insulin-stimulated glucose uptake occurs in specialized cells within the cumulus oocyte complex. Endocrinology 2012, 153, 2444–2454. [Google Scholar] [CrossRef]
- Su, Y.Q.; Sugiura, K.; Eppig, J.J. Mouse oocyte control of granulosa cell development and function: Paracrine regulation of cumulus cell metabolism. Semin. Reprod. Med. 2009, 27, 32–42. [Google Scholar] [CrossRef]
- Kumar, P.; Verma, A.; Kumar, M.; De, S.; Kumar, R.; Datta, T.K. Expression pattern of glucose metabolism genes correlate with development rate of buffalo oocytes and embryos in vitro under low oxygen condition. J. Assist. Reprod. Genet. 2015, 32, 471–478. [Google Scholar] [CrossRef]
- Molinari, E.; Bar, H.; Pyle, A.M.; Patrizio, P. Transcriptome analysis of human cumulus cells reveals hypoxia as the main determinant of follicular senescence. Mol. Hum. Reprod. 2016, 22, 866–876. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef] [PubMed]
- Shaeib, F.; Khan, S.N.; Ali, I.; Thakur, M.; Saed, M.G.; Dai, J.; Awonuga, A.O.; Banerjee, J.; Abu-Soud, H.M. The Defensive Role of Cumulus Cells Against Reactive Oxygen Species Insult in Metaphase II Mouse Oocytes. Reprod. Sci. 2016, 23, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Nikiforov, A.; Kulikova, V.; Ziegler, M. The human NAD metabolome: Functions, metabolism and compartmentalization. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Herrick, J.R.; Brad, A.M.; Krisher, R.L. Chemical manipulation of glucose metabolism in porcine oocytes: Effects on nuclear and cytoplasmic maturation in vitro. Reproduction 2006, 131, 289–298. [Google Scholar] [CrossRef]
- Gutnisky, C.; Dalvit, G.C.; Thompson, J.G.; Cetica, P.D. Pentose phosphate pathway activity: Effect on in vitro maturation and oxidative status of bovine oocytes. Reprod. Fertil. Dev. 2014, 26, 931–942. [Google Scholar] [CrossRef]
- Krisher, R.L.; Brad, A.M.; Herrick, J.R.; Sparman, M.L.; Swain, J.E. A comparative analysis of metabolism and viability in porcine oocytes during in vitro maturation. Anim. Reprod. Sci. 2007, 98, 72–96. [Google Scholar] [CrossRef]
- Steeves, T.E.; Gardner, D.K. Temporal and differential effects of amino acids on bovine embryo development in culture. Biol. Reprod. 1999, 61, 731–740. [Google Scholar] [CrossRef]
- Imai, S.I.; Guarente, L. It takes two to tango: NAD(+) and sirtuins in aging/longevity control. NPJ Aging Mech. Dis. 2016, 2, 16017. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Listijono, D.R.; Ho, W.J.; Riepsamen, A.H.; Goss, D.M.; Richani, D.; Jin, X.L.; Mahbub, S.; Campbell, J.M.; Habibalahi, A.; et al. NAD(+) Repletion Rescues Female Fertility during Reproductive Aging. Cell Rep. 2020, 30, 1670–1681.e1677. [Google Scholar] [CrossRef]
- Wells, L.; Whelan, S.A.; Hart, G.W. O-GlcNAc: A regulatory post-translational modification. Biochem. Biophys. Res. Commun. 2003, 302, 435–441. [Google Scholar] [CrossRef]
