
Genomic Analysis of Romanian Lycium Genotypes: Exploring BODYGUARD Genes for Stress Resistance Breeding


Abstract
:1. Introduction
2. Results
2.1. NGS Data Analysis
2.1.1. Sequencing Data Quality Control
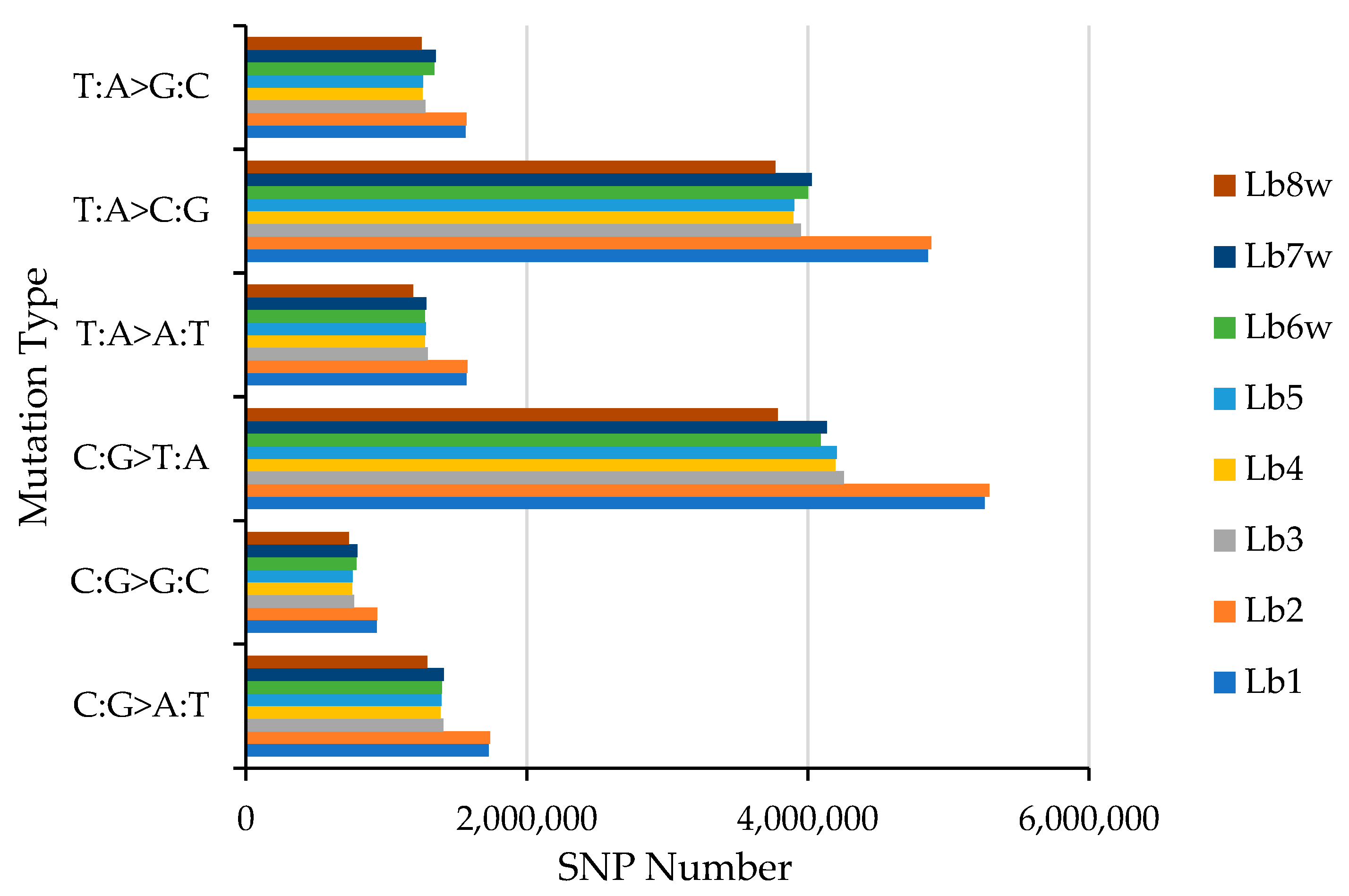
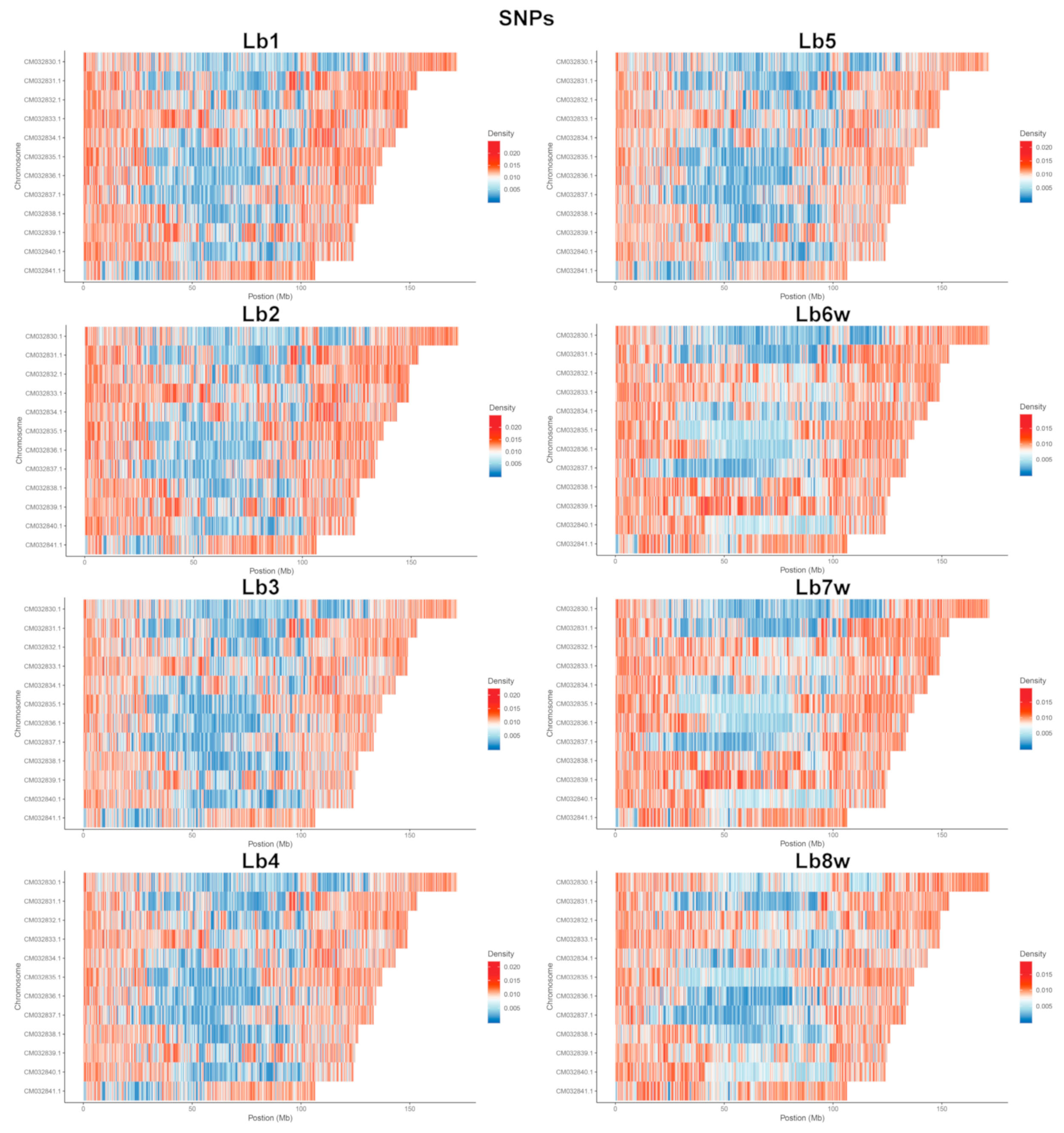
2.1.2. SNP Detection, Distribution, and Mutation Frequency
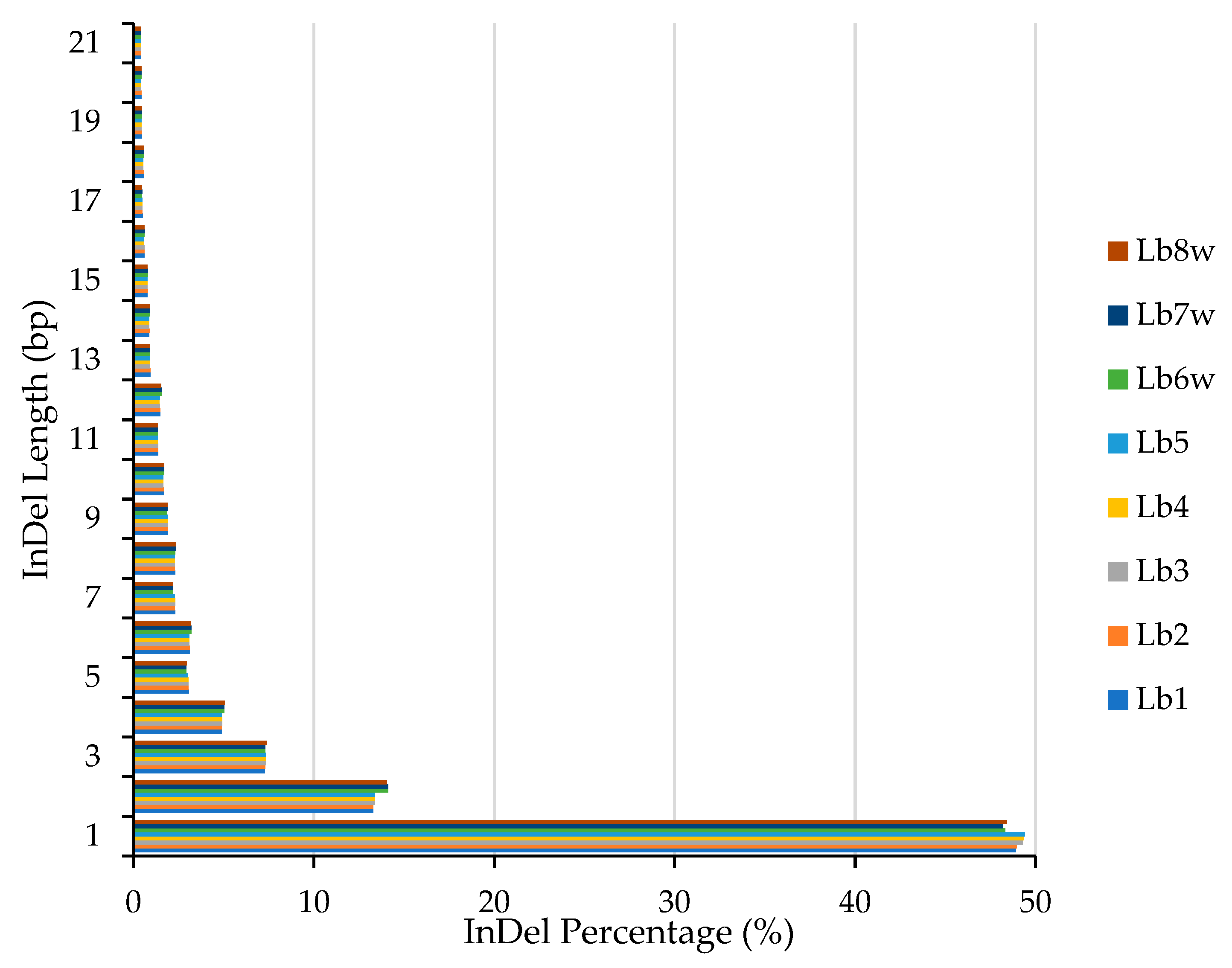
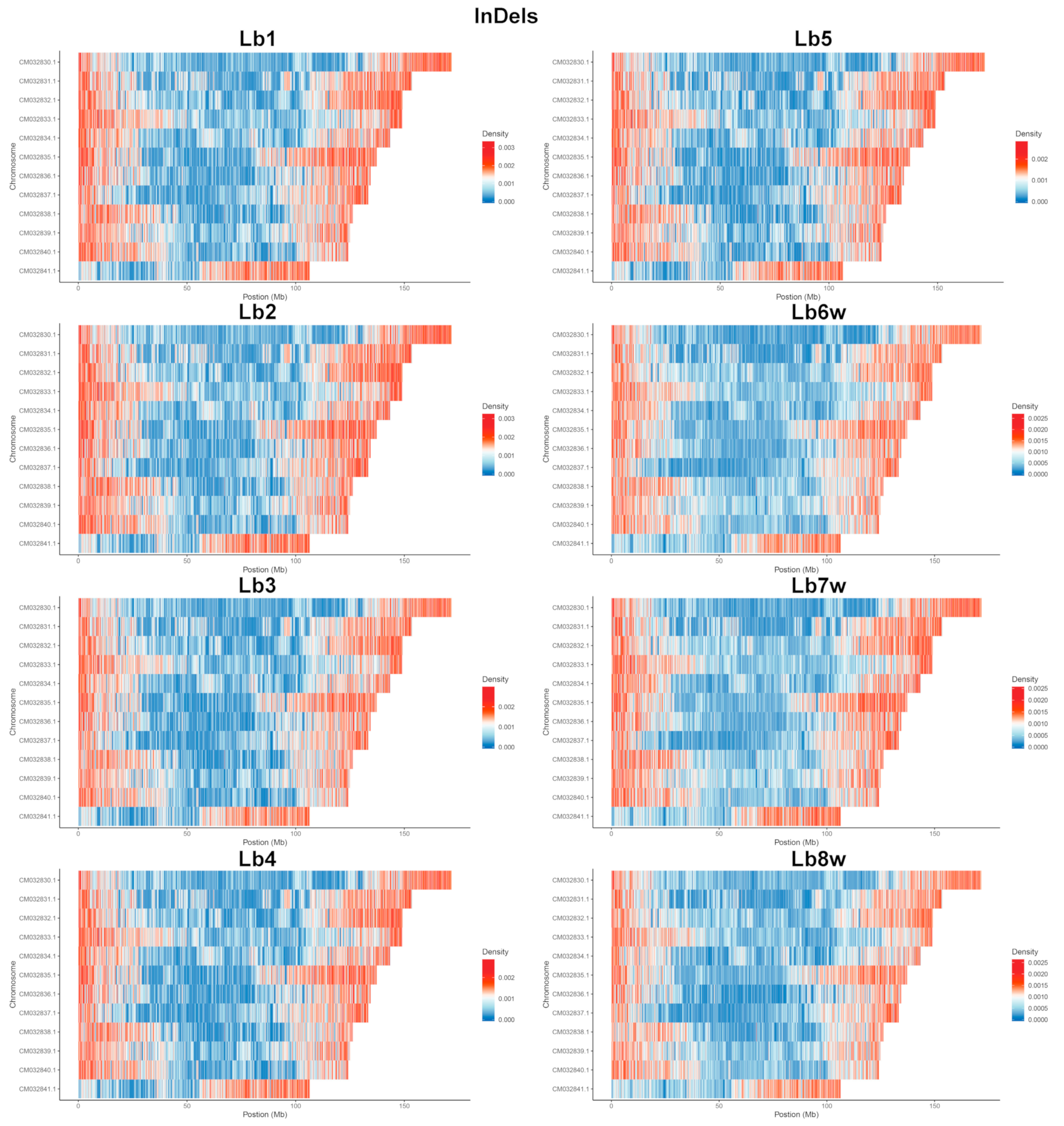
2.1.3. Insertion/Deletion Detection and Distribution
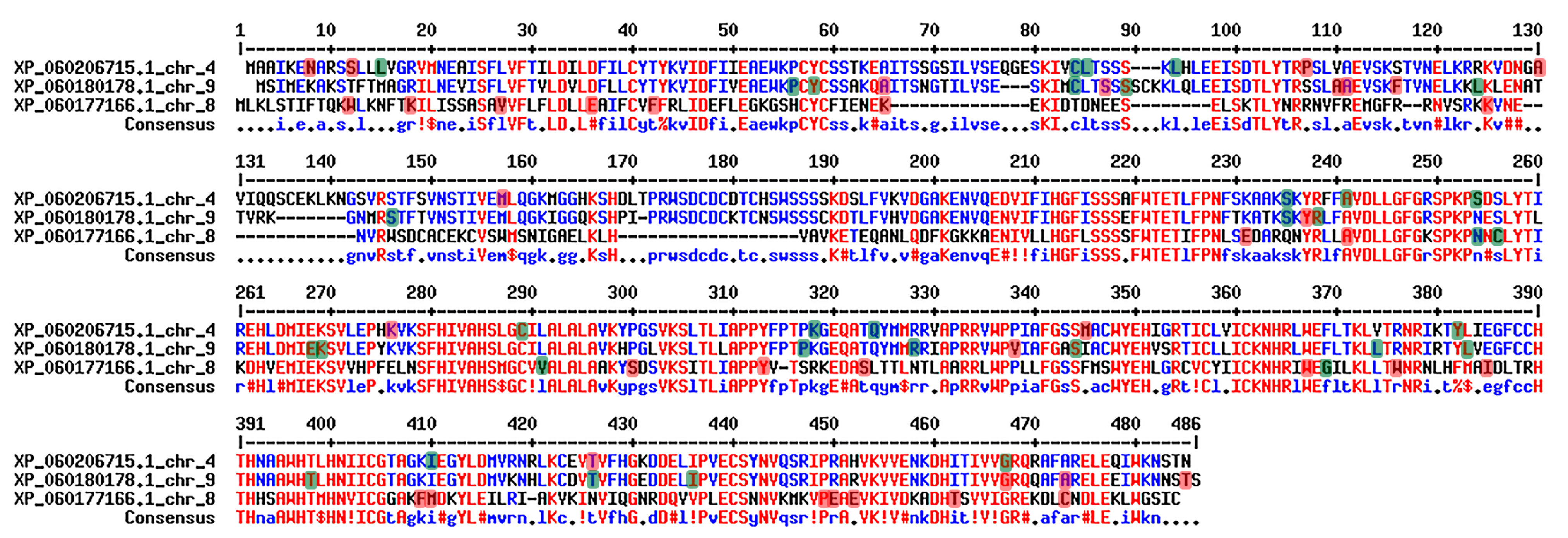
2.1.4. Sequence Analyses of BODYGUARD Genes in Romanian Goji Berry Genomes
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction
4.3. Sequencing and Sequencing Data Quality Control
4.4. Computational Data Processing and Sequencing Analysis
4.5. SNP Detection and Annotation
4.6. Insertion/Deletion (InDel) Detection and Annotation
4.7. Sequence Analysis of the BDG Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Guo, S.; Zhang, F.; Yan, H.; Qian, D.; Shang, E.; Wang, H.; Duan, J. Nutritional Components Characterization of Goji Berries from Different Regions in China. J. Pharm. Biomed. Anal. 2021, 195, 113859. [Google Scholar] [CrossRef] [PubMed]
- Vidović, B.B.; Milinčić, D.D.; Marčetić, M.D.; Djuriš, J.D.; Ilić, T.D.; Kostić, A.Ž.; Pešić, M.B. Health Benefits and Applications of Goji Berries in Functional Food Products Development: A Review. Antioxidants 2022, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, Antioxidant Efficacies, and Health Effects—A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1580–1604. [Google Scholar] [CrossRef] [PubMed]