- Zachara, N.E.; Hart, G.W. O-GlcNAc a sensor of cellular state: The role of nucleocytoplasmic glycosylation in modulating cellular function in response to nutrition and stress. Biochim. Biophys. Acta 2004, 1673, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.; Bacote, V.; Traxinger, R.R. Discovery of a metabolic pathway mediating glucose-induced desensitization of the glucose transport system. Role of hexosamine biosynthesis in the induction of insulin resistance. J. Biol. Chem. 1991, 266, 4706–4712. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.L.; Cetica, P.D.; Beconi, M.T.; Kind, K.L.; Gilchrist, R.B.; Thompson, J.G. Influence of oocyte-secreted factors and culture duration on the metabolic activity of bovine cumulus cell complexes. Reproduction 2003, 126, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Richani, D.; Gilchrist, R.B. The epidermal growth factor network: Role in oocyte growth, maturation and developmental competence. Hum. Reprod. Update 2018, 24, 1–14. [Google Scholar] [CrossRef]
- Bertolin, K.; Meinsohn, M.C.; Suzuki, J.; Gossen, J.; Schoonjans, K.; Duggavathi, R.; Murphy, B.D. Ovary-specific depletion of the nuclear receptor Nr5a2 compromises expansion of the cumulus oophorus but not fertilization by intracytoplasmic sperm injection. Biol. Reprod. 2017, 96, 1231–1243. [Google Scholar] [CrossRef]
- Silva, R.C.; Bao, S.N.; Jivago, J.L.; Lucci, C.M. Ultrastructural characterization of porcine oocytes and adjacent follicular cells during follicle development: Lipid component evolution. Theriogenology 2011, 76, 1647–1657. [Google Scholar] [CrossRef]
- Sturmey, R.G.; O’Toole, P.J.; Leese, H.J. Fluorescence resonance energy transfer analysis of mitochondrial:lipid association in the porcine oocyte. Reproduction 2006, 132, 829–837. [Google Scholar] [CrossRef]
- MacPherson, R.E.; Peters, S.J. Piecing together the puzzle of perilipin proteins and skeletal muscle lipolysis. Appl. Physiol. Nutr. Metab. 2015, 40, 641–651. [Google Scholar] [CrossRef]
- Liu, T.; Qu, J.; Tian, M.; Yang, R.; Song, X.; Li, R.; Yan, J.; Qiao, J. Lipid Metabolic Process Involved in Oocyte Maturation During Folliculogenesis. Front. Cell Dev. Biol. 2022, 10, 806890. [Google Scholar] [CrossRef]
- Simerman, A.A.; Hill, D.L.; Grogan, T.R.; Elashoff, D.; Clarke, N.J.; Goldstein, E.H.; Manrriquez, A.N.; Chazenbalk, G.D.; Dumesic, D.A. Intrafollicular cortisol levels inversely correlate with cumulus cell lipid content as a possible energy source during oocyte meiotic resumption in women undergoing ovarian stimulation for in vitro fertilization. Fertil. Steril. 2015, 103, 249–257. [Google Scholar] [CrossRef]
- Hardie, D.G.; Pan, D.A. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase. Biochem. Soc. Trans. 2002, 30, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Luong, X.G.; Daldello, E.M.; Rajkovic, G.; Yang, C.R.; Conti, M. Genome-wide analysis reveals a switch in the translational program upon oocyte meiotic resumption. Nucleic Acids Res. 2020, 48, 3257–3276. [Google Scholar] [CrossRef] [PubMed]
- Aardema, H.; van Tol, H.T.A.; Wubbolts, R.W.; Brouwers, J.; Gadella, B.M.; Roelen, B.A.J. Stearoyl-CoA desaturase activity in bovine cumulus cells protects the oocyte against saturated fatty acid stress. Biol. Reprod. 2017, 96, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Auclair, S.; Uzbekov, R.; Elis, S.; Sanchez, L.; Kireev, I.; Lardic, L.; Dalbies-Tran, R.; Uzbekova, S. Absence of cumulus cells during in vitro maturation affects lipid metabolism in bovine oocytes. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E599–E613. [Google Scholar] [CrossRef]
- Yong, P.Y.; Thong, K.J.; Andrew, R.; Walker, B.R.; Hillier, S.G. Development-related increase in cortisol biosynthesis by human granulosa cells. J. Clin. Endocrinol. Metab. 2000, 85, 4728–4733. [Google Scholar] [CrossRef] [PubMed]