- Wetters, S.; Horn, T.; Nick, P. Goji Who? Morphological and DNA Based Authentication of a “Superfood”. Front. Plant Sci. 2018, 9, 1859. [Google Scholar] [CrossRef] [PubMed]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, Pharmacology and Safety in the Perspective of Traditional Uses and Recent Popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef]
- Zordan, A. Italian-Grown Fresh Goji Berries, Here’s Where. Available online: https://www.gamberorossointernational.com/news/italian-grown-fresh-goji-berries-here-s-where/ (accessed on 12 November 2023).
- Skenderidis, P.; Leontopoulos, S.; Lampakis, D. Goji Berry: Health Promoting Properties. Nutraceuticals 2022, 2, 32–48. [Google Scholar] [CrossRef]
- Teixeira, F.; Silva, A.M.; Delerue-Matos, C.; Rodrigues, F. Lycium barbarum Berries (Solanaceae) as Source of Bioactive Compounds for Healthy Purposes: A Review. Int. J. Mol. Sci. 2023, 24, 4777. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Q.; Fang, J.; Wang, C.; Wang, D.; Li, M. The Anti-Aging Activity of Lycium barbarum Polysaccharide Extracted by Yeast Fermentation: In Vivo and in Vitro Studies. Int. J. Biol. Macromol. 2022, 209, 2032–2041. [Google Scholar] [CrossRef]
- Yang, F.-L.; Wei, Y.-X.; Liao, B.-Y.; Wei, G.-J.; Qin, H.-M.; Pang, X.-X.; Wang, J.-L. Effects of Lycium barbarum Polysaccharide on Endoplasmic Reticulum Stress and Oxidative Stress in Obese Mice. Front. Pharmacol. 2020, 11, 742. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Xia, M.; Lan, J.; Yang, L.; Wang, Z.; Wang, R.; Tao, H.; Shi, Y. A Comprehensive Review on the Ethnobotany, Phytochemistry, Pharmacology and Quality Control of the Genus Lycium in China. Food Funct. 2023, 14, 2998–3025. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.-J.; Zheng, J.; Wu, J.; Qiao, H.-Q.; Chang, Q.; Niu, Y.; Sun, T.; Li, Y.-X.; Yu, J.-Q. Beneficial Effects of Lycium barbarum Polysaccharide on Spermatogenesis by Improving Antioxidant Activity and Inhibiting Apoptosis in Streptozotocin-Induced Diabetic Male Mice. Food Funct. 2017, 8, 1215–1226. [Google Scholar] [CrossRef]
- İLter Aktaş, G.; Firat, T.; PehliVan Karakaş, F. The Effect of Lycium barbarum on Reproductive System and the Expression of CRISP-1 Protein in Experimentally Diabetic Male Rats. J. Health Sci. Med. 2022, 5, 706–714. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Weckerle, C.S. The Genus Lycium as Food and Medicine: A Botanical, Ethnobotanical and Historical Review. J. Ethnopharmacol. 2018, 212, 50–66. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Zou, Y.; Reich, E.; Zhang, X.; Chen, Y.; Weckerle, C.S. Quality Variation of Goji (Fruits of Lycium spp.) in China: A Comparative Morphological and Metabolomic Analysis. Front. Pharmacol. 2018, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- WFO Plant List. World Flora Online. Available online: https://wfoplantlist.org/plant-list/taxon/wfo-4000022495-2023-06?page=1 (accessed on 12 November 2023).
- Lycium L. Plants of the World Online. Kew Science. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:30001330-2 (accessed on 12 November 2023).