- Stimson, R.H.; Anderson, A.J.; Ramage, L.E.; Macfarlane, D.P.; de Beaux, A.C.; Mole, D.J.; Andrew, R.; Walker, B.R. Acute physiological effects of glucocorticoids on fuel metabolism in humans are permissive but not direct. Diabetes Obes. Metab. 2017, 19, 883–891. [Google Scholar] [CrossRef]
- Annema, W.; Tietge, U.J. Regulation of reverse cholesterol transport—A comprehensive appraisal of available animal studies. Nutr. Metab. 2012, 9, 25. [Google Scholar] [CrossRef]
- Shelness, G.S.; Ledford, A.S. Evolution and mechanism of apolipoprotein B-containing lipoprotein assembly. Curr. Opin. Lipidol. 2005, 16, 325–332. [Google Scholar] [CrossRef]
- Dikkers, A.; Tietge, U.J. Biliary cholesterol secretion: More than a simple ABC. World J. Gastroenterol. 2010, 16, 5936–5945. [Google Scholar]
- Jaspard, B.; Fournier, N.; Vieitez, G.; Atger, V.; Barbaras, R.; Vieu, C.; Manent, J.; Chap, H.; Perret, B.; Collet, X. Structural and functional comparison of HDL from homologous human plasma and follicular fluid. A model for extravascular fluid. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1605–1613. [Google Scholar] [CrossRef]
- Martinez, L.O.; Jacquet, S.; Terce, F.; Collet, X.; Perret, B.; Barbaras, R. New insight on the molecular mechanisms of high-density lipoprotein cellular interactions. Cell Mol. Life Sci. 2004, 61, 2343–2360. [Google Scholar] [CrossRef]
- Hillier, S.G.; Whitelaw, P.F.; Smyth, C.D. Follicular oestrogen synthesis: The ‘two-cell, two-gonadotrophin’ model revisited. Mol. Cell. Endocrinol. 1994, 100, 51–54. [Google Scholar] [CrossRef]
- Miettinen, H.E.; Rayburn, H.; Krieger, M. Abnormal lipoprotein metabolism and reversible female infertility in HDL receptor (SR-BI)-deficient mice. J. Clin. Investig. 2001, 108, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Yesilaltay, A.; Dokshin, G.A.; Busso, D.; Wang, L.; Galiani, D.; Chavarria, T.; Vasile, E.; Quilaqueo, L.; Orellana, J.A.; Walzer, D.; et al. Excess cholesterol induces mouse egg activation and may cause female infertility. Proc. Natl. Acad. Sci. USA 2014, 111, E4972–E4980. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Armanini, D.; Boscaro, M.; Bordin, L.; Sabbadin, C. Controversies in the Pathogenesis, Diagnosis and Treatment of PCOS: Focus on Insulin Resistance, Inflammation, and Hyperandrogenism. Int. J. Mol. Sci. 2022, 23, 4110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, H.; Cui, Y.; Wang, W.; Qin, Y.; You, L.; Chan, W.Y.; Sun, Y.; Chen, Z.J. Metabolic actions of insulin in ovarian granulosa cells were unaffected by hyperandrogenism. Endocrine 2016, 53, 823–830. [Google Scholar] [CrossRef]
- Kim, E.; Seok, H.H.; Lee, S.Y.; Lee, D.R.; Moon, J.; Yoon, T.K.; Lee, W.S.; Lee, K.A. Correlation between Expression of Glucose Transporters in Granulosa Cells and Oocyte Quality in Women with Polycystic Ovary Syndrome. Endocrinol. Metab. 2014, 29, 40–47. [Google Scholar] [CrossRef]
- Liu, X.; Sun, C.; Zou, K.; Li, C.; Chen, X.; Gu, H.; Zhou, Z.; Yang, Z.; Tu, Y.; Qin, N.; et al. Novel PGK1 determines SKP2-dependent AR stability and reprograms granular cell glucose metabolism facilitating ovulation dysfunction. EBioMedicine 2020, 61, 103058. [Google Scholar] [CrossRef]
- Wang, J.; Wu, X. The effects of mitochondrial dysfunction on energy metabolism switch by HIF-1alpha signalling in granulosa cells of polycystic ovary syndrome. Endokrynol. Pol. 2020, 71, 134–145. [Google Scholar] [CrossRef]