- Gao, X.; Li, J.; Song, J.; Guo, Q. The SSR Genetic Diversity of Wild Red Fruit Lycium (Lycium barbarum) in Northwest China. Forests 2023, 14, 1598. [Google Scholar] [CrossRef]
- Ciocârlan, V. Illustrated Flora of Romania: Pteridophyta et Spermatophyta. [Flora ilustrată a României: Pteridophyta et Spermatophyta]; Ceres: Bucharest, Romania, 2009; ISBN 978-973-40-0817-9. (In Romanian) [Google Scholar]
- Lycium halimifolium Mill. Available online: https://www.worldfloraonline.org/taxon/wfo-0001022945 (accessed on 12 November 2023).
- Marian, S.F. Romanian Folk Botany. [Botanica Poporana Romana]; Academiei: Bucharest, Romania, 2008; ISBN 978-973-1974-08-8. (In Romanian) [Google Scholar]
- Lycium vulgare Dunal. Available online: https://www.worldfloraonline.org/taxon/wfo-0001023262 (accessed on 12 November 2023).
- Branişte, N.; Budan, S.; Butac, M.; Militaru, M. Fruit Tree, Small Fruits and Strawberry Cultivars Released in Romania. [Soiuri de Pomi Arbusti Fructiferi Si Capsuni Create in Romania]; Paralela 45: Bucharest, Romania, 2007; ISBN 978-973-47-0177-3. [Google Scholar]
- Lycium barbarum L.—Trees and Shrubs Online. Available online: https://www.treesandshrubsonline.org/articles/lycium/lycium-barbarum/ (accessed on 14 November 2023).
- Agro, D. Attention!!! Confusion between Goji and Catina de Garduri or Gardurarita. [Atentie !!! Confuzie Intre Goji si Catina de Garduri sau Gardurarita]. Available online: https://www.agrodenmar.ro/articole/goji-sau-catina-de-garduri (accessed on 12 November 2023).
- Agrointeligenta How Do You Tell the Difference between True Goji and Toxic Fruited Goji Berry! [Cum faci Diferența Dintre Goji Adevărat și Cătina de Gard cu Fructe Toxice!]. Available online: https://agrointel.ro/79208/cum-faci-diferenta-dintre-goji-adevarat-si-catina-de-gard-cu-fructe-toxice/ (accessed on 12 November 2023).
- Petran, M.; Dragos, D.; Gilca, M. Historical Ethnobotanical Review of Medicinal Plants Used to Treat Children Diseases in Romania (1860s–1970s). J. Ethnobiol. Ethnomedicine 2020, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Luchian, V.; Ciceoi, R.; Gutue, M. Comparative Leaf and Flower Morpho-Anatomical Study of Wild and Cultivated Gojiberry (Lycium barbarum L.) in Romania. Sci. Pap. Ser. B Hortic. 2022, LXVI, 821–829. [Google Scholar]
- Răduțoiu, D.; Bãloniu, L. Invasive and Potentially Invasive Alogen Plants in the Agricultural Crops of Oltenia. Sci. Pap. Ser. B Hortic. 2021, LXV, 782–787. [Google Scholar]
- Yao, R.; Heinrich, M.; Wang, Z.; Weckerle, C.S. Quality Control of Goji (Fruits of Lycium barbarum L. and L. chinense Mill.): A Value Chain Analysis Perspective. J. Ethnopharmacol. 2018, 224, 349–358. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zhu, L.; Wang, Y. Nuclear Genome Size Estimation and Karyotype Analysis of Lycium Species (Solanaceae). Sci. Hortic. 2013, 151, 46–50. [Google Scholar] [CrossRef]
- Lv, W.; Zhao, N.; Zhao, Q.; Huang, S.; Liu, D.; Wang, Z.; Yang, J.; Zhang, X. Discovery and Validation of Biomarkers for Zhongning Goji Berries Using Liquid Chromatography Mass Spectrometry. J. Chromatogr. B 2020, 1142, 122037. [Google Scholar] [CrossRef]
- Gong, H.; Rehman, F.; Ma, Y.; Biao, A.; Zeng, S.; Yang, T.; Huang, J.; Li, Z.; Wu, D.; Wang, Y. Germplasm Resources and Strategy for Genetic Breeding of Lycium Species: A Review. Front. Plant Sci. 2022, 13, 802936. [Google Scholar] [CrossRef]
- Goji Berries Market. Available online: https://www.transparencymarketresearch.com/goji-berries-market.html (accessed on 14 November 2023).
- Chen, J.; Chao, C.T.; Wei, X.; Chen, J.; Chao, C.T.; Wei, X. Gojiberry Breeding: Current Status and Future Prospects. In Breeding and Health Benefits of Fruit and Nut Crops; IntechOpen: Rijeka, Croatia, 2018; ISBN 978-1-78923-273-8. [Google Scholar] [CrossRef]
- Beigel, S. Inspirational Ideas: European Goji Berries. Available online: https://ec.europa.eu/eip/agriculture/en/news/inspirational-ideas-european-goji-berries (accessed on 14 November 2023).
- Zhang, Y.; Qin, J.; Wang, Y.; Zhou, T.; Feng, N.; Ma, C.; Zhu, M. Levels and Health Risk Assessment of Pesticides and Metals in Lycium barbarum L. from Different Sources in Ningxia, China. Sci. Rep. 2022, 12, 561. [Google Scholar] [CrossRef]
- Buda, D. Parliamentary Question|The Largest Goji Berry Cultivation in Europe, in Satu Mare, May Be Forced to Cease Production. E-002842/2018. European Parliament. Available online: https://www.europarl.europa.eu/doceo/document/E-8-2018-002842_EN.html (accessed on 13 November 2023).
- Ciceoi, R.; Mardare, E.S. Aceria Kuko Mites: A Comprehensive Review of Their Phytosanitary Risk, Pathways and Control. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Hortic. 2016, 73, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Ciceoi, R.; Stavrescu-Bedivan, M.-M.; Luchian, V.; Stanica, F.; Venat, O.; Asănică, A. Goji berry cultivation in Romania, a pathway between traditional uses and modern breeding, cultivation, and citizens acceptance. Acta Hortic. 2023, 1381, 383–391. [Google Scholar] [CrossRef]
- Clapa, D.; Fira, A.; Borsai, O.; Hârța, M.; Sisea, C.R.; Dumitraş, A.F.; Pamfil, D. Lycium barbarum L.—A New Cultivated Species in Romania. Acta Hortic. 2021, 1308, 205–212. [Google Scholar] [CrossRef]
- Mörtl, M.; Ciceoi, R.; Ion, V.A.; Klátyik, S.; Székács, A. Environmental Concerns Regarding the Occurrence of Neonicotinoid Insecticides in Berry Fruits. Sci. Pap. Ser. B Hortic. 2022, LXVI, 399–406. [Google Scholar]
- Zhang, D.; Xia, T.; Dang, S.; Fan, G.; Wang, Z. Investigation of Chinese Wolfberry (Lycium spp.) Germplasm by Restriction Site-Associated DNA Sequencing (RAD-Seq). Biochem. Genet. 2018, 56, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Yokoyama, J.; Ohashi, H. Phylogeny and Biogeography of the Genus Lycium (Solanaceae): Inferences from Chloroplast DNA Sequences. Mol. Phylogenet. Evol. 2001, 19, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Polat, M.; Mertoglu, K.; Eskimez, I.; Okatan, V. Effects of the fruiting period and growing seasons on market quality in goji berry (Lycium barbarum L.). Folia Hortic. 2020, 32, 229–239. [Google Scholar] [CrossRef]
- Liu, J.; Shi, X.; Lin, H.; He, C.; Li, Q.; Shen, G.; Feng, J. Geographical Origin Identification and Quality Comparison of Ningxia Goji Berries (Lycium barbarum L.) by NMR-Based Techniques. J. Food Compos. Anal. 2023, 119, 105258. [Google Scholar] [CrossRef]
- Chao, H.; Zhang, S.; Hu, Y.; Ni, Q.; Xin, S.; Zhao, L.; Ivanisenko, V.A.; Orlov, Y.L.; Chen, M. Integrating Omics Databases for Enhanced Crop Breeding. J. Integr. Bioinforma. 2023, 20, 20230012. [Google Scholar] [CrossRef] [PubMed]
- Marks, R.A.; Hotaling, S.; Frandsen, P.B.; VanBuren, R. Representation and Participation across 20 Years of Plant Genome Sequencing. Nat. Plants 2021, 7, 1571–1578. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Yuan, Z.; Song, Z.; Wang, Z.; Gao, J.; Fu, W.; Zhang, G. Chromosome-Level Genome Assembly and Annotation of the Prickly Nightshade Solanum rostratum Dunal. Sci. Data 2023, 10, 341. [Google Scholar] [CrossRef] [PubMed]
- Gladman, N.; Goodwin, S.; Chougule, K.; Richard McCombie, W.; Ware, D. Era of Gapless Plant Genomes: Innovations in Sequencing and Mapping Technologies Revolutionize Genomics and Breeding. Curr. Opin. Biotechnol. 2023, 79, 102886. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-L.; Li, Y.; Fan, Y.-F.; Li, Z.; Yoshida, K.; Wang, J.-Y.; Ma, X.-K.; Wang, N.; Mitsuda, N.; Kotake, T.; et al. Wolfberry Genomes and the Evolution of Lycium (Solanaceae). Commun. Biol. 2021, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- CSIRO Data Access Portal—Chromosomal Level Assembly of African Boxthorn. Available online: https://data.csiro.au/collection/csiro:60003 (accessed on 12 November 2023).
- Lycium barbarum Isolate: ZL-2021 (ID 640228)—BioProject—NCBI. Available online: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA640228/ (accessed on 9 November 2023).
- Tudor, V.; As, A.; Ionu, Z.; Gîdea, M.; Veronica, J. Germination Capacity of Some Lycium barbarum L. and Lycium chinense Mill. Biotypes Seeds. Rom. Biotechnol. Lett. 2017, 22, 12191–12196. [Google Scholar]
- Stavrescu-Bedivan, M.-M.; Pelcaru, C.F.; Croitoru, C.M.; Mihai, C.D.; Ciceoi, R. Preliminary Survey for Mapping the Distribution of Spontaneous Goji Berry Shrubs in Romania. Sci. Pap. Ser. B Hortic. 2022, LXVI, 907–912. [Google Scholar]
- Asănica, A.; Tudor, V.; Teodorescu, R.I.; Iacob, A.; Zolotoi, V.; Tudor, A. Results on Hardwood Cuttings Propagation of Some Lycium sp. Genotypes. Fruit Grow. Res. 2016, XXXII, 63–70. [Google Scholar]
- Asănică, A.; Manole, C.; Tudor, V.; Dobre, A.; Teodorescu, R.I. Lycium barbarum L. Juice—Natural Source of Biologically Active Compounds. AgroLife Sci. J. 2016, 1, 15–20. [Google Scholar]
- Ciceoi, R.; Luchian, V.; Tabacu, A.F.; Gutue, M.; Stavrescu-Bedivan, M.M. Goji Berry Gall Mite Expansion in Europe, with Emphasis on Southeastern Part of Romania. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Food Sci. Technol. 2021, 78, 93. [Google Scholar] [CrossRef] [PubMed]
- Dzhugalov, H.; Lichev, V.; Yordanov, A.; Kaymakanov, P.; Dimitrova, V.; Kutoranov, G.; Dimitrova, V.; Kutoranov, G. First Results of Testing Goji Berry (Lycium barbarum L.) in Plovdiv Region, Bulgaria. Sci. Pap. Ser. B Hortic. 2015, LIX, 47–50. [Google Scholar]
- Chen, H.; Shen, S.; Zhi, H.; Li, W. Pesticides Residues on Goji Berry: A Characteristic Minor Crop in China. J. Food Compos. Anal. 2023, 120, 105342. [Google Scholar] [CrossRef]
- Alexandru, D. Drought Monitoring in Romania. In Proceedings of the Kick-Off of the Network of Drought Observatories in the EU, Brussels, Belgium, 17 July 2022. [Google Scholar]
- Drought Wipes EUR 1 Bln from Romanian Agricultural Sector. Available online: https://www.romania-insider.com/drought-wipes-money-romanian-agricultural-sector (accessed on 14 November 2023).
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, J.; Zhang, H.; Wang, C.; Zhao, L.; Huang, T.; Qing, K. Chemical Composition, Crystal Morphology, and Key Gene Expression of the Cuticular Waxes of Goji (Lycium barbarum L.) Berries. J. Agric. Food Chem. 2021, 69, 7874–7883. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Jiang, B.; Liu, R.; Han, Y.; Fang, X.; Mu, H.; Farag, M.A.; Simal-Gandara, J.; Prieto, M.A.; Chen, H.; et al. Structures and Functions of Cuticular Wax in Postharvest Fruit and Its Regulation: A Comprehensive Review with Future Perspectives. Engineering 2023, 23, 118–129. [Google Scholar] [CrossRef]
- Shaheenuzzamn, M.; Shi, S.; Sohail, K.; Wu, H.; Liu, T.; An, P.; Wang, Z.; Hasanuzzaman, M. Regulation of Cuticular Wax Biosynthesis in Plants under Abiotic Stress. Plant Biotechnol. Rep. 2021, 15, 1–12. [Google Scholar] [CrossRef]
- Jakobson, L.; Lindgren, L.O.; Verdier, G.; Laanemets, K.; Brosché, M.; Beisson, F.; Kollist, H. BODYGUARD Is Required for the Biosynthesis of Cutin in Arabidopsis. New Phytol. 2016, 211, 614–626. [Google Scholar] [CrossRef]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef]
- Simões, R.; Rodrigues, A.; Ferreira-Dias, S.; Miranda, I.; Pereira, H. Chemical Composition of Cuticular Waxes and Pigments and Morphology of Leaves of Quercus Suber Trees of Different Provenance. Plants 2020, 9, 1165. [Google Scholar] [CrossRef]
- Wang, X.; Kong, L.; Zhi, P.; Chang, C. Update on Cuticular Wax Biosynthesis and Its Roles in Plant Disease Resistance. Int. J. Mol. Sci. 2020, 21, 5514. [Google Scholar] [CrossRef]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf Life Potential and the Fruit Cuticle: The Unexpected Player. Front. Plant Sci. 2019, 10, 770. [Google Scholar] [CrossRef]
- Kurdyukov, S.; Faust, A.; Nawrath, C.; Bär, S.; Voisin, D.; Efremova, N.; Franke, R.; Schreiber, L.; Saedler, H.; Métraux, J.-P.; et al. The Epidermis-Specific Extracellular BODYGUARD Controls Cuticle Development and Morphogenesis in Arabidopsis. Plant Cell 2006, 18, 321–339. [Google Scholar] [CrossRef]
- Aragón, W.; Formey, D.; Aviles-Baltazar, N.Y.; Torres, M.; Serrano, M. Arabidopsis thaliana Cuticle Composition Contributes to Differential Defense Response to Botrytis Cinerea. Front. Plant Sci. 2021, 12, 738949. [Google Scholar] [CrossRef]
- Panikashvili, D.; Shi, J.X.; Bocobza, S.; Franke, R.B.; Schreiber, L.; Aharoni, A. The Arabidopsis DSO/ABCG11 Transporter Affects Cutin Metabolism in Reproductive Organs and Suberin in Roots. Mol. Plant 2010, 3, 563–575. [Google Scholar] [CrossRef]
- Evans, J.; McCormick, R.F.; Morishige, D.; Olson, S.N.; Weers, B.; Hilley, J.; Klein, P.; Rooney, W.; Mullet, J. Extensive Variation in the Density and Distribution of DNA Polymorphism in Sorghum Genomes. PLoS ONE 2013, 8, e79192. [Google Scholar] [CrossRef]