- Min, Z.; Gao, Q.; Zhen, X.; Fan, Y.; Tan, T.; Li, R.; Zhao, Y.; Yu, Y. New insights into the genic and metabolic characteristics of induced pluripotent stem cells from polycystic ovary syndrome women. Stem Cell Res. Ther. 2018, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ren, J.; Wang, F.; Pan, M.; Cui, L.; Li, M.; Qu, F. Mitochondrial and glucose metabolic dysfunctions in granulosa cells induce impaired oocytes of polycystic ovary syndrome through Sirtuin 3. Free Radic. Biol. Med. 2022, 187, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, W.; Xiang, Y.; Wang, S.; Wan, S.; Zhu, Y. Aberrant elevation of GDF8 impairs granulosa cell glucose metabolism via upregulating SERPINE1 expression in patients with PCOS. Mol. Ther. Nucleic Acids 2021, 23, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Cheng, Y.; Zhong, X.; Yang, D.; Jiang, S.; Ye, Y.; Ding, M.; Guan, G.; Yang, D.; Zhao, X. LNK promotes granulosa cell apoptosis in PCOS via negatively regulating insulin-stimulated AKT-FOXO3 pathway. Aging 2021, 13, 4617–4633. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yao, Y.; Xu, L.; Wu, H.; Wang, W.; He, Y.; Wang, Y.; Lu, Y.; Qi, J.; Ding, Y.; et al. Elevated SAA1 promotes the development of insulin resistance in ovarian granulosa cells in polycystic ovary syndrome. Reprod. Biol. Endocrinol. 2022, 20, 4. [Google Scholar] [CrossRef]
- Jiang, Q.; Pan, Y.; Li, P.; Zheng, Y.; Bian, Y.; Wang, W.; Wu, G.; Song, T.; Shi, Y. ANGPTL4 Expression in Ovarian Granulosa Cells Is Associated With Polycystic Ovary Syndrome. Front. Endocrinol 2021, 12, 799833. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Q.; Wang, W.; Qi, J.; He, Y.; Wang, Y.; Lu, Y.; Wu, H.; Ding, Y.; Sun, Y. Elevated chemerin induces insulin resistance in human granulosa-lutein cells from polycystic ovary syndrome patients. FASEB J. 2019, 33, 11303–11313. [Google Scholar] [CrossRef]
- Xu, A.; Fan, Y.; Liu, S.; Sheng, L.; Sun, Y.; Yang, H. GIMAP7 induces oxidative stress and apoptosis of ovarian granulosa cells in polycystic ovary syndrome by inhibiting sonic hedgehog signalling pathway. J. Ovarian Res. 2022, 15, 141. [Google Scholar] [CrossRef]
- Wan, T.; Sun, H.; Mao, Z.; Zhang, L.; Chen, X.; Shi, Y.; Shang, Y. Vitamin D deficiency inhibits microRNA-196b-5p which regulates ovarian granulosa cell hormone synthesis, proliferation, and apoptosis by targeting RDX and LRRC17. Ann. Transl. Med. 2021, 9, 1775. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, J.; Geng, X.; Chu, W.; Li, S.; Chen, Z.J.; Du, Y. Long non-coding RNA lnc-CCNL1-3:1 promotes granulosa cell apoptosis and suppresses glucose uptake in women with polycystic ovary syndrome. Mol. Ther. Nucleic Acids 2021, 23, 614–628. [Google Scholar] [CrossRef]
- Vitale, S.G.; Fulghesu, A.M.; Mikus, M.; Watrowski, R.; D’Alterio, M.N.; Lin, L.T.; Shah, M.; Reyes-Munoz, E.; Sathyapalan, T.; Angioni, S. The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside-A Systematic Literature Review. Biomedicines 2022, 10, 1816. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Qi, X.; Yun, C.; Qiao, J.; Pang, Y. Effects of Androgen Excess-Related Metabolic Disturbances on Granulosa Cell Function and Follicular Development. Front. Endocrinol. 2022, 13, 815968. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Ye, Z.; Mu, L.; Zhang, Y.; Long, X.; Zhang, C.; Li, R.; Zhao, Y.; Qiao, J. Elevated Levels of Follicular Fatty Acids Induce Ovarian Inflammation via ERK1/2 and Inflammasome Activation in PCOS. J. Clin. Endocrinol. Metab. 2022, 107, 2307–2317. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zuo, R.; He, Y.; Wang, Y.; Chen, Z.J.; Sun, Y.; Sun, K. Local Regeneration of Cortisol by 11beta-HSD1 Contributes to Insulin Resistance of the Granulosa Cells in PCOS. J. Clin. Endocrinol. Metab. 2016, 101, 2168–2177. [Google Scholar] [CrossRef]