- Bekele, W.A.; Wieckhorst, S.; Friedt, W.; Snowdon, R.J. High-Throughput Genomics in Sorghum: From Whole-Genome Resequencing to a SNP Screening Array. Plant Biotechnol. J. 2013, 11, 1112–1125. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nagasaki, H.; Garcia, V.; Just, D.; Bres, C.; Mauxion, J.-P.; Le Paslier, M.-C.; Brunel, D.; Suda, K.; Minakuchi, Y.; et al. Genome-Wide Analysis of Intraspecific DNA Polymorphism in ‘Micro-Tom’, a Model Cultivar of Tomato (Solanum lycopersicum). Plant Cell Physiol. 2014, 55, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Udriște, A.-A.; Iordachescu, M.; Ciceoi, R.; Bădulescu, L. Next-Generation Sequencing of Local Romanian Tomato Varieties and Bioinformatics Analysis of the Ve Locus. Int. J. Mol. Sci. 2022, 23, 9750. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Lee, H.-Y.; Ro, N.-Y.; Hur, O.-S.; Lee, J.-H.; Kwon, J.-K.; Kang, B.-C. QTL Mapping and GWAS Reveal Candidate Genes Controlling Capsaicinoid Content in Capsicum. Plant Biotechnol. J. 2018, 16, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.-C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; Deynze, A.V.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a Large SNP Genotyping Array and Generation of High-Density Genetic Maps in Tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef] [PubMed]
- Tabăra, M. The anatomical structure of Lycium barbarum L. leaf lamina from spontaneous flora and cultivated varieties. Akademos 2020. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Zhi, P.; Wang, X.; Xu, B.; Gong, Z.; Chang, C. Origins and Evolution of Cuticle Biosynthetic Machinery in Land Plants1 [OPEN]. Plant Physiol. 2020, 184, 1998–2010. [Google Scholar] [CrossRef] [PubMed]
- Chassot, C.; Nawrath, C.; Métraux, J.-P. Cuticular Defects Lead to Full Immunity to a Major Plant Pathogen. Plant J. 2007, 49, 972–980. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, D.R.; Deak, K.I.; Ingram, P.A.; Malamy, J.E. Root System Architecture in Arabidopsis Grown in Culture Is Regulated by Sucrose Uptake in the Aerial Tissues. Plant Cell 2008, 20, 2643–2660. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Xiong, L.; Li, W.; Zhu, J.-K.; Zhu, J. The Plant Cuticle Is Required for Osmotic Stress Regulation of Abscisic Acid Biosynthesis and Osmotic Stress Tolerance in Arabidopsis. Plant Cell 2011, 23, 1971–1984. [Google Scholar] [CrossRef]
- Qian, D.; Ji, R.-F.; Gao, W.; Huang, L.-Q. Advances in research on relationships among Lycium species and origin of cultivated Lycium in China. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 2017, 42, 3282–3285. [Google Scholar] [CrossRef]
- Rehman, F.; Gong, H.; Li, Z.; Zeng, S.; Yang, T.; Ai, P.; Pan, L.; Huang, H.; Wang, Y. Identification of Fruit Size Associated Quantitative Trait Loci Featuring SLAF Based High-Density Linkage Map of Goji Berry (Lycium spp.). BMC Plant Biol. 2020, 20, 474. [Google Scholar] [CrossRef]
- Lam, H.-M.; Xu, X.; Liu, X.; Chen, W.; Yang, G.; Wong, F.-L.; Li, M.-W.; He, W.; Qin, N.; Wang, B.; et al. Resequencing of 31 Wild and Cultivated Soybean Genomes Identifies Patterns of Genetic Diversity and Selection. Nat. Genet. 2010, 42, 1053–1059. [Google Scholar] [CrossRef]
- Lee, Y.-G.; Jeong, N.; Kim, J.H.; Lee, K.; Kim, K.H.; Pirani, A.; Ha, B.-K.; Kang, S.-T.; Park, B.-S.; Moon, J.-K.; et al. Development, Validation and Genetic Analysis of a Large Soybean SNP Genotyping Array. Plant J. 2015, 81, 625–636. [Google Scholar] [CrossRef]
- Pandey, M.K.; Agarwal, G.; Kale, S.M.; Clevenger, J.; Nayak, S.N.; Sriswathi, M.; Chitikineni, A.; Chavarro, C.; Chen, X.; Upadhyaya, H.D.; et al. Development and Evaluation of a High Density Genotyping ‘Axiom_Arachis’ Array with 58 K SNPs for Accelerating Genetics and Breeding in Groundnut. Sci. Rep. 2017, 7, 40577. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Zamir, D. A Genomic Library of Lycopersicon pennellii in L. Esculentum: A Tool for Fine Mapping of Genes. Euphytica 1994, 79, 175–179. [Google Scholar] [CrossRef]
- Ma, B.; Liao, L.; Peng, Q.; Fang, T.; Zhou, H.; Korban, S.S.; Han, Y. Reduced Representation Genome Sequencing Reveals Patterns of Genetic Diversity and Selection in Apple. J. Integr. Plant Biol. 2017, 59, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.A.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ File Format for Sequences with Quality Scores, and the Solexa/Illumina FASTQ Variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Cacho, A.; Smirnova, E.; Huzurbazar, S.; Cui, X. A Comparison of Base-Calling Algorithms for Illumina Sequencing Technology. Brief. Bioinform. 2016, 17, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Illumina, Inc. Quality Scores for Next-Generation Sequencing 2011; Illumina, Inc.: San Diego, CA, USA, 2011. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup the Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional Annotation of Genetic Variants from High-Throughput Sequencing Data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Kuznetsov, A.; Bollin, C.J. NCBI Genome Workbench: Desktop Software for Comparative Genomics, Visualization, and GenBank Data Submission. Methods Mol. Biol. 2021, 2231, 261–295. [Google Scholar] [CrossRef]
- Corpet, F. Multiple Sequence Alignment with Hierarchical Clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name/Gene ID | Description | Location |
|---|---|---|
| LOC132634709 ID: 132634709 | probable lysophospholipase BODYGUARD 3 [Lycium barbarum (goji berry)] | Chromosome 4, NC_083340.1 |
| LOC132607278 ID: 132607278 | probable lysophospholipase BODYGUARD 4 [Lycium barbarum (goji berry)] | Chromosome 8, NC_083344.1 |
| LOC132609965 ID: 132609965 | probable lysophospholipase BODYGUARD 3 [Lycium barbarum (goji berry)] | Chromosome 9, NC_083345.1 |
| LOC132060388 ID: 132060388 | probable lysophospholipase BODYGUARD 3 [Lycium ferocissimum] | Chromosome 1, NC_081342.1 |
| LOC132049371 ID: 132049371 | probable lysophospholipase BODYGUARD 4 [Lycium ferocissimum] | Chromosome 3, NC_081344.1 |
| LOC132030714 ID: 132030714 | probable lysophospholipase BODYGUARD 3 [Lycium ferocissimum] | Chromosome 9, NC_081350.1 |
| Nr. crt. | SNP Position in Coding Sequence | Codon | Amino Acid | Reference Genome ASM1917538v2 | Cultivated Genotypes | Wild Genotypes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lb1 | Lb2 | Lb3 | Lb4 | Lb5 | Lb6w | Lb7w | Lb8w | |||||
| 1 | 517 | AAT/AGT | Asn/Ser | Asn | h69 | h59 | h67 | h50 | h75 | Asn | Asn | Asn |
| 2 | 529 | AGC/ATC | Ser/Ile | Ser | h69 | h57 | h40 | h50 | h75 | Ser | Ser | Ser |
| 3 | 537 | TTA/CTA | Leu/Leu (silent) | Leu (TTA) | Leu (CTA) | Leu (CTA) | Leu (CTA) | Leu (CTA) | Leu (CTA) | h33 | h20 | h30 |
| 4 | 590 | CTT/CTC | Leu/Leu (silent) | Leu (CTT) | Leu (CTC) | Leu (CTC) | Leu (CTC) | Leu (CTC) | Leu (CTC) | h37 | Leu (CTT) | h40 |
| 5 | 746 | TGC/TGT | Cys/Cys (silent) | Cys (TGC) | Cys (TGT) | Cys (TGT) | Cys (TGT) | Cys (TGT) | Cys (TGT) | h50 | h17 | h30 |
| 6 | 749 | CTG/CTC | Leu/Leu (silent) | Leu (CTG) | h35 | h40 | h50 | h33 | h20 | h43 | h29 | h36 |
| 7 | 767 | CTA/CTG | Leu/Leu (silent) | Leu (CTA) | Leu (CTG) | Leu (CTG) | Leu (CTG) | Leu (CTG) | Leu (CTG) | h50 | h37 | h30 |
| 8 | 804 | CCA/ACA | Pro/Thr | Pro | h78 | h45 | h25 | h80 | h60 | Pro | Pro | Pro |
| 9 | 873 | GCT/TCT | Ala/Ser | Ala | h67 | h56 | h18 | h60 | h80 | Ala | Ala | Ala |
| 10 | 956 | ATG/ATT | Met/Ile | Met | h42 | h35 | h37 | h40 | h80 | Met | Met | Met |
| 11 | 1190 | TCA/TCT | Ser/Ser (silent) | Ser (TCA) | h46 | h92 | h50 | h45 | h60 | Ser (TCA) | Ser (TCA) | Ser (TCA) |
| 12 | 1208 | GCA/GCC | Ala/Ala (silent) | Ala (GCA) | h50 | h7 | h50 | h60 | h40 | h75 | h40 | h33 |
| 13 | 1247 | AGT/AGC | Ser/Ser (silent) | Ser (AGT) | h50 | h20 | h57 | h70 | h25 | h80 | h45 | h40 |
| 14 | 1312 | AAA/AGA | Lys/Arg | Lys | Arg | Arg | Arg | Arg | Arg | h80 | h40 | h50 |
| 15 | 1352 | TGC/TGT | Cys/Cys (silent) | Cys(TGC) | h42 | h39 | h53 | h57 | h44 | Cys (TGC) | Cys (TGC) | h14 |
| 16 | 1439 | AAA/AAG | Lys/Lys (silent) | Gly | h59 | h65 | h60 | h18 | h60 | h67 | h57 | h57 |
| 17 | 1457 | CAG/CAA | Gln/Gln (silent) | Gln (CAG) | Gln (CAG) | Gln (CAG) | Gln (CAG) | Gln (CAG) | Gln (CAG) | h67 | h57 | h57 |
| 18 | 1520 | ATG/ATT | Met/Ile | Met | h43 | h54 | h67 | h83 | h45 | Met | Met | Met |
| 19 | 1631 | TAC/TAT | Tyr/Tyr (silent) | Tyr (TAC) | Tyr (TAT) | Tyr (TAT) | Tyr (TAT) | Tyr (TAT) | Tyr (TAT) | h75 | h25 | h73 |
| 20 | 1715 | ATA/ATT | Ile/Ile (silent) | Ile | h61 | h47 | h50 | h50 | h50 | h67 | h33 | h60 |
| 21 | 1761 | ACG/TCG | Thr/Ser | Tyr | h67 | h71 | h50 | h37 | h33 | Tyr | Tyr | Tyr |
| 22 | 1886 | GGC/GGG | Gly/Gly (silent) | Gly (GGC) | Gly (GGC) | h36 | h54 | h86 | h30 | h83 | h43 | h75 |
| Nr. crt. | SNP Position in Coding Sequence | Codon | Amino Acid | Reference Genome ASM1917538v2 | Cultivated Genotypes | Wild Genotypes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lb1 | Lb2 | Lb3 | Lb4 | Lb5 | Lb6w | Lb7w | Lb8w | |||||
| 1 | 129 | TGG/TTG | Trp/Leu | Trp | h50 | h67 | h67 | h57 | h50 | h33 | h75 | h75 |
| 2 | 146 | AAA/GAA | Lys/Glu | Lys | Glu | Glu | Glu | Glu | Glu | Glu | Glu | Glu |
| 3 | 174 | GTA/GCA | Val/Ala | Val | h45 | h58 | h17 | h33 | h71 | Val | Val | Val |
| 4 | 202 | GAG/GAC | Glu/Asp | Glu | Asp | h93 | Asp | Asp | Asp | h41 | h62 | Asp |
| 5 | 209 | TTT/CTT | Phe/Leu | Phe | Phe | Phe | Phe | Phe | Phe | h46 | h62 | h67 |
| 6 | 287-288 | AAA/CGA | Lys/Arg | Lys | h50 | h47 | h20 | h50 | h82 | Lys | h14/Lys | Lys |
| 7 | 392-394 | AAA/GAC | Lys/Asp | Lys | h35 | h53 | h33 | h33 | h67 | Lys | Lys | Lys |
| 8 | 617 | GAA/AAA | Glu/Lys | Glu | Glu | Glu | Glu | Glu | Glu | h57 | h75 | h75 |
| 9 | 648 | GCA/GGA | Ala/Gly | Ala | Ala | Ala | Ala | Ala | Ala | h57 | h12 | h14 |
| 10 | 688 | AAC/AAT | Asn/Asn (sIlent) | Asn | h41 | h36 | h57 | h50 | h25 | Asn | Asn | Asn |
| 11 | 694 | TGC/TGT | Cys/Cys (sIlent) | Cys | h41 | h36 | h57 | h43 | h14 | Cys | Cys | Cys |
| 12 | 799 | GTA/GTG | Val/Val (sIlent) | Val(GTA) | Val(GTA) | Val(GTA) | Val(GTA) | Val(GTA) | Val(GTA) | h50 | h40 | Val(GTG) |
| 13 | 824 | TCT/CCT | Ser/Pro | Pro | Pro | Pro | Pro | Pro | Pro | h43 | h45 | Pro |
| 14 | 864 | TAC/TTC | Tyr/Phe | Tyr | Tyr | Tyr | Tyr | Tyr | Tyr | h50 | h33 | Tyr |
| 15 | 891 | AGT/ATT | Ser/Ile | Ser | h53 | h58 | h56 | h78 | h67 | Ser | Ser | Ser |
| 16 | 1023 | TGG/TTG | Trp/Leu | Trp | h6 | Trp | Trp | Trp | Trp | h50 | h71 | h43 |