- Wu, H.; Zhu, Q.; Li, X.; He, Y.; Wang, Y.; Lu, Y.; Qi, J.; Ding, Y.; Li, J.; Xu, R.; et al. The upregulation of 11beta-HSD1 in ovarian granulosa cells by cortisol and interleukin-1beta in polycystic ovary syndrome. Gynecol. Endocrinol. 2020, 36, 668–672. [Google Scholar] [CrossRef]
- Unfer, V.; Dinicola, S.; Lagana, A.S.; Bizzarri, M. Altered Ovarian Inositol Ratios May Account for Pathological Steroidogenesis in PCOS. Int. J. Mol. Sci. 2020, 21, 7157. [Google Scholar] [CrossRef]
- Sayutti, N.; Abu, M.A.; Ahmad, M.F. PCOS and Role of Cumulus Gene Expression in Assessing Oocytes Quality. Front. Endocrinol. 2022, 13, 843867. [Google Scholar] [CrossRef]
- Chu, W.; Li, S.; Geng, X.; Wang, D.; Zhai, J.; Lu, G.; Chan, W.Y.; Chen, Z.J.; Du, Y. Long-term environmental exposure of darkness induces hyperandrogenism in PCOS via melatonin receptor 1A and aromatase reduction. Front. Cell Dev. Biol. 2022, 10, 954186. [Google Scholar] [CrossRef]
- Guo, R.; Zheng, H.; Li, Q.; Qiu, X.; Zhang, J.; Cheng, Z. Melatonin alleviates insulin resistance through the PI3K/AKT signaling pathway in ovary granulosa cells of polycystic ovary syndrome. Reprod. Biol. 2022, 22, 100594. [Google Scholar] [CrossRef]
- de Medeiros, S.F.; Rodgers, R.J.; Norman, R.J. Adipocyte and steroidogenic cell cross-talk in polycystic ovary syndrome. Hum. Reprod. Update 2021, 27, 771–796. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, Q.; Dang, X.; He, Y.; Li, X.; Sun, Y. Local effect of bisphenol A on the estradiol synthesis of ovarian granulosa cells from PCOS. Gynecol. Endocrinol. 2017, 33, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ruan, Y.C.; Yang, Y.J.; Wang, K.; Liang, S.S.; Han, Y.B.; Teng, X.M.; Yang, J.Z. Follicular hyperandrogenism downregulates aromatase in luteinized granulosa cells in polycystic ovary syndrome women. Reproduction 2015, 150, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Froment, P.; Plotton, I.; Giulivi, C.; Fabre, S.; Khoueiry, R.; Mourad, N.I.; Horman, S.; Rame, C.; Rouillon, C.; Grandhaye, J.; et al. At the crossroads of fertility and metabolism: The importance of AMPK-dependent signaling in female infertility associated with hyperandrogenism. Hum. Reprod. 2022, 37, 1207–1228. [Google Scholar] [CrossRef] [PubMed]
- Estienne, A.; Mellouk, N.; Bongrani, A.; Plotton, I.; Langer, I.; Rame, C.; Petit, C.; Guerif, F.; Froment, P.; Dupont, J. Involvement of chemerin and CMKLR1 in the progesterone decrease by PCOS granulosa cells. Reproduction 2021, 162, 427–436. [Google Scholar] [CrossRef]
- Qi, J.; Li, J.; Wang, Y.; Wang, W.; Zhu, Q.; He, Y.; Lu, Y.; Wu, H.; Li, X.; Zhu, Z.; et al. Novel role of CXCL14 in modulating STAR expression in luteinized granulosa cells: Implication for progesterone synthesis in people with PCOS. Transl. Res. 2021, 230, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Baddela, V.S.; Sharma, A.; Vanselow, J. Non-esterified fatty acids in the ovary: Friends or foes? Reprod. Biol. Endocrinol. 2020, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Weng, X.; Tian, Y.; Wang, C.; Yang, Y.; Xu, K.; Zhang, C. Effects of omega-3 polyunsaturated fatty acids on steroidogenesis and cellular development in PCOS rats. Food Funct. 2019, 10, 2504–2514. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Wang, M.; Ponikwicka-Tyszko, D.; Ma, W.; Krentowska, A.; Kowalska, I.; Huhtaniemi, I.; Wolczynski, S.; Rahman, N.A.; et al. Role and mechanism of miR-335-5p in the pathogenesis and treatment of polycystic ovary syndrome. Transl. Res. 2023, 252, 64–78. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.D.; Zhou, X.Y.; Chen, S.L.; Chen, X.; Zhe, J.; Zhang, J.; Zhang, Q.Y.; Chen, Y.X. MiR-29a regulates the proliferation, aromatase expression, and estradiol biosynthesis of human granulosa cells in polycystic ovary syndrome. Mol. Cell. Endocrinol. 2019, 498, 110540. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wang, H.; Yan, C.Y.; Gao, X.F.; Ling, X.J. Deregulation of RUNX2 by miR-320a deficiency impairs steroidogenesis in cumulus granulosa cells from polycystic ovary syndrome (PCOS) patients. Biochem. Biophys. Res. Commun. 2017, 482, 1469–1476. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Y.; Lv, M.; Xing, Q.; Zhang, Z.; He, X.; Xu, Y.; Wei, Z.; Cao, Y. miR-323-3p regulates the steroidogenesis and cell apoptosis in polycystic ovary syndrome (PCOS) by targeting IGF-1. Gene 2019, 683, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, M.; Sun, J.; Jia, L.; Ma, S.; Gao, J.; Xu, Y.; Zhang, H.; Tsang, S.Y.; Li, X. MicroRNA-27a-3p affects estradiol and androgen imbalance by targeting Creb1 in the granulosa cells in mouse polycytic ovary syndrome model. Reprod. Biol. 2017, 17, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Van Hoeck, V.; Sturmey, R.G.; Bermejo-Alvarez, P.; Rizos, D.; Gutierrez-Adan, A.; Leese, H.J.; Bols, P.E.; Leroy, J.L. Elevated non-esterified fatty acid concentrations during bovine oocyte maturation compromise early embryo physiology. PLoS ONE 2011, 6, e23183. [Google Scholar] [CrossRef] [PubMed]
- Paczkowski, M.; Schoolcraft, W.B.; Krisher, R.L. Fatty acid metabolism during maturation affects glucose uptake and is essential to oocyte competence. Reproduction 2014, 148, 429–439. [Google Scholar] [CrossRef]
- De Bie, J.; Marei, W.F.A.; Maillo, V.; Jordaens, L.; Gutierrez-Adan, A.; Bols, P.E.J.; Leroy, J. Differential effects of high and low glucose concentrations during lipolysis-like conditions on bovine in vitro oocyte quality, metabolism and subsequent embryo development. Reprod. Fertil. Dev. 2017, 29, 2284–2300. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.-H.; Liu, X.-Y.; Wang, J. Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome. Int. J. Mol. Sci. 2023, 24, 16247. https://doi.org/10.3390/ijms242216247
Zhang C-H, Liu X-Y, Wang J. Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome. International Journal of Molecular Sciences. 2023; 24(22):16247. https://doi.org/10.3390/ijms242216247
Chicago/Turabian StyleZhang, Chen-Hua, Xiang-Yi Liu, and Jing Wang. 2023. "Essential Role of Granulosa Cell Glucose and Lipid Metabolism on Oocytes and the Potential Metabolic Imbalance in Polycystic Ovary Syndrome" International Journal of Molecular Sciences 24, no. 22: 16247. https://doi.org/10.3390/ijms242216247