| 17 | 1030 | GGA/GGT | Gly/Gly (sIlent) | Gly (GGA) | h58 | h22 | h43 | h56 | h33 | Gly (GGA) | Gly (GGA) | Gly (GGA) |
| 18 | 1049 | TGG/GGG | Trp/Gly | Trp | h38 | h78 | h43 | h44 | h71 | Trp | Trp | Trp |
| 19 | 1076 | ATT/GTT | Ile/Val | Ile | Ile | Ile | Ile | Ile | Ile | h40 | h50 | h67 |
| 20 | 1150 | TTT/TTA | Phe/Leu | Phe | Phe | Phe | Phe | Phe | Phe | h29 | h50 | h50 |
| 21 | 1151 | ATG/GTG | Met/Val | Met | Met | Met | Met | Met | Met | h29 | h50 | h50 |
| 22 | 1266 | CCT/CTT | Pro/Leu | Pro | h64 | h75 | h60 | h64 | h50 | Pro | Pro | Pro |
| 23 | 1270 | GAA/GAT | Glu/Asp | Glu | Asp | Asp | Asp | Asp | Asp | h60 | h62 | Asp |
| 24 | 1274 | GAG/AAG | Glu/Lys | Glu | Glu | Glu | Glu | Glu | Glu | h60 | h57 | Lys |
| 25 | 1304 | ACT/TCT | Thr/Ser | Thr | h64 | h73 | h71 | h86 | h40 | Thr | Thr | Thr |
| 26 | 1338 | TGT/TCT | Cys/Ser | Cys | h36 | h23 | h37 | Ser | h71 | Cys | Cys | Cys |
| Nr. crt. | SNP Position in Coding Sequence | Codon | Amino Acid | Reference Genome ASM1917538v2 | Cultivated Genotypes | Wild Genotypes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lb1 | Lb2 | Lb3 | Lb4 | Lb5 | Lb6w | Lb7w | Lb8w | |||||
| 1 | 473 | CCT/CCC | Pro/Pro (silent) | Pro | Pro | Pro | Pro | Pro | Pro | h75 | h56 | Pro |
| 2 | 479 | TAC/TAT | Tyr/Tyr (silent) | Tyr (TAC) | Tyr(TAT) | Tyr(TAT) | Tyr(TAT) | Tyr(TAT) | Tyr(TAT) | h50 | h44 | h33 |
| 3 | 498 | GCC/ACC | Ala/Thr | Ala | h61 | h41 | h67 | h67 | h17 | h40 | h50 | Ala |
| 4 | 548 | TGT/TGC | Cys/Cys (silent) | Cys (TGT) | Cys (TGT) | Cys (TGT) | Cys (TGT) | Cys (TGT) | Cys (TGT) | h50 | h54 | Cys (TGT) |
| 5 | 555-557 | TCT/GCT/TCA/TCG | Ser/Ala/Ser/Ser | Ser | Ala | Ala | Ala | Ala | Ala | h50 | h46 | h33 |
| 6 | 563 | TCT/TCC | Ser/Ser (silent) | Ser (TCT) | Ser(TCC) | Ser(TCC) | Ser(TCC) | Ser(TCC) | Ser(TCC) | h67 | h46 | Ser(TCT) |
| 7 | 625 | GCG/GTG | Ala/Val | Ala | Val | Val | Val | Val | Val | Ala | h70 | h40 |
| 8 | 627 | GCT/TCT | Ala/Ser | Ala | Ser | Ser | Ser | Ser | Ser | Ser | Ser | h40 |
| 9 | 643 | TTC/TCC | Phe/Ser | Phe | Ser | Ser | Ser | Ser | Ser | Ser | Ser | Ser |
| 10 | 668 | CTT/CTC | Leu/Leu (silent) | Leu (CTT) | Leu(CTC) | Leu(CTC) | Leu(CTC) | Leu(CTC) | Leu(CTC) | h33 | h60 | h50 |
| 11 | 713 | TCG/TCC | Ser/Ser (silent) | Ser (TCG) | Ser(TCC) | Ser(TCC) | Ser(TCC) | Ser(TCC) | Ser(TCC) | h25 | h33 | h75 |
| 12 | 977 | TCG/TCA | Ser/Ser (silent) | Ser (TCG) | Ser (TCA) | Ser (TCA) | Ser (TCA) | Ser (TCA) | Ser (TCA) | h33 | h20 | h60 |
| 13 | 982 | TAT/TGT | Tyr/Ser | Tyr | h50 | h67 | h50 | h50 | h50 | Tyr | Tyr | Tyr |
| 14 | 986 | CGG/CGA | Arg/Arg (silent) | Arg (CGG) | Arg (CGA) | Arg (CGA) | Arg (CGA) | Arg (CGA) | Arg (CGA) | h33 | h20 | h63 |
| 15 | 1076 | GAG/GAA | Glu/Glu (silent) | Glu(GAG) | Glu (GAA) | Glu (GAA) | Glu (GAA) | Glu (GAA) | Glu (GAA) | h33 | h20 | h70 |
| 16 | 1079 | AAA/AAG | Lys/Lys (silent) | Lys (AAA) | h58 | h47 | h40 | h43 | h67 | h25 | h25 | h70 |
| 17 | 1223 | CCA/CCC | Pro/Pro (silent) | Pro (CCA) | h50 | h48 | h22 | h60 | h57 | Pro (CCA) | Pro (CCA) | Pro (CCA) |
| 18 | 1256 | AGG/AGA | Arg/Arg (silent) | Arg (AGG) | h41 | h56 | h71 | h56 | h50 | h37 | h67 | Arg(AGA) |
| 19 | 1285 | GTG/GCG | Val/Ala | Val | h42 | h44 | h67 | h60 | h62 | h37 | h775 | Ala |
| 20 | 1304 | TCG/TCT | Ser/Ser (silent) | Ser(TCG) | h65 | h48 | h17 | h50 | h33 | Ser (TCG) | Ser (TCG) | Ser (TCG) |
| 21 | 1394 | CTG/CTC | Leu/Leu (silent) | Leu(CTG) | Leu(CTC) | Leu(CTC) | Leu(CTC) | Leu(CTC) | Leu(CTC) | h50 | h50 | Leu(CTC) |
| 22 | 1421 | TTA/TTG | Leu/Leu (silent) | Leu (TTA) | h67 | h62 | h60 | Leu (TTG) | Leu (TTA) | Leu (TTA) | Leu (TTA) | Leu (TTA) |
| 23 | 1466 | ACT/ACA | Thr/Thr (silent) | Thr(ACT) | Thr (ACA) | Thr (ACA) | Thr (ACA) | Thr (ACA) | Thr (ACA) | h50 | h40 | h57 |
| 24 | 1550 | ACA/ACG | Thr/Thr (silent) | Thr(ACA) | Thr (ACG) | Thr (ACG) | Thr (ACG) | Thr (ACG) | Thr (ACG) | h33 | h50 | h57 |
| 25 | 1580 | ATC/ATA | Ile/Ile (silent) | Ile (ATC) | Ile (ATA) | Ile (ATA) | Ile (ATA) | Ile (ATA) | Ile (ATA) | h33 | h57 | h71 |
| 26 | 1671-1672 | GGA/AGA/GAA | Gly/Arg/Glu | Gly | h16 | h9 | h16 | h11 | h6 | Gly | Gly | Gly |
| 27 | 1689 | GCT/TCT | Ala/Ser | Ala | h23 | h15 | h21 | h20 | h24 | Ala | Ala | Ala |
| 28 | 1726 | ACA/AAA | Thr/Lys | Thr | h10 | h17 | h33 | h18 | h23 | h57 | h17 | h67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciceoi, R.; Asanica, A.; Luchian, V.; Iordachescu, M. Genomic Analysis of Romanian Lycium Genotypes: Exploring BODYGUARD Genes for Stress Resistance Breeding. Int. J. Mol. Sci. 2024, 25, 2130. https://doi.org/10.3390/ijms25042130
Ciceoi R, Asanica A, Luchian V, Iordachescu M. Genomic Analysis of Romanian Lycium Genotypes: Exploring BODYGUARD Genes for Stress Resistance Breeding. International Journal of Molecular Sciences. 2024; 25(4):2130. https://doi.org/10.3390/ijms25042130
Chicago/Turabian StyleCiceoi, Roxana, Adrian Asanica, Vasilica Luchian, and Mihaela Iordachescu. 2024. "Genomic Analysis of Romanian Lycium Genotypes: Exploring BODYGUARD Genes for Stress Resistance Breeding" International Journal of Molecular Sciences 25, no. 4: 2130. https://doi.org/10.3390/ijms25042130